糯高粱F2群体农艺性状数量遗传分析
2023-06-14姜昱雯张国兵周棱波邵明波
姜昱雯,赵 强,张国兵,周棱波,汪 灿,邵明波
(贵州省旱粮研究所,贵州 贵阳 550006)
0 引言
【研究意义】高粱〔Sorghumbicolor(L.) Moench〕是世界五大粮食作物之一,也是当前农业产业结构调整、生产方式转变的理想作物[1]。我国高粱常被用作饲料、酿制酒和醋的原料以及其他的工业原料,因此在保证国家粮食安全和经济发展中起着重要作用。近年来,由于受经济全球化和我国一系列政策影响,我国在国际上的贸易较为频繁,尤其是经济作物的贸易量逐年上升,贸易量加大的同时,也刺激着国内对经济作物的种植,高粱作为其中代表之一,在国内外大环境和白酒行业需求量影响下,我国高粱播种面积逐年增长。贵州省高粱主栽品种如红缨子、红珍珠等的穗型和株型结构限制高粱单产水平的提高,不利于机械化生产,培育高产优质高粱品种成为首要攻坚任务[2]。高产优质品种受多个表型性状综合影响,且各性状之间相互关联、彼此制约,要想培育高产高粱品种,必须对与高粱产量相关的农艺性状进行分析。因此,分析高粱农艺性状遗传特性,对优质高产高粱新品种选育具有重要意义。【前人研究进展】不同种质资源的农艺性状存在较大差异,应根据不同的育种目标筛选基础材料[3]。李萌等[4]对1 285份山西省高粱地方品种18个农艺性状分析,获得代表山西省高粱地方品种的核心种质198份,为地方品种评价和利用奠定了种质基础。乔靖等[5]对粒用高粱种质资源主要农艺性状的相关性及主成分分析表明,不同粒用高粱种质资源农艺性状间存在较大差异,应根据不同育种目标筛选基础材料。王官等[6]对15份甜高粱种质资源的7个农艺性状进行分析,明确了甜高粱种质资源的多样性,对提高甜高粱育种效率具有重要的促进作用。【研究切入点】贵州省盛产优质酱香型白酒,白酒产业已成为贵州省的特色产业、优势产业和支柱产业之一,糯高粱是酿酒的主要原料,前人针对酒用糯高粱进行了农艺性状数量遗传研究,但到目前为止关于酒用糯高粱的遗传解析未见报道。【拟解决的关键问题】选取湘10721和青稞洋2个糯高粱品种作亲本进行杂交,以F2群体单株的旗叶长、旗叶宽、茎粗、株高、穗长及穗柄长作为参数进行主基因-多基因分析,探究其遗传规律,为酒用高粱新品种选育提供基础材料。
1 材料与方法
1.1 试验材料
青稞洋为酱香型酒用有机高粱,湘10721为四川骨干自交系。选取湘10721和青稞洋2个糯高粱品种作亲本进行杂交,以亲本、F1及F2群体单株作为研究对象。试验材料均由贵州省农业科学院旱粮研究所提供。
1.2 试验方法
2021年8月在贵州省农业科学院试验地(26°3′N,106°6′E)进行亲本杂交,配制正反交组合,同年在海南试验田进行加代繁殖,2022年初获得F2群体种子。2022年5月在安顺山京农场(26°1′N,106°1′E)按顺序播种亲本、F1、F2,其中,亲本各种植2行,F1正反交各种植4行,F2分离群体种植20行,每行24株。田间种植行长3.5 m,行间距0.5 m,株距0.4 m,水肥管理同常规。高粱开花20 d后,参照《高粱种质资源描述规范和数据标准》[7]中的有关方法,分别对亲本、F1、F2群体的株高、茎粗、旗叶长、旗叶宽、穗长和穗柄长等性状进行考察。从每行第3株开始,连续测量5株。
1.3 数据统计与分析
常规数据通过Microsoft Excel 2010和SPSS Statistics 17.0进行统计分析。F2世代分析:根据单个分离世代群体的遗传模型方法[8]、数量性状主基因+多基因混合遗传分析方法[9],得到遗传模型以及AIC值,选择AIC值较小的模型作为备选模型;根据适合性检验结果确定最优模型,计算最优遗传模型的主基因效应值和主基因遗传率等遗传参数,分析糯高粱F2群体农艺性状数量遗传规律。
2 结果与分析
2.1 糯高粱农艺性状的相关性
由表1可知,糯高粱的旗叶长与旗叶宽、茎粗、株高、穗长、穗柄长均呈极显著负相关,相关系数分别为-0.889、-0.889、-0.632和-0.814和-0.549;旗叶宽与茎粗、株高、穗长、穗柄长均呈极显著正相关,相关系数分别为0.918、0.735、0.834和0.676;茎粗与株高、穗长、穗柄长呈极显著正相关,相关系数分别为0.757、0.830和0.637;株高与穗长、穗柄长均呈极显著正相关,相关系数分别为0.616和0.564;穗长与穗柄长呈极显著正相关,相关系数为0.537。F2群体6个性状之间均存在显著相关性,说明性状间存在一定关联。
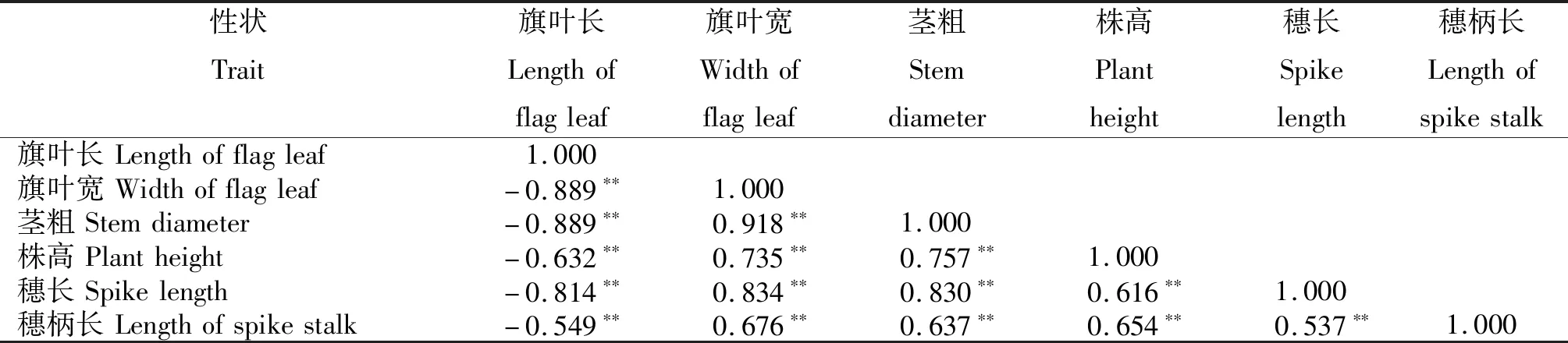
表1 糯高粱F2群体农艺性状的相关系数Table 1 Correlation coefficients of agronomic traits of F2 from waxy sorghum
2.2 糯高粱F2农艺性状的变异性
由表2可知,糯高粱F2群体6个农艺性状的变异系数差异较大,具有丰富的遗传变异信息。变异系数大小依次为穗柄长、旗叶宽、旗叶长、株高、茎粗、穗长。其中,穗柄长、旗叶宽、旗叶长的变异系数较大,分别为31.27%、20.03%、17.25%,说明这3个性状在糯高粱F2中差异较大,遗传改良潜力较大。结合各指标性状的分布区间与分布频数(图1)可知,该6个农艺性状均呈连续正态分布,符合数量性状遗传特征。

表2 糯高粱F2的农艺性状统计值Table 2 Statistical value of agronomic traits of F2 from waxy sorghum
2.3 糯高粱F2群体农艺性状的遗传模型
2.3.1 遗传模型选择 根据 AIC值最小原则,在6个性状的11个遗传模型中,选择AIC值较低的4个模型进行遗传模型检验。从表3看出,旗叶长选出的模型为0MG、1M-A、2MG-ADI和2MG-EA模型,旗叶宽选出的模型为0MG、2MG-ADI、2MG-CD和2MG-EAD,茎粗选出的模型为2MG-ADI、2MG-AD、2MG-A和2MG-EA,株高选出的模型为2MG-ADI、2MG-AD、2MG-A和2MG-EA,穗长选出的模型为2MG-A、2MG-EA、2MG-CD和2MG-EAD,穗柄长选出的模型为2MG-ADI、2MG-AD、2MG-A和2MG-EA模型。
2.3.2 遗传模型检验 依据均匀性检验(U12、U22、U32)、Smirnov检验(nW2)和Kolmogorov检验(Dn),分别对旗叶长、旗叶宽、茎粗、株高、穗长及穗柄长6个农艺性状的候选模型进行适合性检验,选出6个性状的最适遗传模型(表4)。
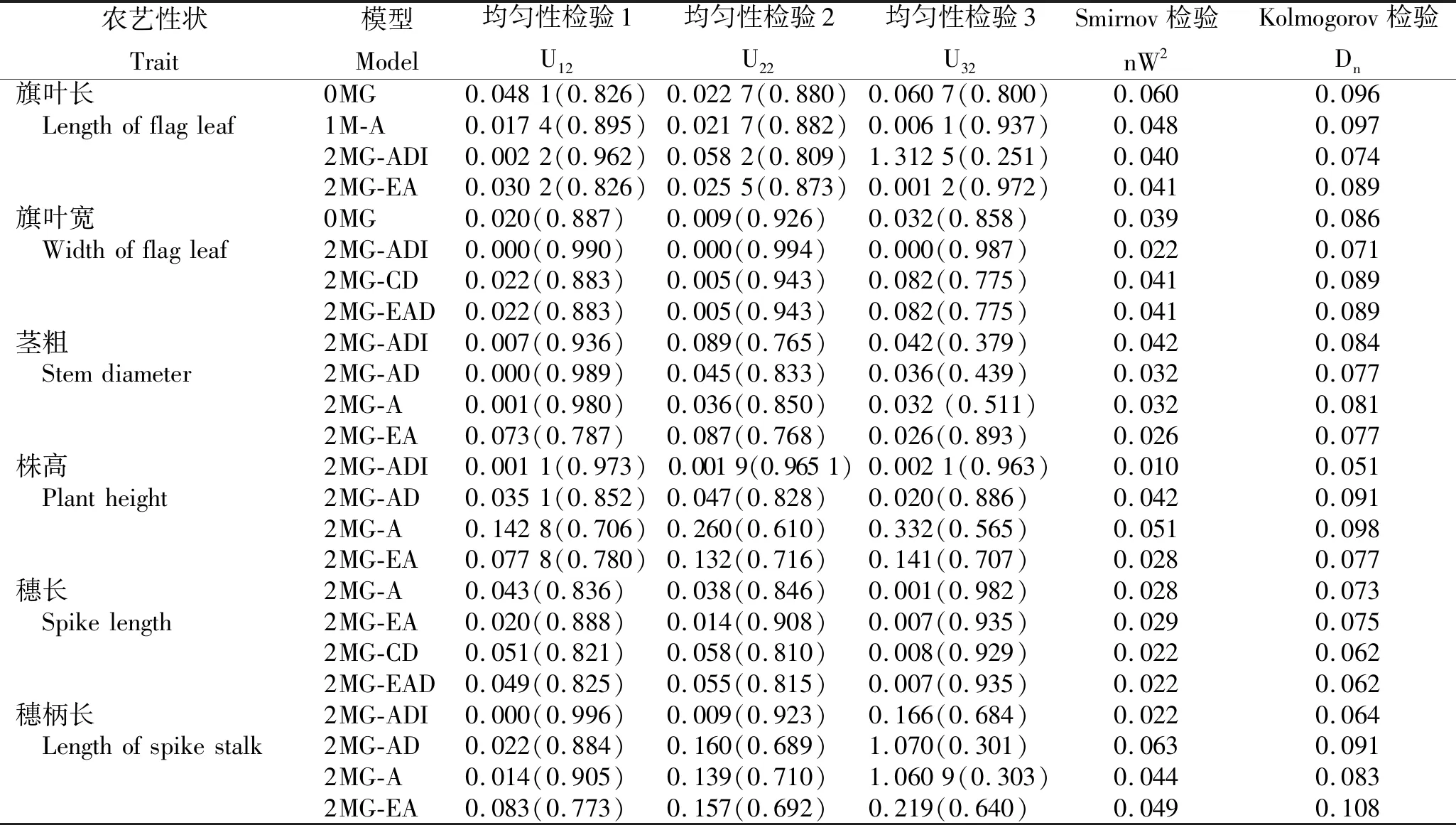
表4 F2群体各农艺性状备选模型的适合性检验结果Table 4 Suitability test results of candidate models for agronomic traits of F2 population
1) 旗叶长遗传模型。在旗叶长的 4种备选模型中,Kolmogorov检验均未达显著水平,Smirnov检验中除0MG模型外,均达显著水平(P<0.05),且相较于其他3种模型,以 2MG-ADI的AIC值最小。因此,选择 2MG-ADI作为该群体旗叶长的最适遗传模型。说明,旗叶长是受2对主基因控制的性状,为加性-显性-上位性混合遗传模型。
2) 旗叶宽遗传模型。在旗叶宽的4种备选模型中,Kolmogorov检验均未达显著水平,Smirnov检验均达显著水平(P<0.05),且相较于其他3种模型,以0MG的AIC值最小。因此,选择 0MG作为该群体旗叶宽的最适遗传模型。说明,旗叶宽是仅受微效多基因控制而不受主基因控制的性状。
3) 茎粗遗传模型。在茎粗的4种备选模型中,Kolmogorov检验均未达显著水平,Smirnov检验均达显著水平(P<0.05),且相较于其他3种模型,以2MG-ADI的AIC值最小。因此,选择2MG-ADI作为该群体茎粗的最适遗传模型。说明,茎粗是受2对主基因控制的性状,为加性-显性-上位性混合遗传模型。
4) 株高遗传模型。在株高的 4种备选模型中,Kolmogorov检验均未达到显著水平,Smirnov检验中除2MG-A模型外,其他模型均达到显著水平(P<0.05),且相较于其他3种模型,以2MG-ADI的AIC值最小。因此,选择2MG-ADI作为该群体株高的最适遗传模型。说明,株高是受2对主基因控制的性状,为加性-显性-上位性混合遗传模型。
5) 穗长遗传模型。在穗长的 4种备选模型中,Kolmogorov检验均未达到显著水平,Smirnov检验均达到显著水平(P<0.05),且相较于其他3种模型,以2MG-CD的AIC值最小。因此,选择2MG-CD作为该群体穗长的最适遗传模型。说明,穗长是受2对主基因控制的性状,为完全显性遗传模型。
6) 穗柄长遗传模型。在穗柄长的 4种备选模型中,Kolmogorov检验均未达到显著水平,Smirnov检验中除2MG-AD模型外,其他模型均达到显著水平(P<0.05),且相较于其他3种模型,以2MG-A的AIC值最小。因此,选择2MG-A作为该群体穗柄长的最适遗传模型。说明,穗柄长是受2对主基因控制的性状,为加性遗传模型。
2.4 遗传参数估计
根据已确定的最优模型对 F2群体农艺性状进行遗传参数估计。从表5看出,旗叶长的群体均方为51.35,加性效应之和为14.36,显性效应之和为-19.65,显性效应绝对值明显大于加性效应,说明应以显性效应为主对旗叶长加以控制,主基因遗传率为 99.90%;茎粗的群体均方为14.50,控制茎粗的2对主基因的加性效应之和为2.18,显性效应之和为0.11,加性效应明显大于显性效应,说明应以加性效应为主对茎粗加以控制,主基因遗传率为 99.99%;株高的群体均方为188.48,控制株高的2对主基因的加性效应都为正向效应,加性效应之和为35.75,显性效应之和为-21.50,加性效应明显大于显性效应,说明应以显性效应为主对株高加以控制,主基因遗传率为 99.72%;穗长的主基显性效应之和为2.65,主基因遗传率为95.83%。穗柄长的加性效应之和为5.76,主基因遗传率为 99.79%。
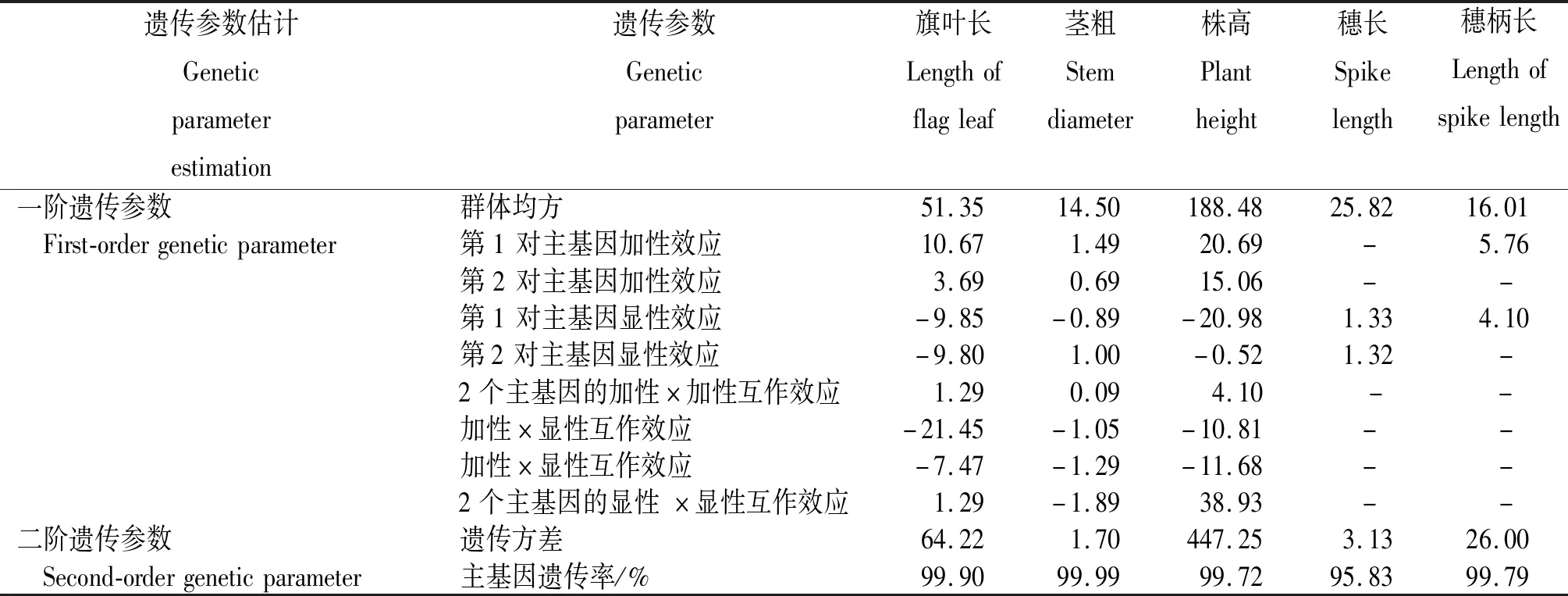
表5 各农艺性状遗传模型的参数估计Table 5 Parameter estimation of genetic model for agronomic traits
3 讨论
糯高粱F2群体旗叶长、茎粗、株高均由2对主基因控制且主基因遗传率均大于99.5%,表明后代遗传比较稳定,不易受环境影响。其中,旗叶长的研究结果与卢华雨等[10]的研究结果一致,符合加性-显性-上位性混合遗传模型;茎粗的数量遗传分析结果与栾金花等[11]对中国高粱地方品种的研究结果一致。李佳俊等[12]研究表明,茎粗多以2对主基因的加性、显性、上位效应为主。但赵博等[13]研究表明,茎粗符合无主基因而仅有多基因存在的A-0模型,受微效多基因控制。对糯高梁株高的数量遗传分析表明,F2代分离群体中,株高符合Model A-0模型,无主基因控制,属于微效多基因控制的数量性状,与白晓倩等[14]的研究结果不同。原因可能是由于一方面选择杂交的亲本不同,另一方面种植环境不同[15]。糯高粱穗长由2对主基因控制,属完全显性遗传模型,主基因遗传率为95.83%,与白晓倩等[14,16]的研究不一致,白晓倩[14]研究结果显示,高梁穗长符合2对主基因+多基因控制的等显性模型;于澎湃等[16]研究表明,粒用高粱穗长和旗叶鞘长不受主基因控制,属微效多基因遗传。穗柄长是受2对主基因控制的性状,属加性遗传,与王立新[17]的研究结果一致。
4 结论
糯高粱湘10721和青稞洋杂交F2群体的旗叶长、茎粗及株高均为加性-显性-上位性混合遗传模型,主基因遗传率分别为99.90%、99.99%、99.72%;穗长为完全显性遗传模型,主基因遗传率为95.83%;穗柄长为加性混合遗传模型,主基因遗传率为99.79%;旗叶宽遗传不受主基因控制,为受微效多基因控制的性状。旗叶长、茎粗、株高、穗长、穗柄长等农艺性状的遗传率较高,能够稳定遗传,受环境因素影响较小。旗叶宽受微效多基因遗传,在后代中遗传不稳定,受环境因素影响较大。为获得理想育种材料,尤其应注意环境因素对数量性状产生的影响。
