烟草NtCCX2 基因的克隆与表达分析
2023-06-14杨园园郝浩浩王平平金维环郭红祥
冯 康,杨园园,刘 琼,程 岚,郝浩浩,王平平,金维环,郭红祥
(1.河南农业大学 生命科学学院,河南 郑州 450002;2.河南省烟草公司 驻马店市公司,河南 驻马店 463000;3.中国烟草总公司 陕西省公司,陕西 西安 710061)
随着工业化的发展,重金属镉(Cd)造成的土壤污染问题越来越严重。研究表明,Cd 具有高聚集、难降解、毒性强等特性,对生物生长发育和食品安全均能造成威胁。植物中过量积累的Cd 会抑制种子发芽、叶绿素合成、植物生长等,对幼苗产生不利影响[1]。Cd 也很容易进入农作物的可食用部分,进而在人体中富集。进入人体的Cd 主要积累分布在肝脏、胰腺和骨骼中,引发肾衰竭、心血管和内分泌失调等疾病。另外,Cd 还可以破坏人体骨骼系统,引起骨骼疏松,严重威胁人体健康[2]。因此,将植物的Cd 含量降低到安全水平,减轻Cd 对人体健康的损害,是降低Cd污染危害切实可行的方法[3]。
植物主要利用锌(Zn2+)、锰(Mn2+)和铁(Fe2+)等离子的转运体来实现对Cd的吸收和转运。例如:水稻OsNramp5 是在根系中表达的膜定位Mn2+转运体,研究数据显示,水稻OsNramp5突变体植株中Cd总含量和籽粒中Cd 积累量均呈现大幅度降低的现象,表明其在Cd 吸收过程中发挥作用[4]。IRT1 为铁的转运载体,可以将重金属Cd 运往植物茎部[5]。阳离子/钙离子(Ca2+)交换器(Cation/Ca2+exchangers,CCX)属于Ca2+/阳离子反转运蛋白(Ca2+/cation antiporter,CaCA)超家族,能通过氢离子(H+)、钾离子(K+)或钠离子(Na+)等阳离子逆电化学梯度交换Ca2+[6]。CCX 广泛存在于细菌、植物和动物中,不仅具有转运离子的功能,而且在响应外界环境胁迫的过 程 中 起 着 重 要 作 用[7‐8]。AtCCX1、AtCCX3和AtCCX4是拟南芥中CCX 家族成员。AtCCX1在拟南芥叶片衰老过程中高度表达,基因敲除AtCCX1和AtCCX4能使叶片保持常绿,而过表达AtCCX1则加速叶片衰老[9]。Ca2+缺失的条件下,AtCCX1和AtCCX4突变体幼苗均表现出明显的生长障碍,表明AtCCX1可能通过Ca2+信号来调节叶片的衰老[9]。AtCCX3和AtCCX4定位于液泡膜,显示出H+依赖的K+、Mn2+和K+运输调节能力[10]。在烟草中过表达AtCCX3后发现,转基因烟草中积累了大量的阳离子[10]。在水稻基因组中,目前共发现4个CCX 成员,其中OsCCX2被证明能增强酵母和拟南芥的Cd 耐受性,进一步研究发现,OsCCX2能将Cd2+装载到木质部中,通过介导Ca2+转运途径促进水稻籽粒中Cd元素的积累[11]。目前,烟草CCX 基因家族成员及其功能还未见报道。
烟草是一种重要的叶用经济作物,同时也是Cd富集植物,吸收的Cd主要累积于烟叶中[12]。植烟土壤中过量的Cd不仅会抑制烟株生长发育、降低烟叶的产量和品质,烟叶中的Cd还会通过抽吸过程中产生的主流烟气被人体所吸收,造成潜在危害。有文献报道,水稻OsCCX2参与Cd的转运[11]。因此,拟克隆烟草NtCCX2基因及其启动子,研究该基因对Cd吸收、转运与累积的影响,探讨通过调节NtCCX2降低烟叶Cd含量的途径,为优质烟叶生产提供理论依据,以期为基因编辑改良烟草品种找到新靶标。
1 材料和方法
1.1 试验材料与试剂
1.1.1 试验材料 野生型K326 土培烟草(Nicotiana tabacumL.)种植在河南农业大学温室(东经113°79′,北纬34°79′),温度26~28 ℃,相对湿度60%,光周期为16 h光照/8 h黑暗。
组织特异性表达试验:野生型K326 土培烟草生长到幼苗期、旺长期和成熟期时,取根、茎、叶样品,置于-80 ℃冰箱中保存,用于RNA的提取。
镉胁迫试验:在八叶一心时期,选取长势大小一致的18 株野生型K326 土培烟草,平均分为3 组,用浓度为0、200、1 000 µmol/L 的氯化镉溶液浇灌,每组处理浇灌400 mL,每隔1 d 浇1 次,处理7 d 后取叶和根,置于-80 ℃冰箱中保存,用于Cd 含量的测定和RNA的提取。
激素信号响应试验:在六叶一心时期,选取长势大小一致的25 株野生型K326 土培烟草,分别用脱落酸(ABA)、乙烯利和茉莉酸甲酯(MeJA)处理,浓 度 均 为4 mmol/L[13‐15],每 株 喷 洒20 mL,分 别 在ABA 和乙烯利处理后0、2、4、8、24 h 取烟草的叶和根,在MeJA 处理后0、4、8、24 h 取烟草的叶和根,置于-80 ℃冰箱中保存,用于RNA的提取。
干旱胁迫与盐胁迫试验:在烟草旺长期,选取长势一致的9 株野生型K326 土培烟草,其中3 株正常浇水作为对照(CK),其余的分别进行200 mmol/L NaCl 和干旱7 d 胁迫处理[16],观察其表型变化,处理7 d后取叶和根置于-80 ℃冰箱中保存,用于RNA的提取。
降镉试验:大田试验品种为中烟100,种植于驻马店泌阳县,设置4 个处理:CK、喷洒降镉灵(佛山市铁人环保科技有限公司)、喷洒富硒宝(四川爱隆植物营业科技有限公司)、喷洒微量元素肥(山东海岱绿洲生物工程有限公司)。分别在移栽后30、60 d 按照使用说明喷施降镉灵、富硒宝、微量元素肥。喷施7 d 后取样,置于-80 ℃保存,用于Cd 含量的测定和RNA的提取。
1.1.2 试剂 DNA凝胶回收试剂盒和KK快提RNA试剂盒购自北京庄盟国际生物基因科技有限公司;质粒小提试剂盒购自天根生化科技有限公司;限制性内切酶购于NEB 有限公司;cDNA 合成试剂盒MonScriptTMRT ⅢSuper Mix with dsDNase(Two-Step)购自莫纳生物科技有限公司;大肠杆菌(Escherichia coli)菌株DH5α 购自上海昂羽生物技术有限公司。
1.2 方法
1.2.1 烟草DNA、总RNA 的提取及cDNA 的合成采用CTAB 法提取野生型K326 土培烟草叶片中的DNA。利用庄盟公司植物KK 快提试剂盒提取烟草根、茎、叶中的RNA,利用Nanodrop 仪器测RNA 浓度,经琼脂糖电泳检测RNA 降解和污染情况。用莫纳生物试剂反转录得到cDNA,于-80 ℃冰箱中保存用于基因的克隆和荧光定量PCR。
1.2.2 烟草NtCCX2基因的克隆及测序 在NCBI网站查找烟草基因组中NtCCX2基因序列,利用NCBI 网站中的Primer-BLAST 设计引物CCX2-F/CCX2-R(表1)。提取野生型K326 土培烟草叶片中的RNA,反转录得到cDNA,进行PCR,扩增体系:2×TaqPlus Master MixⅡ25µL,上、下游引物和cDNA模板各1µL,以无菌水补足50µL。PCR 反应条件:95 ℃预变性5 min;95 ℃变性15 s,56 ℃退火20 s,72 ℃延伸2.25 min,33 个循环;72 ℃延伸8 min。利用庄盟公司的DNA 凝胶回收试剂盒对扩增的目的片段进行回收,回收产物送至尚亚生物技术有限公司进行测序。
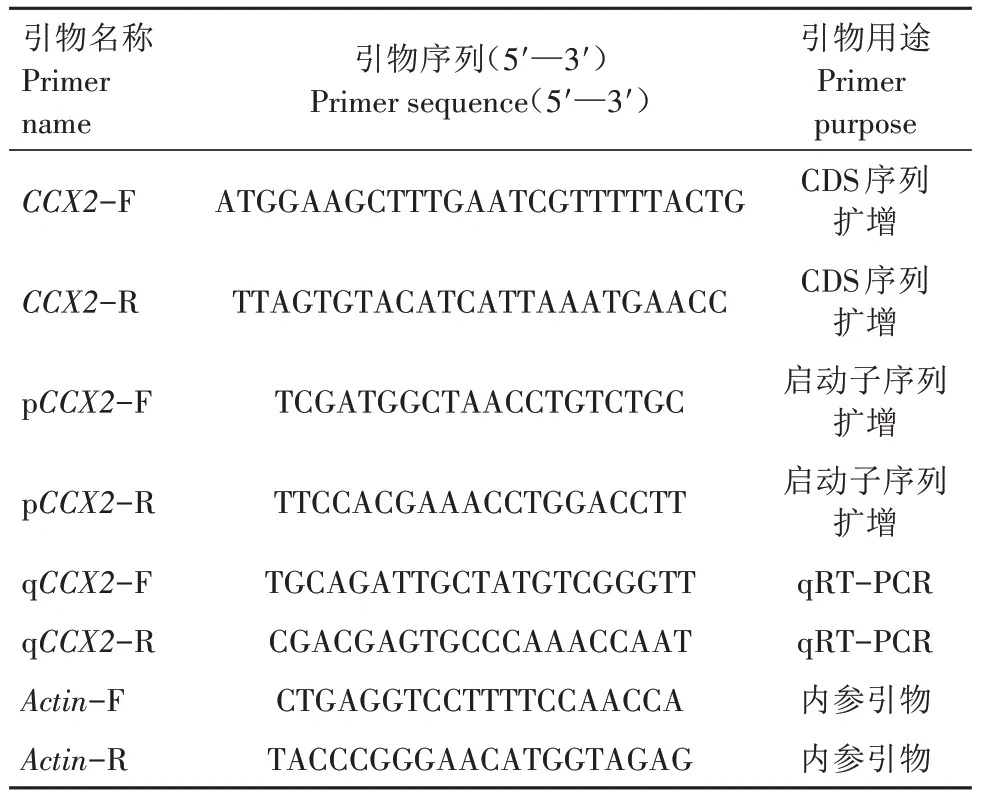
表1 本研究所用引物信息Tab.1 Information of primers used in this study
1.2.3 烟草NtCCX2基因的生物信息学分析 利用DNAMAN 软件将NtCCX2基因的核苷酸序列翻译成氨基酸序列;根据其氨基酸序列在NCBI(http://www.ncbi.nlm.nih.gov/)的BLASTp 上 搜 索 同 家 族CCX 蛋白;利用MEGA 7.0 构建不同植物CCX 蛋白进化树;用NCBI 的Conserved domain 预测蛋白质结构域;利用DNAMAN 8 软件将烟草和其他物种的同家族CCX氨基酸序列进行比对。
1.2.4 烟草NtCCX2启动子的克隆 在NCBI 网站上查找烟草的基因组序列,将该基因上游2 000 bp左右片段作为其启动子区域,利用NCBI 网站中的Primer-BLAST 设计引物pCCX2-F/pCCX2-R(表1),以野生型K326 烟草基因组DNA 为模板,扩增启动子片段,将得到的目的片段导入到pMD19-T(TaKaRa,北京)中。
1.2.5 烟草NtCCX2启动子序列分析 通过PlantCARE 网站分析NtCCX2启动子中顺式作用元件位置、序列及功能。
1.2.6 烟草NtCCX2基因表达分析 利用引物设计软件Primer Premier 5.0 设计实时荧光定量引物qCCX2-F 和qCCX2-R(表1),以NtActin基因作为内参,用2×SYBR Premix WizTaq酶(诺贝莱,北京)进行荧光定量PCR,采用2-ΔΔCt法计算NtCCX2基因的相对表达量。
1.2.7 Cd 含量检测 委托河南华测检测技术有限公司采用石墨炉原子吸收分光光度法测定Cd含量。
2 结果与分析
2.1 烟草NtCCX2基因的克隆与序列分析
在NCBI 网站上查找拟南芥AtCCX2的氨基酸序列,利用BLASTp查找到烟草CCX2同源基因。该基因全长2 464 bp,CDS 区1 926 bp,编码641 个氨基酸。以野生型K326 土培烟草的叶片cDNA 为模板进行PCR,得到1 条长度为1 926 bp 的条带(图1),该基因编码641 个氨基酸,GenBank 登录号为XM_016577905。
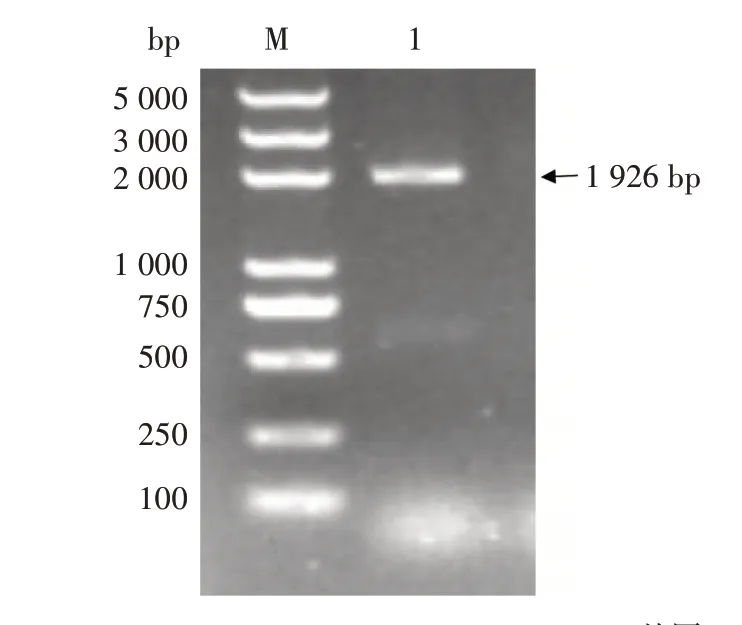
图1 烟草NtCCX2基因的克隆Fig.1 Cloning of NtCCX2 gene in tobacco
基于WOLF PSORT 亚细胞定位预测结果显示,该蛋白质最可能位于质膜。利用NCBI 的Conserved domain 预测NtCCX2 蛋白结构域发现,该蛋白质在23—439 位氨基酸含有1 个保守的Na+/Ca2+-K+INDEPENDENT EXCHANGER 结构域。NtCCX2 与其他物种CCX2 氨基酸序列比对的结果见图2。由图2可知,NtCCX2与马铃薯StCCX2和辣椒CaCCX2 的氨基酸序列相似性最高,并具有CCX家族高度保守的特征,在跨膜区有2 个α-重复区域:α1 模式基序GNGAPD 和α2 模式基序G(N/D)SxGD[17‐19]。利用MEGA 7.0 软件对NtCCX2 同家族21种植物的CCX2 蛋白构建进化树,结果见图3,烟草NtCCX2 在进化上与番茄(Solanum lycopersicum,XP 004243702.1)、辣 椒(Capsicum annuum,XP 016580161.1)、马 铃 薯(Solanum tuberosum,XP 006342390.1)、曼 陀 罗(Datura stramonium,MCD 9559449.1)CCX 蛋白相似度最高,说明其亲缘关系最近,与拟南芥(Arabidopsis thaliana,NP 564650.1)、水稻(Oryza sativa japonicagroup,XP 015632178.1)等亲缘关系次之,而与黄瓜(Cucumis sativus,XP 004144128.1)、冬 瓜(Benincasa hispida,XP 038878908.1)亲缘关系最远。
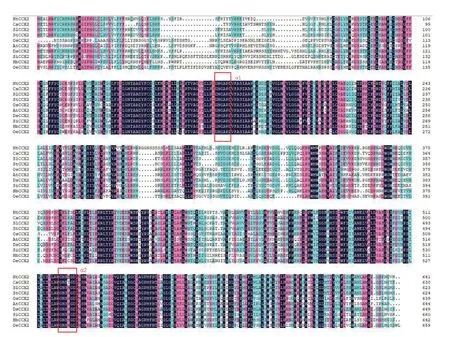
图2 CCX2蛋白序列比对分析Fig.2 Multiple sequence alignment of CCX2 proteins
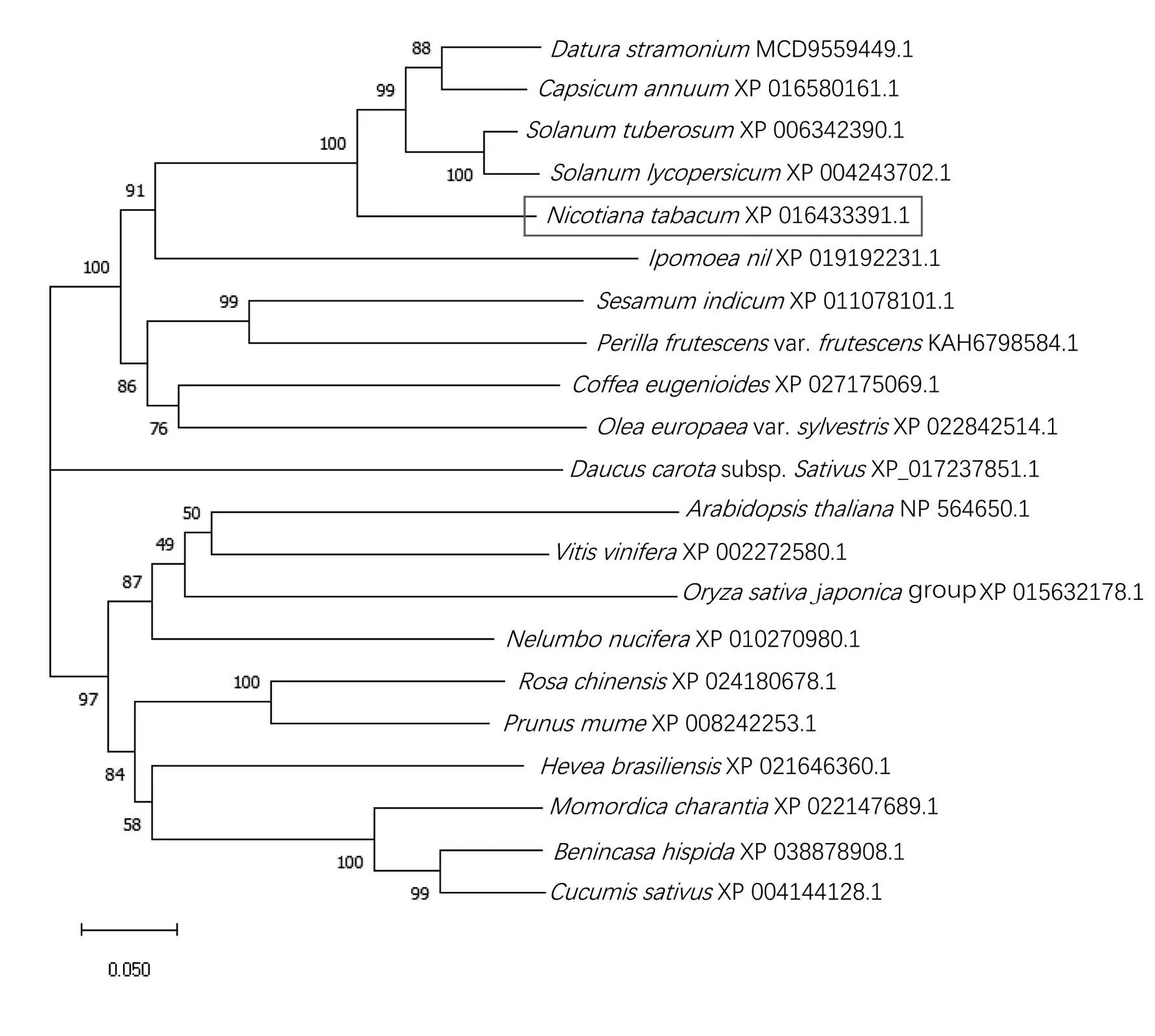
图3 植物CCX蛋白的系统进化分析Fig.3 Phylogenetic analysis of CCX proteins in plants
2.2 烟草NtCCX2启动子克隆及顺式作用元件分析
以野生型K326 土培烟草的叶片DNA 为模板扩增NtCCX2启动子,得到1 条长度为2 160 bp 的条带(图4),产物纯化后连接到T 载体并转化到大肠杆菌DH5α 中,经过菌液PCR 检测后送至尚亚生物技术有限公司测序,测序结果与烟草基因组的结果高度相似。
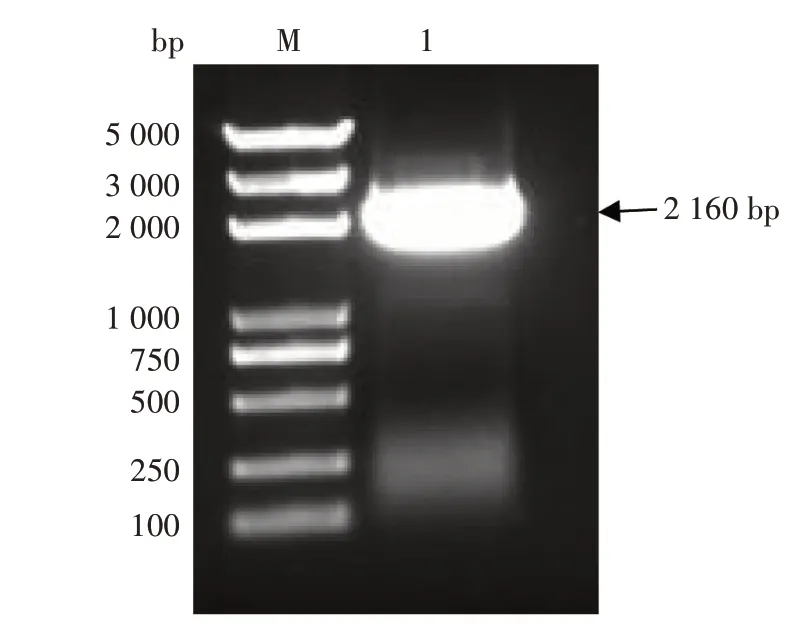
图4 烟草NtCCX2基因启动子的克隆Fig.4 Cloning of NtCCX2 promoter in tobacco
用PlantCARE 分析NtCCX2启动子序列顺式作用元件,结果见表2,NtCCX2启动子包含与光响应有关的元件AE-BOX 和一些植物激素响应元件,如脱落酸应答元件ABRE、茉莉酸应答元件CGTCAmotif、水杨酸响应元件TCA-element 等,表明光、茉莉酸、脱落酸和水杨酸等环境因素可能影响NtCCX2基因的表达。另外,该启动子区域还包含有植物逆境应答相关元件,如干旱、高盐、低温响应元件MYB,说明该基因可能与植物抗逆相关。

表2 烟草NtCCX2基因启动子部分顺式作用元件Tab.2 Cis‑acting elements of NtCCX2 promoter in tobacco
2.3 烟草NtCCX2基因的组织特异性表达分析
烟草NtCCX2基因表达具有明显的组织特异性。图5显示,幼苗期和旺长期NtCCX2基因在叶中的相对表达量最高,分别是茎中相对表达量的37.4、6.2 倍,而成熟期NtCCX2基因在根和花中的相对表达量明显高于茎和叶,分别是茎中相对表达量的5.9、4.1倍,是叶中相对表达量的3.6、2.4倍。
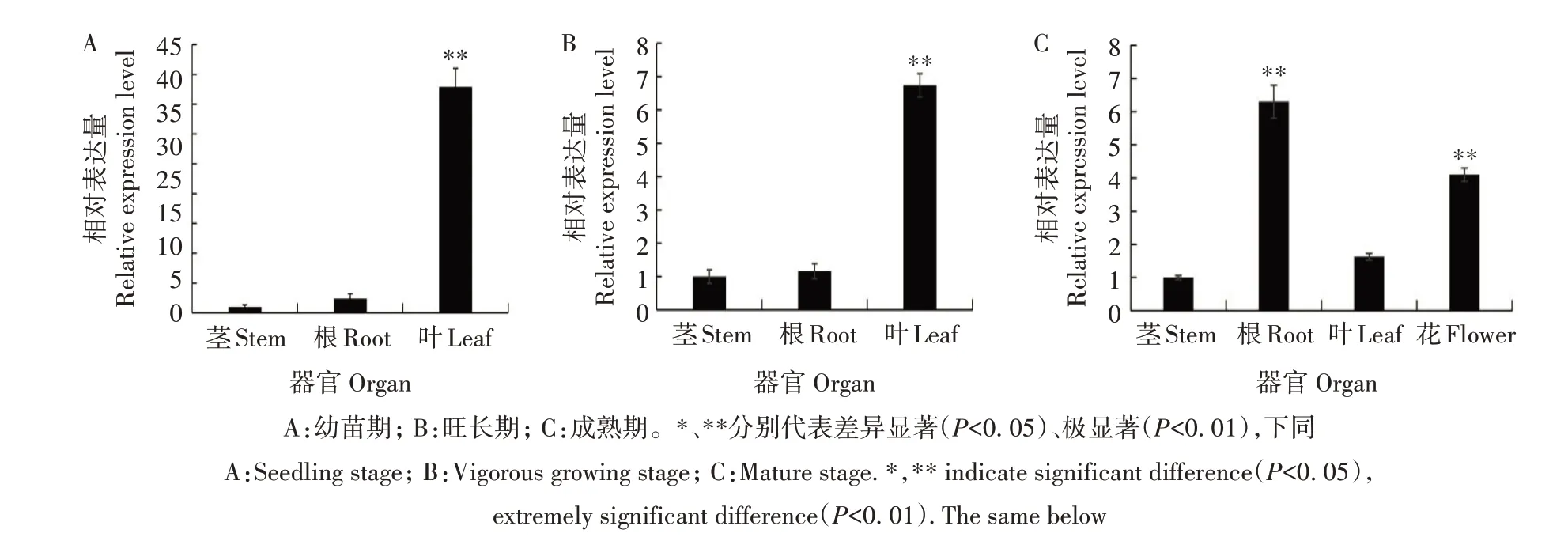
图5 NtCCX2基因的组织表达模式分析Fig.5 Expression pattern analysis of NtCCX2 gene
2.4 烟草NtCCX2基因对干旱、盐胁迫的响应
干旱与盐胁迫是2 种常见的非生物逆境胁迫,NtCCX2基因启动子中含有干旱与盐胁迫的响应元件。图6显示,干旱胁迫后叶中NtCCX2基因相对表达量是对照的2.1 倍。盐胁迫后叶中NtCCX2基因相对表达量是对照的5.1 倍,根中相对表达量是对照的3.4 倍。因此,干旱和盐胁迫均能促进烟草根和叶中NtCCX2基因的表达,在叶中的诱导效应大于根中,表明NtCCX2基因参与烟草的干旱与盐胁迫响应过程。
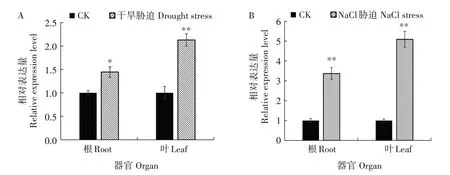
图6 干旱胁迫(A)和NaCl胁迫(B)下NtCCX2基因表达水平Fig.6 Expression of NtCCX2 under drought stress(A)and NaCl stress(B)
2.5 烟草NtCCX2基因对激素信号的响应
干旱、盐碱和低温等非生物胁迫会导致植物水分散失,即产生渗透胁迫,进而促进植物激素ABA的合成,因此,ABA 信号通路是植物非生物胁迫反应过程的中枢调节因子[20]。图7A、7B 显示,ABA 处理2 h 后,NtCCX2基因在根中相对表达量明显上调,是0 h 的3.7 倍;ABA 处理8 h 后,在叶中相对表达量是0 h 的2.1 倍。表明ABA 能够明显诱导烟草根和叶中NtCCX2基因的表达,在根中的诱导效应大于叶中,其具体机制尚需进一步研究。MeJA 是响应植物损伤的激素信号分子,外源应用能够激发植物防御基因的表达。图7C、7D 显示,MeJA 处理后,根中NtCCX2基因相对表达量没有明显的变化;MeJA 处理8 h 后,叶中NtCCX2基因相对表达量是0 h 的2.1 倍,表明MeJA 处理8 h 后能够显著诱导叶中NtCCX2基因表达。作为一种重要的植物激素,乙烯利不仅在植物生长发育过程中发挥重要作用,也参与植物对干旱、水淹、盐和低温等各种非生物胁迫的响应[21]。图7E、7F 显示,乙烯利处理2 h 后,NtCCX2基因在根和叶中相对表达量均增加;处理8 h 后,NtCCX2基因在根中的相对表达量是0 h 的2.4倍;处理24 h后,NtCCX2基因在叶中的相对表达量是0 h 的5.7 倍。表明乙烯利处理能够显著诱导NtCCX2基因的表达。
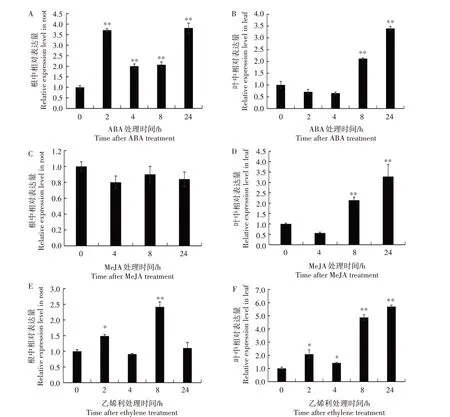
图7 烟草NtCCX2基因对ABA(A和B)、MeJA(C和D)和乙烯利(E和F)的响应Fig.7 Response of NtCCX2 to ABA(A and B),MeJA(C and D)and ethylene(E and F)
2.6 烟草NtCCX2基因对Cd胁迫的响应
烟草是一种Cd 富集植物,烟叶中Cd 的含量受到土壤Cd含量、水肥管理等众多因素的影响。温室试验结果显示(图8),200 µmol/L Cd 胁迫后,根中NtCCX2基因的相对表达量上调10 倍以上,叶中的表达量上调5 倍以上,1 000 µmol/L Cd 胁迫后根和叶中NtCCX2基因的相对表达量均上调10 倍以上,表明该基因的表达受Cd 胁迫的诱导。根和叶中的Cd 含量也明显高于对照,表明NtCCX2基因与烟株Cd 的吸收运输相关。土壤修复剂、植物生长调节剂、营养元素的添加以及现代生物技术的应用是当前重要的控镉技术措施。大田试验结果(图9)显示,降镉灵、富硒宝、微量元素肥能够降低大田烟株根和叶中NtCCX2基因的表达量,减少各部位叶和根的Cd 含量,其中降镉灵降低下部叶Cd 含量的效果比较明显,进一步表明NtCCX2基因参与烟株中Cd的吸收与运输。
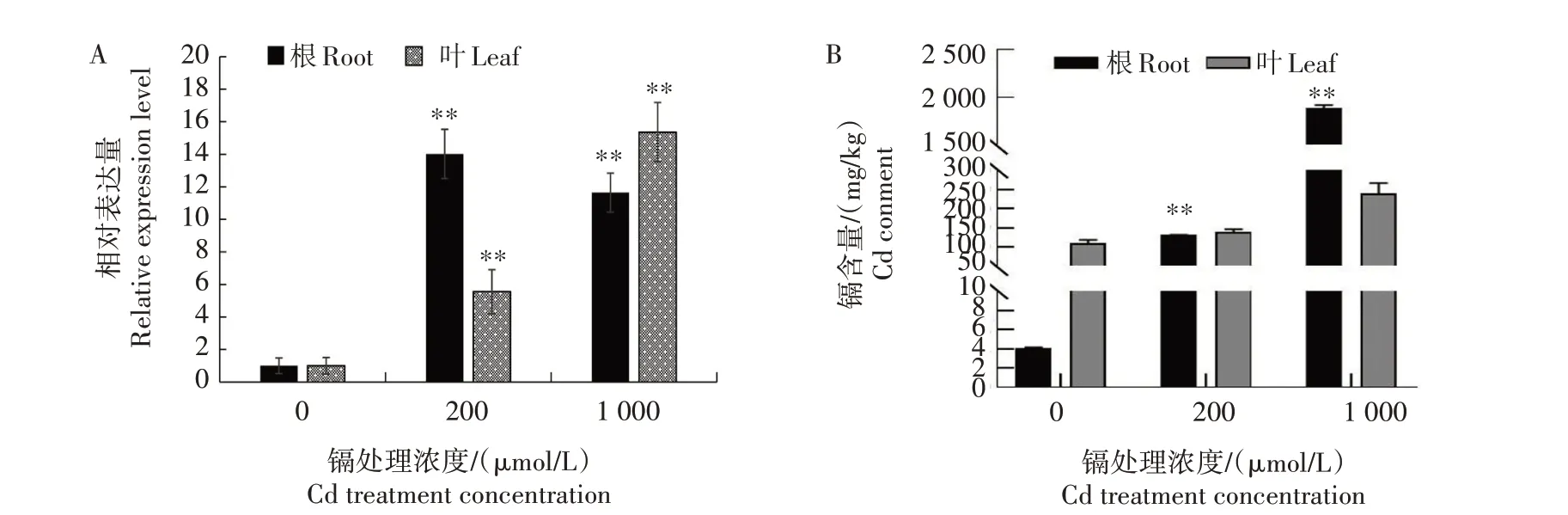
图8 Cd胁迫对NtCCX2基因表达(A)与烟株Cd含量(B)的影响Fig.8 Effects of cadmium stress on NtCCX2 gene expression(A)and cadmium content(B)in tobacco plants
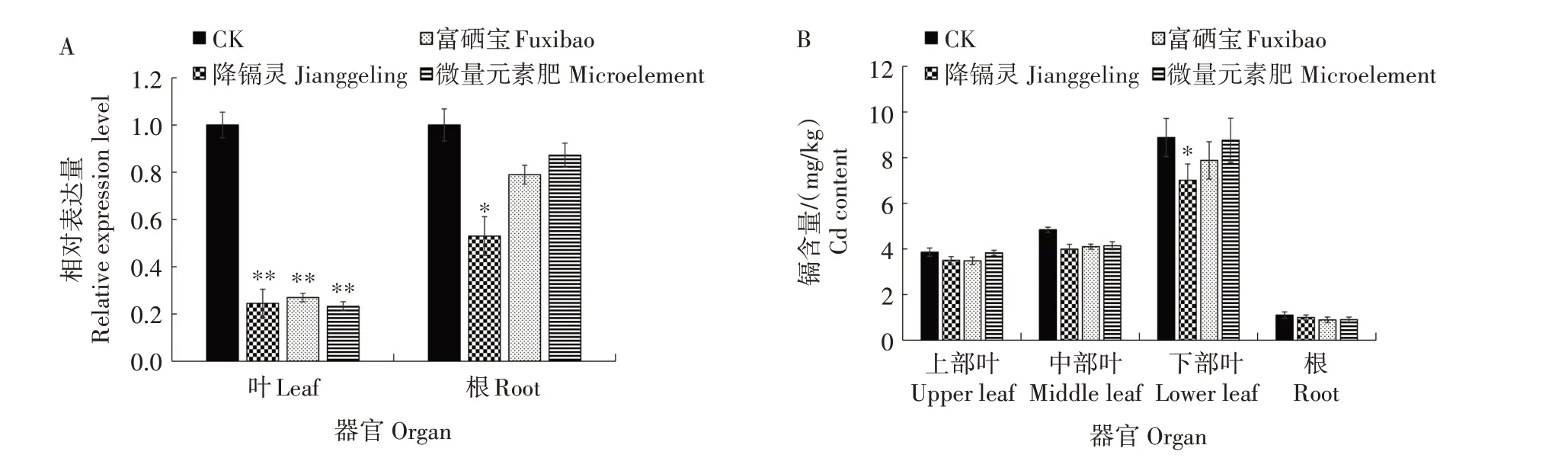
图9 降镉药剂对大田烟株NtCCX2基因表达(A)和Cd含量(B)的影响Fig.9 Effect of cadmium‑reducing agents on expression of NtCCX2 gene(A)and cadmium content(B)in tobacco plants
3 结论与讨论
Ca2+/阳离子反转运蛋白超家族(CaCA)是一类膜定位的离子转运蛋白,主要包含YRBG、CAX、CCX、NCX和NCKX 5个亚家族成员,CAX和CCX普遍存在于植物中,在植物非生物胁迫响应中起到重要作用[22‐24]。CCX一般有CCX1—CCX5 5个成员,跨膜区是包含2 个α-重复区域的12 个区段,保守的α1 模式基序GNGAPD 和α2 模式基序G(N/D)SxGD分别存在于N-端的第3—4 区段和C-端的第9—10区段[25],本研究结果表明,NtCCX2具有这种典型的保守结构。
CCX2 参与体内的Na+、K+、Mn2+、Ca2+等的运输。拟南芥中的CCX 家族具有H+/K+转运活性,通过调控Ca2+浓度,AtCCX1、AtCCX2、AtCCX3参与干旱、盐胁迫响应,AtCCX1参与叶片的衰老过程[26]。研究发现,小麦TaCCXs、水稻OsCCX2和OsCCX4的表达受干旱胁迫诱导,而OsCCX2还与Cd 的运输有关[27‐29]。本研究结果显示,NtCCX2基因启动子含有ABA 等激素信号以及抗逆相关的响应元件,其在根和叶中的表达除了受到干旱和盐胁迫的诱导外,还对ABA、MeJA 和乙烯利产生不同的响应,Ca2+等离子的调控是否也参与这些响应过程尚需进一步研究。另外,本研究结果还表明,Cd 胁迫能显著增加NtCCX2基因的表达,降镉灵等降镉药剂能够下调该基因的表达,不仅表明NtCCX2与烟草中Cd 的吸收、累积有关,同时也为降镉药剂的推广使用提供了理论基础。
