白菜型油菜开花调控基因BrFT的生物信息学特性和表达分析
2023-06-09姚彦林刘丽君蒲媛媛李学才王旺田孙万仓武军艳
姚彦林, 马 骊, 刘丽君, 蒲媛媛, 李学才, 王旺田,方 彦 , 孙万仓,武军艳,*
(1.甘肃农业大学 农学院/甘肃省油菜工程技术研究中心,甘肃 兰州 730070; 2.省部共建干旱生境作物学国家重点实验室,甘肃 兰州 730070; 3.甘肃农业大学 生命科学技术学院,甘肃 兰州 730070)
开花是植物由营养生长转向生殖生长的重要阶段[1]。植物开花不但受外界环境的影响还受多基因调控。近些年随着分子生物技术的迅速发展,通过对一些模式植物拟南芥、水稻的研究,有大量与开花相关的基因被成功鉴定和分离。已有研究表明,植物成花过程存在6条途径,分别是光周期途径、春化途径、赤霉素途径、自主途径、年龄途径以及温敏途径,各途径中的信号通过开花整合因子相互联系,植物才能完成开花[2-3]。FT、SOC1是植物开花过程中重要的开花整合因子,其中FT基因是光周期途径与春化途径中各信号的整合点,在调控植物开花中发挥着重要作用[4-5]。
FT基因编码的蛋白属于磷脂酰乙醇胺结合蛋白(PEBP),前人在单子叶与双子叶植物中研究发现该蛋白对植物开花时间具有重要作用,PEBP对于株型分化也产生调控作用[6-7]。FT基因对植物开花产生促进作用最早是在拟南芥中发现的,发现FT基因在拟南芥叶片中被诱导表达,然后经韧皮部运输到芽顶端分生组织,与FD蛋白形成复合体来调控下游开花基因的表达来促进植物开花[8-10]。FT基因已在多种植物被鉴定分析、克隆并验证基因功能,例如一些园艺植物如甘蔗[11]、月季[12]、葡萄[13]、大豆[14]、柑橘[15]等,FT基因的过表达都能出现早花现象[16],但同时也有一些FT的同源基因属于晚花基因,例如龙眼[17]的DlFT1、辣椒[18]、郁金香[19]的FT同源基因都是抑制成花的基因,说明不同物种间由于成花特性的差异,FT基因的功能也有差异。
白菜型冬油菜目前是北方地区主要的越冬类新型冬季覆盖作物,具有优异的抗寒性,提高产量和品质一直是油菜研究的首要目标,而开花时间是影响产量和品质的重要因素。低温和长日照均可促使冬性植物提早开花,这不但可以有效解决与其他作物的茬口矛盾,还可以促进育种进程。FT基因作为开花调控网络中的重要整合因子,在白菜型油菜中至今未报道,本研究通过生物信息学方法,首次利用甘肃农业大学最新测序的白菜型油菜基因组数据,研究了BrFT基因的结构特征,结合BrFT基因在白菜型油菜各器官的表达情况,分析了BrFT1基因在不同白菜型油菜4 ℃,16 h光照/8 h黑暗光周期中的表达模式,来揭示BrFT1对白菜型冬油菜开花的调控作用,为进一步研究白菜型油菜开花调控的分子机制提供参考依据。
1 材料与方法
1.1 实验材料与处理
以白菜型油菜陇油6号、陇油7号(强冬性)、陇油17号、天油2号(冬性)以及天祝小油菜(春性)为实验材料。选取各品种籽粒饱满的种子,用5%的次氯酸钠消毒5 min,蒸馏水冲洗干净后播于铺有两层滤纸的培养皿中,置于室温条件下,24 h种子露白后,将天祝小油菜种植于花盆,待到始蕾期取根、茎、叶、花蕾,初花期取花并液氮速冻,之后放于-80 ℃冰箱保存备用。将天油2号、陇油17号、陇油6号和陇油7号露白后的种子放于培养箱开始4 ℃,16 h光照/8 h黑暗处理,以0 d作为对照,处理时间为20、30、40、50、60、70 d,每处理完成后,取出植物材料,一部分蒸馏水冲洗干净后用滤纸迅速吸干水分,液氮速冻保存备用,一部分种植于花盆,每处理每个品种调查植株数15株,用于观察统计开花株率。实验设计3次重复。
1.2 相关物种数据
使用的拟南芥蛋白序列下载于(http://www.arabidopsis.org/)数据库,大白菜全基因组数据下载于Ensembl网站(http://plants.ensembl.org/index.html),白菜型冬油菜基因组数据来自于甘肃农业大学最新测序结果。
1.3 白菜型油菜BrFTs基因的鉴定与分析
利用FT基因保守结构域(http://Pfam.xfam.org/family/PF01161),通过HMMER3.0软件在白菜型油菜基因组数据中提取FT蛋白序列,将获得的蛋白序列提交至SMART(https://www.omicsclass.com/article/)和NCBI CDD(https://www.omicsclass.com/article/)数据库进行验证,剔除不包含FT结构域及不完整的序列,最终确定BrFT家族成员。参照马骊等[20]方法分析各成员的蛋白理化性质。通过在线工具MEME(http://meme-suite.org/)和GSGD(http://gsds.gao-lab.org/)分别分析各成员的保守结构域、内含子、外显子结构特征。由BrFT基因的位置信息,结合染色体的名称、长度信息通过软件Mapchart绘制BrFT基因染色体分布图。运用MCScanX软件分析基因共线性关系。提取基因上游2 000 bp作为启动子区,利用在线软件PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测启动子区顺式作用元件。
1.4 RNA的提取和cDNA的合成
按照TIANGEN(天根生化科技公司)植物RNA快速提取试剂盒说明书提取植物组织总RNA,利用FastKing gDNA Dispelling RT SuperMix反转录获得cDNA。
1.5 基因的克隆与表达分析
根据BrFT基因序列设计引物(表1),进行实时荧光定量PCR反应,检测BrFT基因在天祝小油菜不同组织中的表达情况,根据表达量筛选BrFT1基因设计克隆引物(表1),以常温下陇油6号、陇油17号和天油2号叶片cDNA为模板进行PCR扩增,扩增产物经1%琼脂糖凝胶电泳检测后参照TIANGEN(天根生化科技公司)的DNA纯化回收试剂盒说明书进行纯化,纯化后的产物与PMD19-T克隆载体过夜连接,再与DH5α感受态混匀培养,筛选白斑进行菌落PCR验证,将验证正确的阳性克隆送上海生工测序。根据测序结果设计荧光定量引物(表1,其中Actin为内参基因),以不同感温性冬油菜4 ℃,16 h光照/8 h黑暗处理中植株的cDNA为模板进行PCR反应,每个处理时间3个生物学重复。通过公式2-△△CT计算基因相对表达量,利用SPSS22.0软件分析显著性,作图采用Origin2021软件。

表1 引物序列Table 1 Primer sequence
2 结果与分析
2.1 白菜型油菜BrFT基因家族成员鉴定与蛋白理化性质
通过HMM搜索在白菜型油菜基因组中共搜索到3个BrFT基因,分别将它们命名为BrFT1、BrFT2、BrFT3(表2)。BrFT基因序列长度差异不大,从528 bp到531 bp,编码的氨基酸数目在175~176 aa,分子量为19.77~19.93 ku,理论等电点为7.72~7.82,说明该家族均为碱性氨基酸。蛋白的不稳定系数反映出该家族蛋白中BrFT2和BrFT3为稳定蛋白。蛋白质的亲水性均小于0,说明BrFT均为亲水性蛋白。BrFT蛋白均由α-螺旋、β-折叠、无规则卷曲和延伸链组成,其中无规则卷曲在3条BrFT蛋白中所占比例高于其他类型,其次为延伸链、α-螺旋,β-折叠最低,由此可知,无规则卷曲是BrFT蛋白二级结构的主要组成部分。
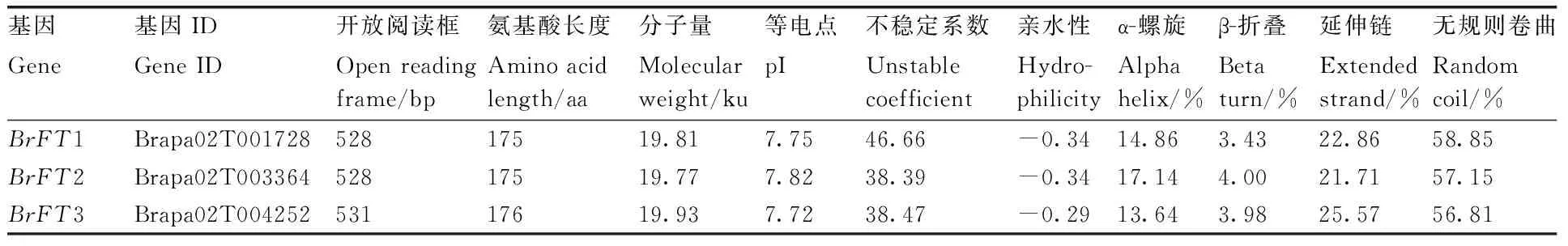
表2 BrFT基因家族成员基本信息Table 2 Basic information of BrFT gene family members
2.2 BrFT基因结构和保守基序分析
根据motif分析结果(图1),该家族3个成员均含有4个保守基序,motif1、motif2、motif3和motif4。进一步分析保守结构域发现,BrFT编码的蛋白结构域主要是磷脂酰乙醇胺结合蛋白PEBP结构域,大约由23个氨基酸组成,该结构域作为信号转导过程中的调节器来调控基因的表达。基因结构显示,该家族基因均含有4个外显子,3个内含子。BrFT1和BrFT2基因结构极度相似,BrFT3基因结构最长,长度3 500 bp左右(图2)。
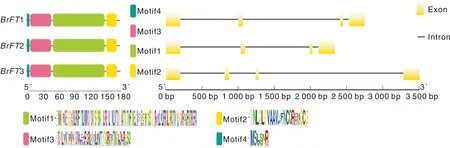
图1 BrFT基因保守结构域、基因结构图Fig.1 The conserved domain and gene structure of BrFT gene
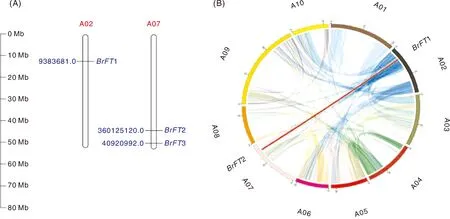
背景区域为白菜型油菜基因组中的所有共线性部分,红色线条代表存在共线性关系的BrFT基因对,不同颜色的半圈代表染色体。The background area is all the collinearity parts in B. rapa genome. The red lines represent the contributing BrFT gene pairs, and the half circles with different colors represent chromosomes.图2 BrFT基因在染色体上的位置信息及共线性关系Fig.2 Location information and collinearity of BrFT gene on chromosomes
2.3 白菜型油菜BrFT在染色体上的分布以及共线性分析
染色体定位结果显示,3个BrFT基因分别位于白菜型油菜A02、A07染色体上,其中,A02染色体上只有一个基因,其余2个基因位于A07染色体上(图2-A)。白菜型油菜组内共线性分析表明,位于A02染色体上的BrFT1与A07染色体上的BrFT2存在共线性关系,推测片段复制可能是BrFT扩增的主要因素,BrFT1和BrFT2可能在功能方面具有一定的相通性(图2-B)。
2.4 BrFT基因启动子区顺式作用元件
BrFT基因启动子区除含有启动子区和增强子区共同的顺式作用元件CAAT-box,核心启动子元件TATA-box外,主要富含4类调控元件,包括参与光反应、植物生长发育、非生物胁迫和激素诱导的相关元件(图3)。分析每个基因所含顺式作用元件,发现3个成员含有光反应的顺式作用元件最多,共38个,另外均含参与厌氧诱导、防御和应激反应的调控元件,除此之外BrFT1和BrFT2含有参与胚乳表达、生长素反应、水杨酸反应、脱落酸反应的作用元件。
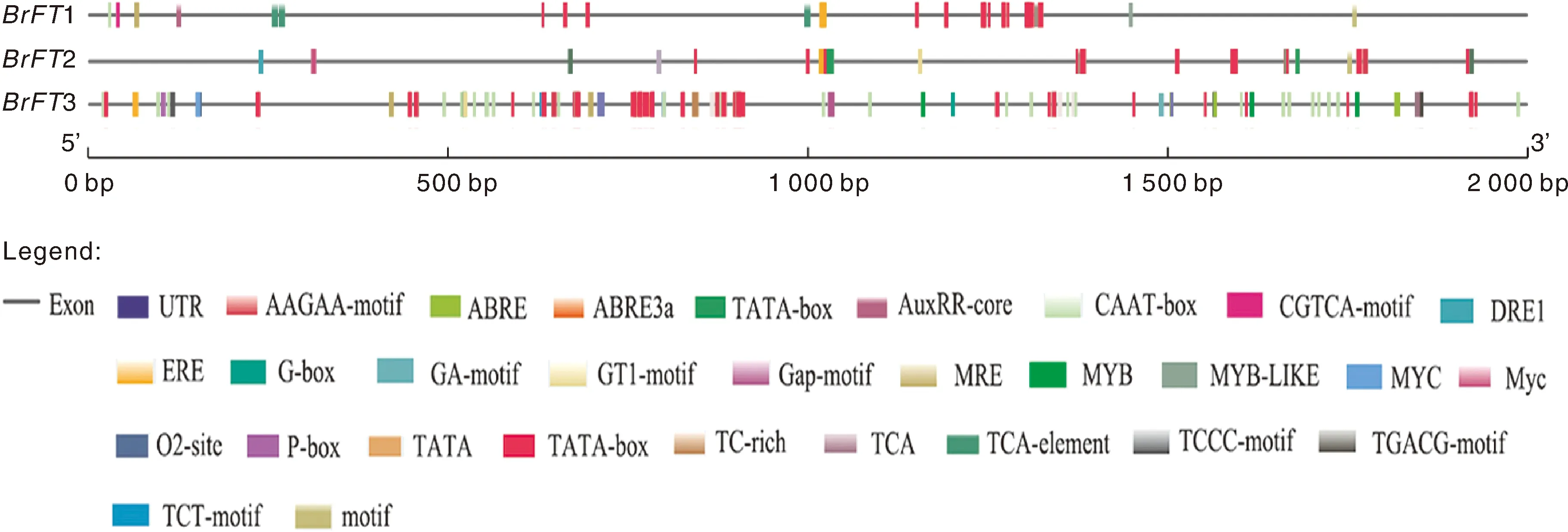
图3 BrFT基因顺式作用元件分析Fig.3 Cis-acting element analysis of BrFT gene
2.5 白菜型油菜BrFT基因在不同器官中的表达模式分析
利用课题组前期完成的陇油7号材料的转录组数据,分析BrFT基因在4 ℃,16 h光照/8 h黑暗处理30 d的表达情况,结果显示,当4 ℃,16 h光照/8 h黑暗处理30 d时,BrFT1、BrFT2、BrFT3均上调表达,其中BrFT1表达量明显高于其余两个成员(图4)。通过qRT-PCR检测BrFT在陇油7号和天祝小油菜根、茎、叶、花蕾和花各器官中的表达。结果表明,在强冬性陇油7号和春性品种天祝小油菜中,BrFT在根、茎、叶、花蕾和花各器官中均表达,其中在叶中的表达量高于其他器官部位,且BrFT1在叶中的表达量显著高于BrFT2和BrFT3(图5)。
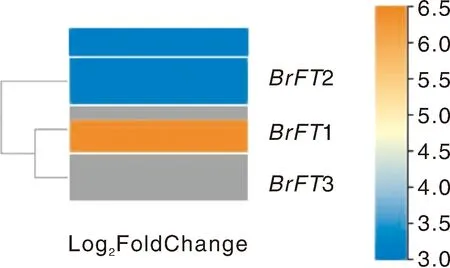
图4 陇油7号在4 ℃,16 h光照/8 h黑暗处理30 d时BrFT的表达模式Fig.4 Expression pattern of BrFT in Longyou 7 at 4 ℃, 16 h light/8 h darkness for 30 days
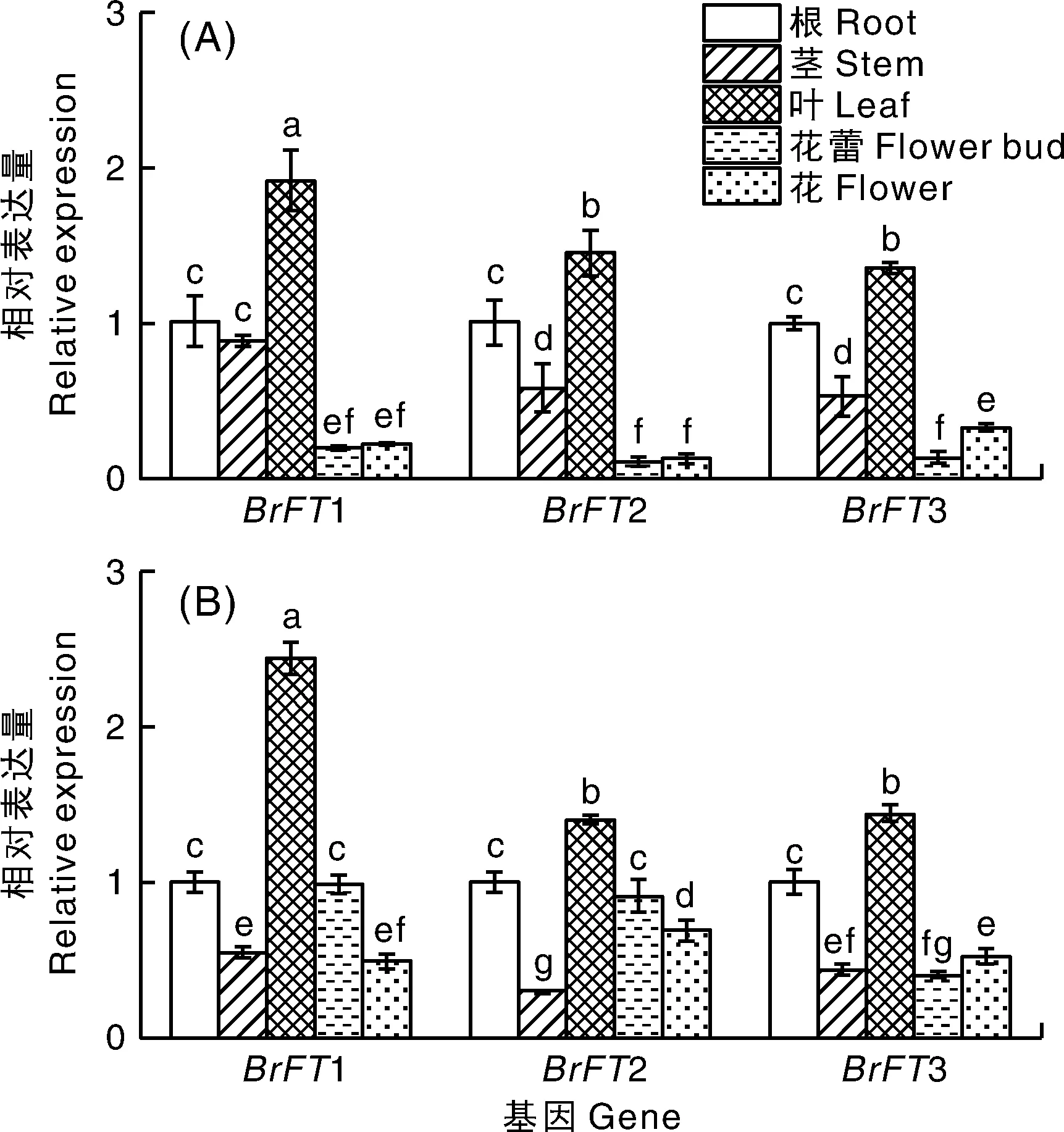
不同数据柱上没有相同小写字母的表示差异显著(P<0.05)。下同。The bars with different lowercase letters showed the significant difference (P<0.05). The same as below.图5 BrFT基因在陇油7号(A)和天祝小油菜(B)各器官中的表达Fig.5 Expression of BrFT genes in organs of Longyou 7(A) and Tianzhu small rape (B)
2.6 BrFT1基因的克隆
根据BrFT基因的表达模式,筛选表达量最高的BrFT1基因克隆,结果显示(图6),BrFT1基因CDS长528 bp,起始密码子ATG,终止密码子TAG,编码175个氨基酸。不同品种BrFT1序列比对分析表明,陇油17号中178 bp处发生了碱基突变,A突变为G,233 bp处碱基由C突变为T,与陇油7号相比,序列相似性为99.92%,陇油6号、天油2号与陇油7号相比,序列没有发生碱基突变。氨基酸序列比对结果表明,陇油17号第233 bp处发生的碱基突变并没有引起编码氨基酸的突变,而178 bp处的碱基突变致使编码的氨基酸也发生了改变,氨基酸序列相似性为99.50%(图7)。
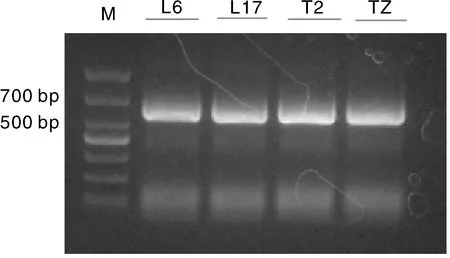
L6,陇油6号;L17,陇油17号;T2,天油2号。L6, Longyou 6; L17, Longyou 17; T2, Tianyou 2.图6 BrFT1基因扩增产物电泳图Fig.6 Electrophoretic image of BrFT1 gene amplification product

图7 不同品种白菜型冬油菜BrFT1基因氨基酸序列比对分析Fig.7 Alignment analysis of the amino acid sequence of BrFT1 gene in different varieties of winter rapeseed
2.7 BrFT1基因的表达分析
检测BrFT1基因在不同感温性白菜型冬油菜4 ℃,16 h光照/8 h黑暗处理下不同阶段的表达情况,表明该基因整体呈现上调表达(图8),这与2.5节中BrFT1基因在陇油7号中的表达模式相近。但在相同处理时间下不同品种中BrFT1基因表达不尽相同,当处理30 d时,天油2号中BrFT1的表达量高于其他各品种,结合对开花株率的统计,发现处理30 d,天油2号开花株率在各品种间最高(表3)。处理40、60 d时,BrFT1分别在天油2号和陇油17号中表达量极显著升高,发现天油2号在处理40 d的时候开花株率为98.3%,基本全部开花,而陇油17号处理60 d的时候开花株率为93.3%,开花植株数相比其他处理时间增多。而在强冬性陇油6号和陇油7号中,当处理70 d时,BrFT1基因表达量达到最大,处理70 d时,陇油6号和陇油7号开花株率明显升高,开花植株数最多。发现BrFT1的过量表达能够引起不同感温性白菜型冬油菜开花。
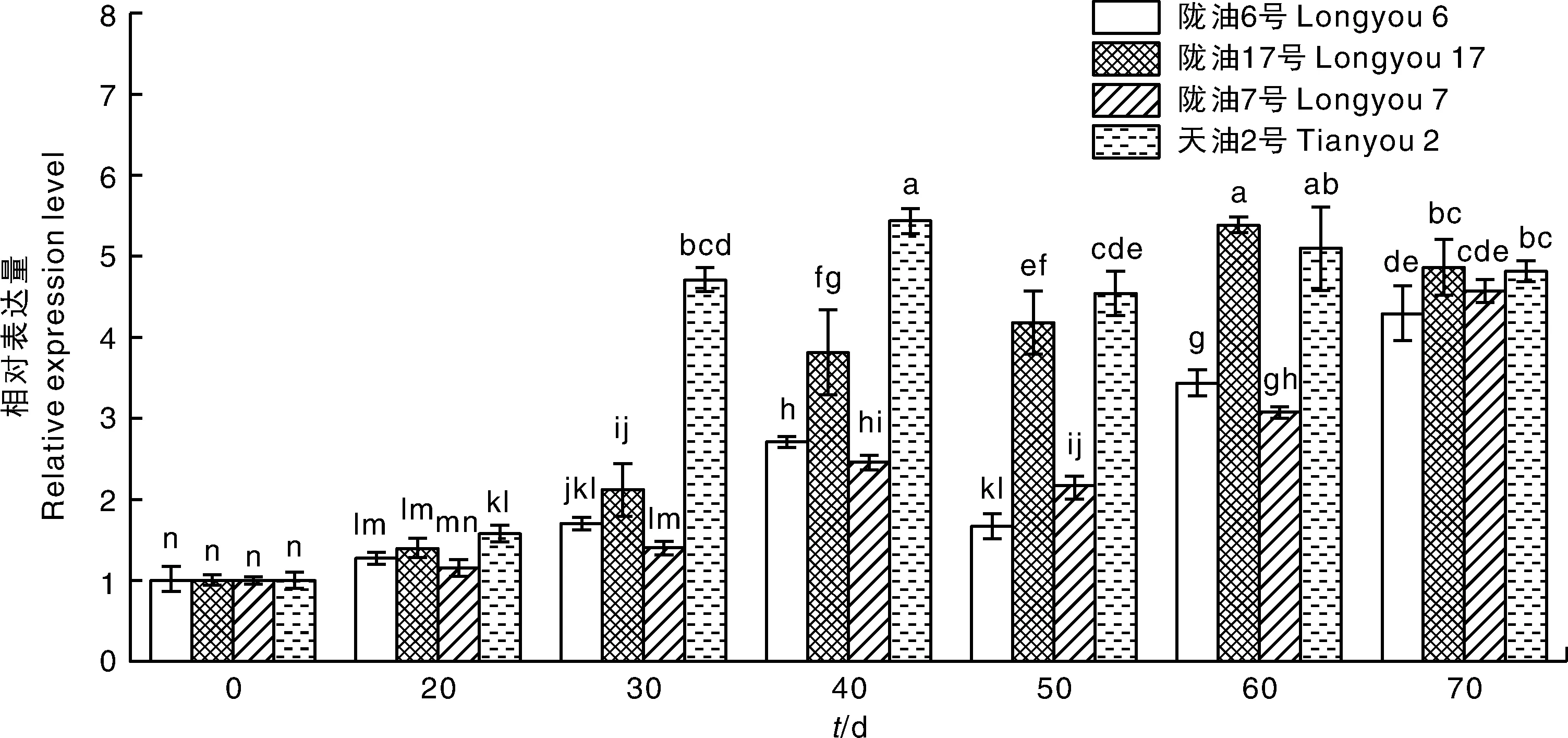
图8 BrFT1基因在不同白菜型冬油菜4 ℃,16 h光照/8 h黑暗处理中的荧光定量表达Fig.8 Quantitative expression of BrFT1 gene in different varieties of winter rapeseed at 4 ℃, 16 h light/8 h darkness
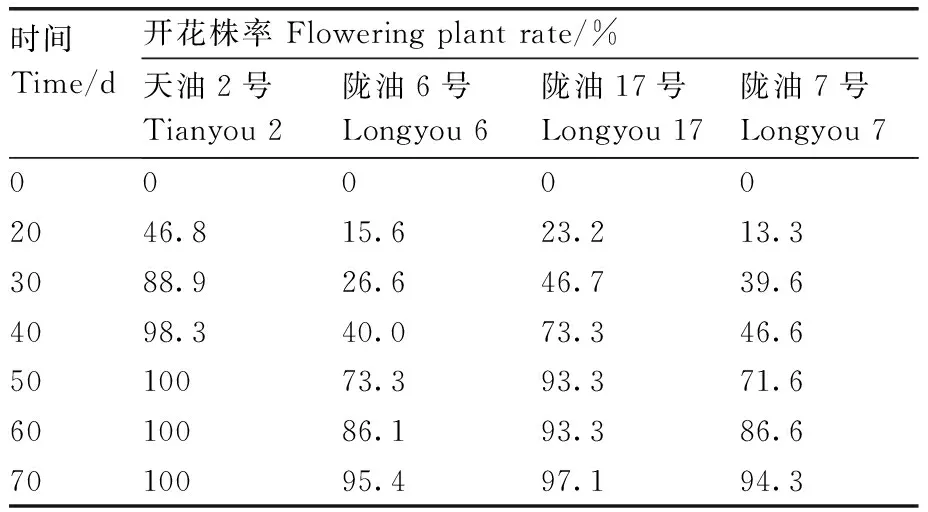
表3 不同处理时间对白菜型油菜开花株率的影响Table 3 Effect of different treatment time on flowering plant rate of Brassica rapa L.
3 讨论
PEBP蛋白在植物种子发育,花序结构形成和成花转变过程中发挥重要作用[21-22],FT属于PEBP蛋白的一个亚类,目前已报到的FT拟南芥中2个[23],甘蓝型油菜中6个FT的拷贝[24],玉米中12个[25],水稻中13个[26],棉花、桃中各1个[27],苹果中2个[28]、草莓中3个[29]。本研究参照拟南芥中FT基因,通过甘肃农业大学最新测序获得的白菜型冬油菜基因组,共鉴定得到3个BrFT基因,其数量与拟南芥接近,对白菜型油菜BrFT蛋白结构详细分析发现,所编码的蛋白均含有PEBP特定的保守结构域,且都含有相同或相似的外显子和内含子结构,共线性分析表明,BrFT1和BrFT2存在明显的共线性关系,由此推测,二者在功能方面具有一定的相通性。启动子元件分析表明,BrFT含有大量的光响应元件,推测它们能够对白菜型油菜外界光照做出应答,这与甘蓝型油菜中A7-FT启动子区调控元件的研究结果相似[30]。另外BrFT1和BrFT2在启动子区存在参与植物生长发育、激素诱导的作用元件,推测它们能够参与植物生长发育、植物激素的调控网络。因此,下一步将对激素和白菜型冬油菜BrFT基因的相互关系进行研究。
本研究发现,BrFT在白菜型油菜的各器官均表达,且叶中的表达量高于其他器官部位,这与已有研究报道FT在叶片和维管束组织中特异性表达相似[31]。克隆得到了白菜型油菜陇油6号、陇油17号、天油2号BrFT1基因,发现不同品种的感温性与BrFT1基因的序列并无关系,可能不同白菜型油菜的开花时间不受BrFT基因本身结构特征的影响。qRT-PCR分析结果表明,在4 ℃,16 h光照/8 h黑暗处理下,使BrFT1的表达被激活,呈现出上调表达,并且在不同感温性白菜型油菜品种中BrFT1基因的过量表达能够相应的引起植株开花,推测长日照通过提高BrFT1基因的表达量来相应激活蛋白活性,从而利于白菜型油菜开花。研究发现,相同处理时间下,BrFT1的表达水平与不同感温性白菜型油菜开花时间有关,推断BrFT1的表达可能还受其他调控因子的影响,结合前面对于启动子区调控元件的分析,预测这些因子有可能是参与基因调控植物生长发育通路中的上游调控因子,这些结果也为白菜型冬油菜的进一步研究提供了线索。
4 结论
本研究首次从白菜型冬油菜基因组中检索到3个BrFT家族基因,生物信息学分析显示,3个家族成员外显子、内含子、保守Motif分布相似,基因启动子区均含光应答元件,另外存在参与植物生长、激素诱导元件。筛选在白菜型油菜中表达量最高的BrFT1基因进行克隆及qRT-PCR,表明白菜型冬油菜的不同感温性与BrFT1基因序列无关,但不同感温性冬油菜开花时间与该基因表达模式有关,可能在4 ℃,16 h光照/8 h黑暗处理过程中该基因的表达受到了其他调控因子的影响。该结论为进一步研究基因功能提供了参考依据。
