半干旱区河岸带灌木镰叶锦鸡儿种子萌发特征
2023-06-11王高峰韩大勇吴晨阳木依丁吾布力卡司木
王高峰 韩大勇 吴晨阳 木依丁?吾布力卡司木
摘要 通过室内模拟试验,探讨了土壤水分、凋落物覆盖和种子大小对镰叶锦鸡儿种子萌发的影响。镰叶锦鸡儿种子萌发主要受土壤含水量制约,在适宜的水分条件(8%含水量)下萌发迅速、整齐,发芽率达到80%~100%,而在低含水量(2%)时发芽率仅20%~40%;其次是凋落物,但凋落物的作用受土壤含水量的影响,在2%含水量条件下,凋落物具有明显的抑制作用,延长萌发天数,降低发芽率,但是在8%含水量下凋落物无明显作用;种子大小未明显影响发芽率。镰叶锦鸡儿具有一次、大量萌发的萌发对策,发芽率很高,在萌发阶段不存在限制种群更新问题,未来应从镰叶锦鸡儿种群种子散布以及微生境对幼苗存活影响等方面开展进一步研究。
关键词 种群更新;土壤水分;凋落物;种子大小;伊犁河谷
中图分类号 S722 文献标识码 A
文章编号 0517-6611(2023)10-0093-05
doi:10.3969/j.issn.0517-6611.2023.10.020
Abstract The effects of soil moisture,litter cover and seed size on seed germination of Caragana aurantiaca were studied through manipulated simulation experiments.The seed germination of Caragana aurantiaca is mainly limited by soil water content.It germinates rapidly and neatly under appropriate water conditions (8% water content),and the germination rate reaches 80%-100%,while at low water content (2%),the germination rate is only 20%-40%; The second factor is litter,but the effect of litter is affected by soil water content.Under the condition of 2% water content,litter has obvious inhibitory effect,prolonging germination days and reducing germination rate,but litter has no obvious effect under 8% water content; Seed size did not significantly affect the germination rate.Caragana aurantiaca has the germination strategy of onetime and largescale germination,and the germination rate is very high.There is no problem of limiting population regeneration in the germination stage.In the future,further research should be carried out from the aspects of seed dispersion of Caragana aurantiaca population and the impact of microhabitat on seedling survival.
Key words Population regeneration;Soil moisture;Litter;Seed size; Ili River Valley
种子萌发是影响植物种群更新的关键过程之一,受多种因素的综合影响,尤其与土壤水分状况密切相关[1]。种子萌发对土壤水分的反映可以表现在萌发需水量和萌发率2个方面。一般而言,干旱区的植物萌发需水量较低,如生活于干旱草原的无芒隐子草种子萌发的适宜土壤含水量为6%[2],旱生植物梭梭[3]的种子发芽要求含水量至少8%,泡果沙拐枣种子萌发的启动含水量为1%即可[4],而生活于湿润环境的地枫皮[5]、坡垒[6]种子萌发的适宜含水量分别高达40%和70%。就萌发率而言,生活于干旱区的植物种子往往具有休眠特性,在萌发特征上表现为萌发率低、萌发不整齐等,如沙生植物月见草[7]、沙米[8]以及其他沙漠植物[9-11],这种萌发方式也被称为两头下注策略[12-13],是降低远期风险的表现。因此,种子萌发过程中无论是需水量还是萌发率,都是长期适应当地环境条件(如降水量)的结果,只有在进化的背景下才能更深刻地理解植物种子萌发的水分需求特征。
凋落物通常改变地表覆盖情况和土壤水热条件,进而影响种子萌发。以往研究表明,凋落物对植物种子萌发存在促进作用[14]、抑制作用[15]和中性作用[16-17]等多种效应。其原因在于凋落物的作用受环境制约,如凋落物对黧蒴锥种子萌发的作用与土壤湿润程度有关[18]。目前,关于凋落物的作用在不同土壤水分条件下发生何种变化尚无统一结论,需要在不同水分条件下开展多因素对比研究。
除了上述生物与非生物环境因素外,种子自身性状也是重要的影响方面。种子大小是种子的核心功能性状之一[19],关乎种子发芽率和幼苗存活。从早期研究来看,有研究支持大、小種子的萌发有差异[20],也有研究支持大、小种子的萌发没有差异[16,21],具体与环境胁迫程度有关。如在干旱胁迫下侧柏[22]种子或者盐胁迫下盐角草[23]种子萌发均表现为在高胁迫下大种子更有萌发优势,但是在低胁迫环境时无差别或者差别变小,说明环境胁迫越强,大种子的萌发优势越明显,抗逆性也更强。因此,对比研究不同环境条件下种子大小对萌发过程的影响对于了解特定植物种群的更新具有重要意义。
镰叶锦鸡儿是分布于伊犁特克斯河河岸带的一种灌木,是河岸带植被的建群种之一,存在天然更新不良的问题。以往为了了解影响种群更新障碍的原因,韩大勇等[24-26]报道了镰叶锦鸡儿的幼株种群年龄结构、植物多样性、结实特征等,但是关于其种子萌发特征尚未涉及。尽管有研究探讨了盐胁迫和温度对镰叶锦鸡儿种子萌发的影响[27],但是这2个因素在天然种群更新中不是主要问题。事实上,镰叶锦鸡儿群落的下层草本植物甚为发达,常形成致密的凋落物层,这对镰叶锦鸡儿种子萌发有何影响,以及如何影响种群更新尚不清楚。因此,了解土壤微生境特征对镰叶锦鸡儿种子萌发过程的影响既是迫切需要了解的理论问题,也是生产实际中需要解决的问题。笔者通过开展室内模拟试验,初步探讨了土壤水分、凋落物覆盖和种子大小对种子萌发的影响,旨在进一步了解镰叶锦鸡儿种群更新,也为河岸带植被恢复提供参考依据。
1 材料与方法
1.1 研究区概况 伊犁河谷位于新疆北部(80°9′42″~91°01′45″E,40°14′16″~49°10′45″N) ,行政区划隶属伊犁哈萨克自治州。地貌上伊犁河谷为天山山脉山间盆地,东、南、北三面环山,形成向西敞开的喇叭形谷地[24]。特殊的地形使得北冰洋的寒冷气流和塔克拉玛干沙漠的干热风难以到达,而来自西面的温暖气流可以顺利进入。因此,伊犁河谷形成了特殊气候环境,具有典型的干旱气候特征,属寒温带半干旱的大陆性气候类型。研究地点位于昭苏县解放桥湿地,为镰叶锦鸡儿集中分布区域。采样区为阶地生境,地势较高,河床下切 1.0~1.5 m,仅洪水期能够接受河水补给,地表较干,镰叶锦鸡儿发育良好,灌叢密度平均1.09丛/m2[26]。
1.2 研究方法
1.2.1 试验材料。
2021年8月中旬在昭苏县解放桥湿地采集健康、成熟镰叶锦鸡儿种子,采样区生境类型为阶地,管理方式为围封管理,镰叶锦鸡儿灌丛基本特征见文献[26]。凋落物取自昭苏镰叶锦鸡儿样地,为短柱苔草(Carex turkestanica)、看麦娘(Alopecurus aequalis)等单子叶植物当年死亡残体。室内85 ℃烘干,紫外线照射杀菌后,剪成1 cm左右的短段备用。
1.2.2 种子吸胀特征。
试验前抽取50粒成熟饱满的镰叶锦鸡儿种子并称重,置于温水(50 ℃)中浸泡48 h,每隔6 h观察1次种子吸胀情况,称重并做记录。重复3组。试验未区分大种子和小种子。
1.2.3 萌发试验。
2021年10月进行室内萌发试验,设置土壤含水量、凋落物覆盖和种子大小3个因素。其中土壤含水量设2%和8% 2个水平,凋落物覆盖设0、4和8 g 3个水平(按基质质量比为0、3.3%和6.6%),种子大小分为大种子(种子百粒重≥0.38 g)和小种子(种子百粒重<0.38 g)2个水平。采用完全随机区组设计,12个处理,3次重复。具体见表1。
萌发条件为恒温25 ℃,14 h光照,10 h黑暗。采用培养皿进行萌发,基质为细沙,每个培养皿放细沙120 g,细沙事先105 ℃烘干、消毒,时间为2 h。处理好之后,在每个培养皿中放置镰叶锦鸡儿种子20粒,播种深度1 cm。凋落物直接覆盖在沙子表面。每天记录萌发情况。萌发标准:胚根突破种皮露白。当连续3 d不再有种子萌发时结束试验,总计培养15 d。
1.3 数据分析
1.3.1 发芽指标。
采用发芽率、发芽势、萌发历期、开始萌发天数等指标从不同方面反映镰叶锦鸡儿种子萌发特征。发芽率=最终萌发种子数/全部种子数×100%;发芽势为5 d的发芽率;萌发历期是达到最大萌发数所经的天数;开始萌发天数是种子开始萌发所需天数。
1.3.2 数据统计。
采用多重比较,对不同处理的种子萌发指标(发芽率、发芽势、萌发历期、开始萌发天数)进行差异显著性分析(采用Duncan方法);采用单因素方差分析研究各因素对各发芽指标的影响及其交互效应;基于线性回归模型,对各因素对种子萌发影响的相对大小进行了权重分析。所有统计分析显著水平P=0.05。数据分析统计处理用SPSS软件(SPSS Inc,Chicago,IL)和R语言软件(R Development Core Team 2015)完成。
2 结果与分析
2.1 种子吸胀特征
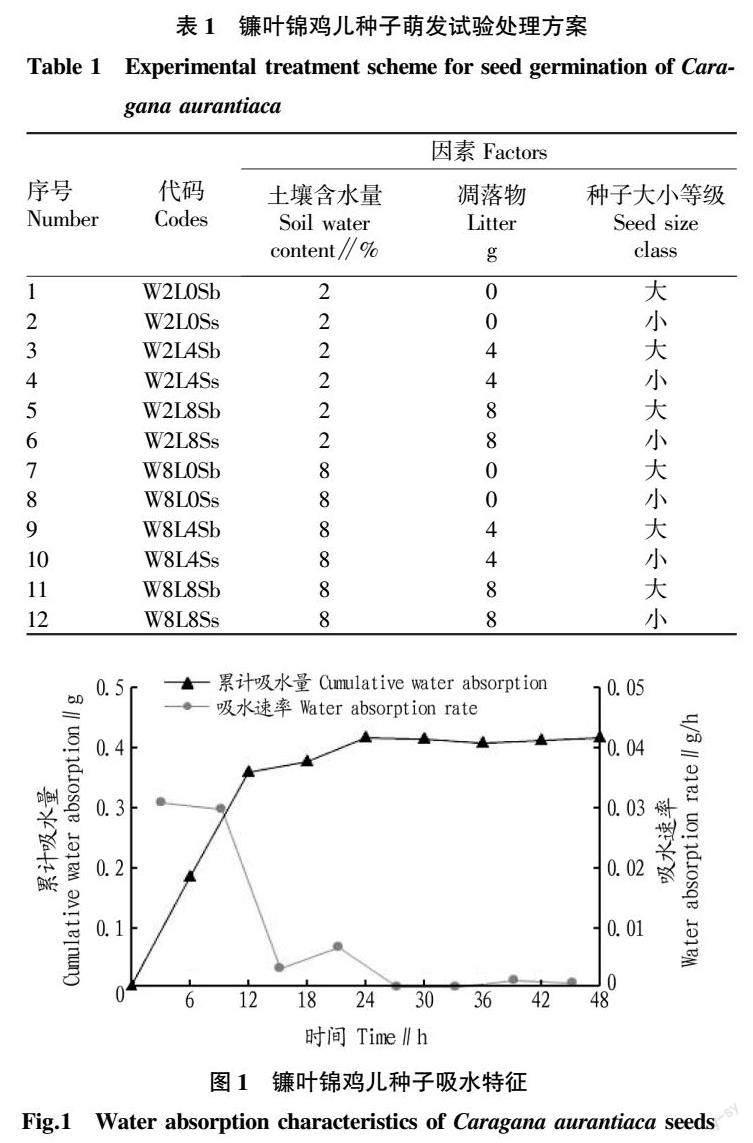
根据镰叶锦鸡儿种子的吸水特征(图1),可以大致划分出3个阶段:0~12 h为镰叶锦鸡儿种子快速吸水期,吸水量平均0.38 g,吸水速率为0.031 3 g/h,占自身种子重量的87.3%;12~24 h为缓慢吸水期,吸水量为0.04 g,吸水速率为0.006 7 g/h,占自身种子重量的9.27%;24 h之后为饱和期,种子重量基本保持稳定,最终吸水量为0.415 4 g。
2.2 种子发芽特征
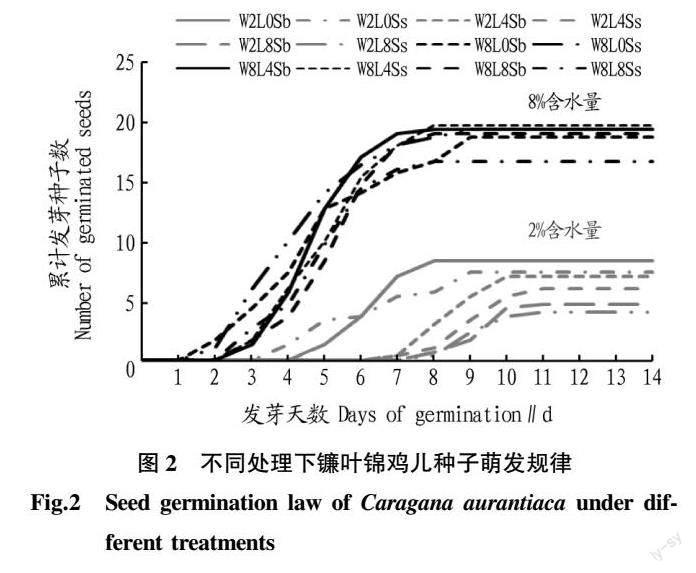
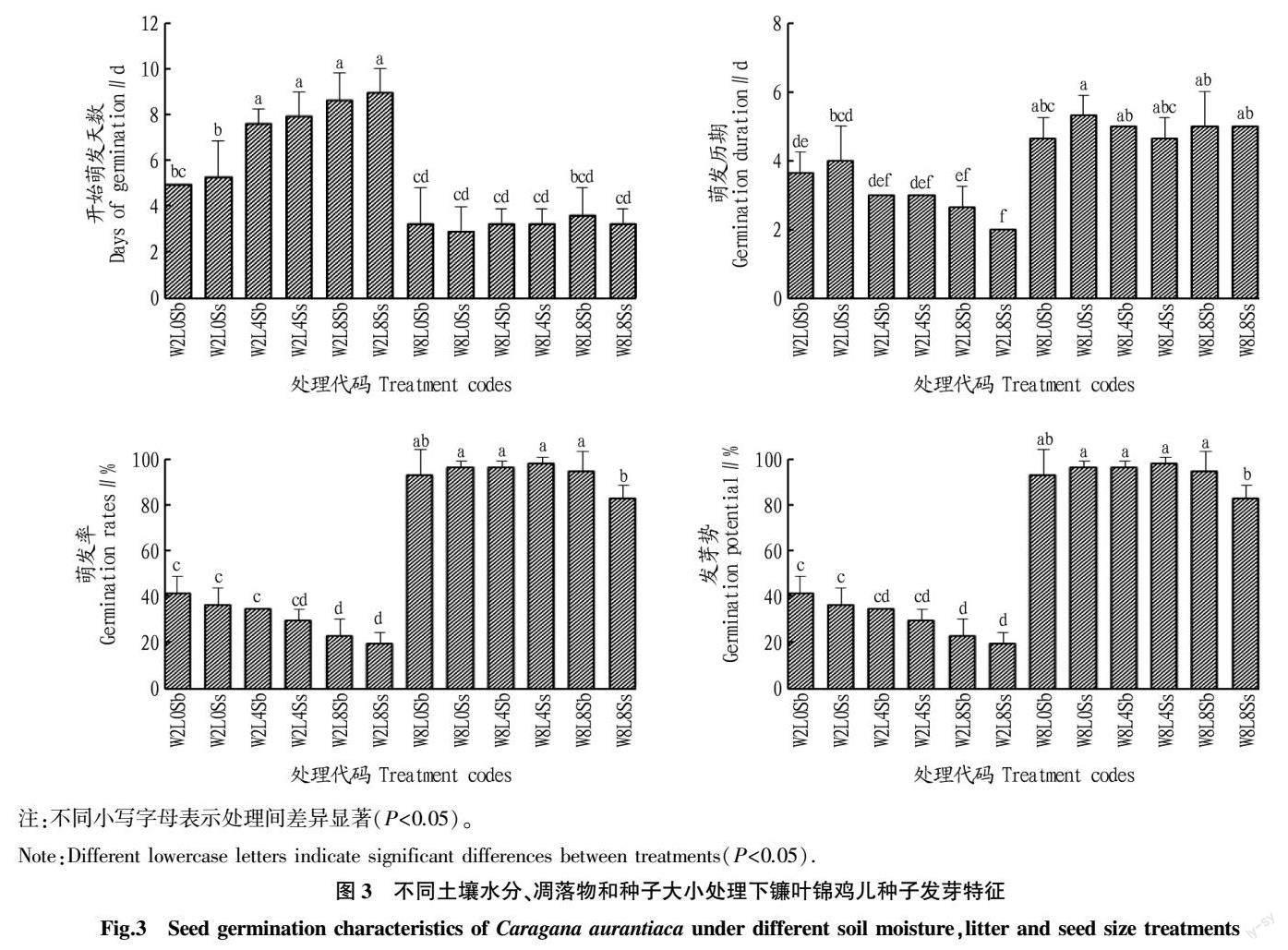
在基质含水量8%的条件下,各处理镰叶锦鸡儿种子萌发过程基本一致,在播种后第2天开始萌发,且萌发整齐,发芽势高,尽管萌发历期要高于2%水分条件,但是最终的发芽率更高,为80%~100%,而2%水分条件下的发芽率仅20%~40%(图2、3)。方差分析结果表明,基质含水量和凋落物对镰叶锦鸡儿种子发芽均有显著影响(P<0.05),种子大小的影响并不显著(P>0.05),各因素之间无显著交互作用(P>0.05)(表2)。但从多重比较结果看,凋落物与含水量有关,在8%含水量下作用不明显,在2%含水量下有显著的负面影响(P<0.05)(图3)。权重分析表明,土壤含水量对各萌发指标的解释率在85.42%~97.50%,凋落物解释率在1.97%~14.52%,种子大小解释率极低,均不足1.00%(表3)。
3 讨论
在自然选择作用下,植物种子萌发存在2个进化方向,既有大量、一次萌发对策,也有少量、多次萌发对策,但是不论采取哪种萌发对策,目的都是提高种群适合度[28]。根据最优萌发理论,在波动环境下(主要是干旱),植物种子萌发采取两头下注的方式,既要保障一定数量的种子萌发,又要避免大量萌发后幼苗死亡的问题[12]。因此,干旱区的植物一般具有少量、多次萌发的特征,这是对降水特征(一次偶然降水事件后往往出现长期干旱)的反映[29]。反之,如果不存在种子大量萌发后幼苗死亡的远期风险,则大量、一次萌发对种群适合度是更有利的。可见,具体采取哪种萌发对策与干旱的频率、强度以及持续时间有关,确切地说与当地降水特征有关。该研究区所在的伊犁河谷是整个新疆干旱区最湿润的区域之一,伊犁河东西向贯穿河谷,镰叶锦鸡儿分布于伊犁河主源支流特克斯河两岸,水分条件优越,平原区降水量普遍在400 mm,山区能达到800~1 000 mm。根据其分布区的降水事件,4—8月出现干旱的频率不足10%,干旱主要出现在非生长季[30]。显然,镰叶锦鸡儿分布区的水分条件相对于荒漠而言是相当优越的,不存在萌发后遇到长期干旱的情况。因此,作为分布于河岸带隐域生境的物种,镰叶锦鸡儿未表现出干旱区植物种子萌发的两头下注方式,而是表现出在适宜水分条件下迅速吸水并大量、一次萌发的特征。但是,镰叶锦鸡儿种子萌发需水量具备干旱区植物的一般特征,即萌发需水量较低。该研究中,8%含水量就已经满足其萌发需求,发芽率最高可达100%,这与同样分布于河岸带的多刺锦鸡儿的需水特征接近[31]。
根据植物地理学的研究成果,镰叶锦鸡儿是锦鸡儿属植物因青藏高原抬升而在中亚地区向旱生方向演化的种类[32-33],特征是叶片小、复叶假掌状。植物的適应固然是因素驱动的结果,种子萌发策略难以表达出全部内涵,但也携带着演化趋势及其与环境关系的丰富信息。该研究中镰叶锦鸡儿种子萌发较低的需水量验证了其旱生演化特点,但是,其大量、一次萌发策略则是长期适应河岸带生活的结果。
凋落物在该研究中也表现出对镰叶锦鸡儿种子萌发的显著影响,可降低发芽率,这与以往的部分研究结果一致[15]。同时还发现,凋落物的作用程度明显受到土壤水分的制约,在干旱条件(2%含水量)下,凋落物具有明显的抑制作用,延长萌发天数,降低发芽率,但是在水分充足(8%含水量)时,作用不明显,在几个处理间差异不显著。凋落物对种子萌发的影响有以下途径:①化感作用[34-35];②机械隔离[36];③遮阴[37];④增加幼苗感染真菌的机会[14]。该研究认为,化感作用是最可能的途径之一,原因是:机械隔离往往对大种子影响大,如壳斗科植物种子[36,38];该研究中采取沙埋发芽,凋落物覆盖在沙子上面,可以排除遮光的作用;至试验结束,无腐烂种子,未发生真菌感染导致种子失去活力现象。另外,在8%含水量条件下,凋落物的作用不显著,可能是因为水分含量高稀释了化感物质的浓度,降低其抑制作用,这也是一个佐证,需要后续开展试验进一步验证。但该研究至少可以给出部分启示,凋落物层既可能通过保水作用有利于种子萌发,也可能因为化感作用而抑制种子萌发。若以覆盖原地凋落物的方式促进种群更新,应充分考虑凋落物的化感效应。
以往研究表明,大种子在胁迫生境下发芽率更高[22-23],但该研究中,无论在哪种含水量处理下,镰叶锦鸡儿大种子和小种子的发芽率均无显著差异,大种子未表现出萌发优势。该研究认为,这与镰叶锦鸡儿的萌发对策有关,如果小种子萌发不齐,意味着出现延迟萌发,因此,不论大种子还是小种子,只要能保障存活、萌发即可,并不一定非要产生大种子。这种现象在更多的旱生植物中都有发现[21,39],包括沙鞭[40]、沙蒿[41]、梭梭[3]、华北驼绒藜[42]、柠条锦鸡儿[43]等,这些旱生植物的大种子和小种子萌发率均无差异,种子大小不构成选择压力,可能是趋同适应的结果。
4 结论
种群更新限制发生于各个生活史阶段,受到多个因素的制约,包括种源限制、扩散限制、微生境限制等[1]。以往研究证实,镰叶锦鸡儿种群不存在种源限制问题[26],该研究则表明,种子无明显休眠现象,种子萌发主要受土壤含水量制约,且影响最大,其次是凋落物,种子大小无影响。在合适水分条件下发芽率高,达到80%~100%,且萌发迅速、整齐,在萌发阶段不存在限制种群更新问题。镰叶锦鸡儿分布的河岸带土壤含水量4.63%~6.98%[25],是种子萌发的合适水分,但是实生苗种群年龄结构是衰退型的[24]。因此,镰叶锦鸡儿种群更新限制主要发生在扩散和幼苗这2个生活史过程,未来应从镰叶锦鸡儿种群种子散布以及微生境对幼苗存活影响等方面开展进一步研究。
参考文献
[1]韩大勇,张维,努尔买买提·依力亚斯,等.植物种群更新的补充限制[J].植物生态学报,2021,45(1):1-12.
[2] 邰建辉,王彦荣,陈谷.无芒隐子草种子萌发、出苗和幼苗生长对土壤水分的响应[J].草业学报,2008,17(3):105-110.
[3] 朱淑娟,王方琳,刘有军,等.土壤水分和种子大小对梭梭种子萌发、出苗和幼苗生长的影响[J].西北林学院学报,2021,36(6):16-21.
[4] 朱成林,李晓梅,阿依帕夏·居麦,等.泡果沙拐枣种子萌发对模拟降雨间隔的响应[J].防护林科技,2020(9):1-6.
[5] 文彬,兰芹英,何惠英.光、温度和土壤水分对坡垒种子萌发的影响[J].热带亚热带植物学报,2002,10(3):258-262.
[6] 韩愈,韦霄,唐辉,等.土壤水分对地枫皮种子萌发与早期幼苗生长的影响[J].种子,2018,37(7):10-15.
[7] EVANS M E K,FERRIRE R,KANE M J,et al.Bet hedging via seed banking in desert evening primroses (Oenothera,Onagraceae):Demographic evidence from natural populations[J].The American naturalist,2007,169(2):184-194.
[8] 王宗灵,徐雨清,王刚.沙区有限降水制约下一年生植物种子萌发与生存对策研究[J].兰州大学学报(自然科学版),1998,34(2):98-103.
[9] VENABLE D L.Bet hedging in a guild of desert annuals[J].Ecology,2007,88(5):1086-1090.
[10] 王彦荣,杨磊,胡小文.埋藏条件下3种干旱荒漠植物的种子休眠释放和土壤种子库[J].植物生态学报,2012,36(8):774-780.
[11] GREMER J R,KIMBALL S,VENABLE D L.Withinand amongyear germination in Sonoran Desert winter annuals:Bet hedging and predictive germination in a variable environment[J].Ecology letters,2016,19(10):1209-1218.
[12] COHEN D.Optimizing reproduction in a randomly varying environment[J].Journal of theoretical biology,1966,12(1):119-129.
[13] SANCHEZMARTIN R,GOMEZ J M,CHEPTOU P O,等.两型结实豆科植物种子休眠和萌发的差异:空间和时间上多样性的两头下注策略[J].植物生态学报:英文版,2021,14(4):662-672.
[14] HU J J,LUO C C,TURKINGTON R,et al.Effects of herbivores and litter on Lithocarpus hancei seed germination and seedling survival in the understorey of a high diversity forest in SW China[J].Plant ecology,2016,217(1):1429-1440.
[15] MOLOFSKY J,LANZA J,CRONE E E.Plant litter feedback and population dynamics in an annual plant,Cardamine pensylvanica[J].Oecologia,2000,124(4):522-528.
[16] GREEN P T.Seed germination in Chrysophyllum sp.nov.,a largeseeded rainforest species in north Queensland:Effects of seed size,litter depth and seed position[J].Australian journal of ecology,1999,24(6):608-613.
[17] LOYDI A,ECKSTEIN R L,OTTE A,et al.Effects of litter on seedling establishment in natural and seminatural grasslands:A metaanalysis[J].Journal of ecology,2013,101(2):454-464.
[18] 王俊,王卓晗,杨龙,等.浇水频率和凋落物覆盖量对黧蒴锥种子萌发及幼苗存活的影响[J].应用生态学报,2008,19(10):2097-2102.
[19] 張红香,周道玮.种子生态学研究现状[J].草业科学,2016,33(11):2221-2236.
[20] 王慧慧,王普昶,赵钢,等.干旱胁迫下白刺花种子大小与萌发对策[J].生态学报,2016,36(2):335-341.
[21] 王桔红,崔现亮,陈学林,等.中、旱生植物萌发特性及其与种子大小关系的比较[J].植物生态学报,2007,31(6):1037-1045.
[22] 付锦雪,张晨星,王晓霞,等.水分胁迫和种子大小对侧柏种子萌发的影响[J].种子,2020,39(2):50-57.
[23] AMEIXA O M C C,MARQUES B,FERNANDES V S,et al.Dimorphic seeds of Salicornia ramosissima display contrasting germination responses under different salinities[J].Ecological engineering,2016,87:120-123.
[24] 韩大勇,赵玉,贾娜尔·阿汗,等.新疆伊犁河谷镰叶锦鸡儿(Caragana aurantiaca)幼株种群年龄结构[J].生态学杂志,2019,38(12):3601-3608.
[25] HAN D Y,WANG J F,AHAN J,et al.Soil and shrub differentially determine understorey herbaceous plant richness and abundance in a semiarid riparian meadow[J].Community ecology,2021,22(1):113-125.
[26] 韩大勇,胡鹏飞,张维,等.伊犁特克斯河河岸带镰叶锦鸡儿灌丛分株结荚特征[J].生态学杂志,2021,40(3):740-748.
[27] 苗昊翠,刘博,邓江宇,等.新疆两种锦鸡儿种子萌发特性研究[J].新疆农业科学,2009,46(4):810-817.
[28] 李良,王刚.种子萌发对策:理论与实验[J].生态学报,2003,23(6):1165-1174.
[29] CLAUSS M J,VENABLE D L.Seed germination in desert annuals:An empirical test of adaptive bet hedging[J].The American naturalist,2000,155(2):168-186.
[30] 宋蕾.基于M-K法和小波分析法的昭苏县四季降水变化特征分析[J].水利科学与寒区工程,2020,3(6):35-39.
[31] 王璐瑶,张云玲,赵晓英,等.河流湿地灌木多刺锦鸡儿的种子萌发特性[J].中国草地学报,2020,42(4):35-41.
[32] 周道玮,刘钟龄,马毓泉.豆科锦鸡儿属(Caragana Fabr.)植物地理分布与分化研究[J].植物研究,2005,25(4):471-487.
[33] 李惠茹,杜诚.中国锦鸡儿属植物地理分布及价值研究[J].园艺与种苗,2013,33(5):63-67.
[34] HALVORSON J J,BELESKY D P,WEST M S.Inhibition of forage seed germination by leaf litter extracts of overstory hardwoods used in silvopastoral systems[J].Agroforestry systems,2017,91(1):69-83.
[35] 马冬雪,刘仁林.天然群落枯枝落叶浸提液与其它处理对伯乐树种子发芽的比较研究[J].林业科学研究,2012,25(5):632-637.
[36] 朱静,刘金福,何中声,等.凋落物物理阻隔对格氏栲种子萌发及胚根生长的影响[J].生态学报,2020,40(16):5630-5637.
[37] 周艳,陈训,韋小丽,等.凋落物对迷人杜鹃幼苗更新和种子萌发的影响[J].林业科学,2015,51(3):65-74.
[38] 张建国,黄财智,陈永华,等.枯落物对辽东栎种子萌发及生长的影响[J].西北农林科技大学学报(自然科学版),2018,46(6):59-64.
[39] 张玲卫,刘会良,盛建东.埋藏对10种短命植物种子萌发特性的影响[J].干旱区研究,2018,35(3):633-639.
[40] 朱雅娟,董鸣,黄振英.沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响[J].植物生态学报,2005,29(5):730-739.
[41] 王方琳,柴成武,尉秋实,等.沙埋和种子大小对沙蒿种子萌发及幼苗生长的影响[J].西北林学院学报,2020,35(6):129-134.
[42] 潘慧超,王俊锋,敖云娜,等.干旱胁迫下华北驼绒藜种子大小及苞片对萌发和幼苗生长的影响[J].应用生态学报,2021,32(2):399-405.
[43] 杨慧玲,梁振雷,朱选伟,等.沙埋和种子大小对柠条锦鸡儿种子萌发、出苗和幼苗生长的影响[J].生态学报,2012,32(24):7757-7763.
