基于线粒体DNA D-loop 全序列的麒麟鸡遗传多样性研究
2023-06-07李威娜郭美慧温锦添翁茁先陈洁波杜炳旺黄勋和
李威娜,郭美慧,温锦添,翁茁先,陈洁波,杜炳旺,黄勋和
(1.嘉应学院生命科学学院,广东 梅州 514015;2.广东省山区特色农业资源保护与精准利用重点实验室,广东 梅州 514015;3.广东省五华三黄鸡科技创新中心,广东 梅州 514015)
【研究意义】麒麟鸡,又称粤西卷羽鸡、翻毛鸡,是产于我国大陆最南端粤西一带的特色畜禽品种资源,具有悠久的饲养历史[1-3]。但麒麟鸡因长期没有受到必要的保护而几近灭绝。全面摸查麒麟鸡保种群体的遗传多样性水平和品种,对该品种资源的保护和利用具有重要的科学价值。【前人研究进展】线粒体DNA(mtDNA)D-loop因突变速率快、无重组、含量高等特点,被广泛应用于家鸡遗传资源评估和历史追溯[4]。基于mtDNA D-loop 序列研究表明,五华三黄鸡保种群具有较高的遗传多样性水平,品种的形成可能受北方家鸡南迁的影响[5];广东省黄鸡保持较高的遗传多样性水平,品种的形成受到邻省、北方家鸡和东南亚红原鸡的影响[6]。对华东地区11 个地方鸡品种的遗传多样性和母系起源进行研究,检测出欧美高产品系的遗传渗入信号[7]。黄勋和等[8]基于mtDNA D-loop 序列评估了我国南方28个黄鸡地方品种的遗传多样性水平,结果表明黄鸡地方品种存在杂交现象,群体经历了历史扩张,提示东南亚和我国西南地区的家鸡对我国南方黄鸡地方品种的形成有一定的遗传贡献。麒麟鸡是粤西一带的特色品种,为保护这一珍稀地方品种资源,2011 年起广东海洋大学对其开展了系统研究,全面摸清了麒麟鸡的生物学特性[9-11],阐明了麒麟鸡卷羽性状的发育规律[12]、遗传规律[13],并揭示了卷羽性状独立的分子形成机制[14-15]。
【本研究切入点】目前已形成了以黄羽麒麟鸡为主体、白羽麒麟鸡和黑羽麒麟鸡为特色的资源保护群体。这3 个资源群的主要区别在于羽色,黄羽麒麟鸡的羽毛为黄色,白羽麒麟鸡属于隐性白羽类型,黑羽麒麟鸡的羽毛为黑色,少部分有乌骨性状。本研究团队前期已利用mtDNA D-loop部分序列研究了黄羽麒麟鸡的遗传多样性,并探讨其遗传起源[16]。但目前黄羽麒麟鸡未与周边地方鸡品种进行遗传比较分析,白羽麒麟鸡和黑羽麒麟鸡的遗传多样性及其与周边地方鸡品种的亲缘关系也尚未明确。【拟解决的关键问题】本研究利用mtDNA D-loop 全序列研究麒麟鸡3 个资源群体的遗传多样性,探讨麒麟鸡与周边地方鸡品种的遗传关系,以揭示麒麟鸡的品种形成历史。
1 材料与方法
1.1 试验材料
试验于2022 年在广东省山区特色农业资源保护与精准利用重点实验室开展。90 份麒麟鸡(来自黄羽、白羽、黑羽群体各30 份)血液样品采自广东盈富农业发展有限公司,其他地方鸡血液样品来自当地保种场,各10 份(表1),均为母鸡样品。血液样品的基因组DNA 提取采用HiPure Blood DNA Mini Kit 试剂盒(广州美基生物科技有限公司)。
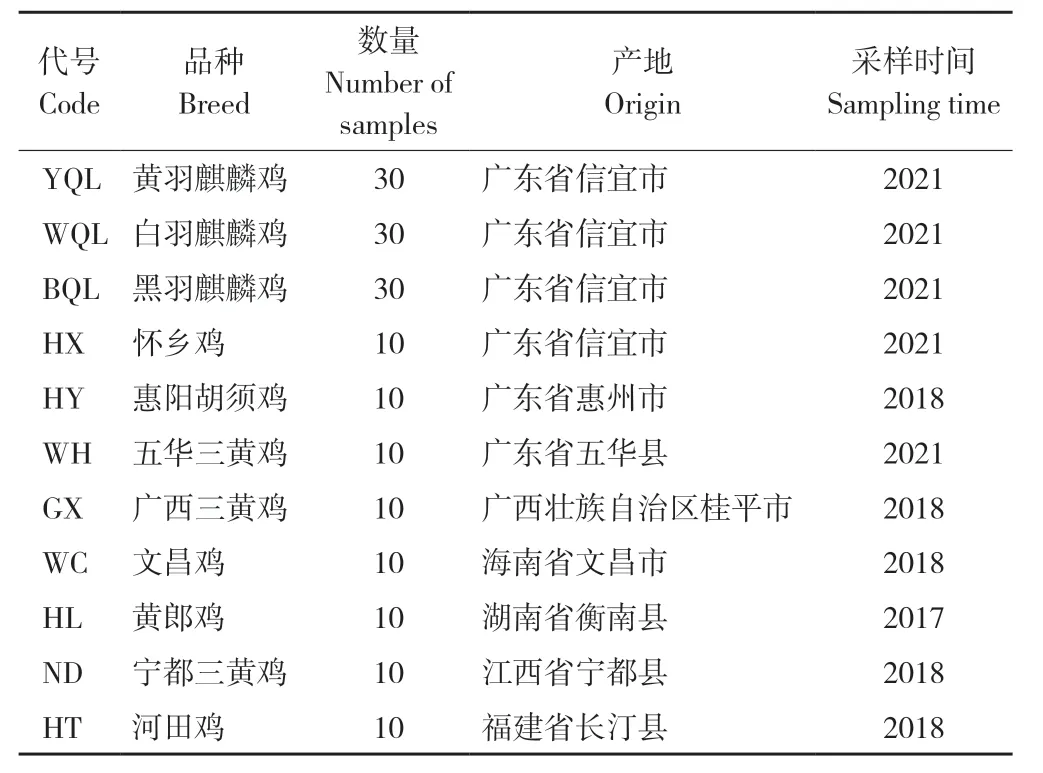
表1 样品信息Table 1 Sample information
1.2 PCR 扩增与序列测定
mtDNA D-loop全序列PCR扩增引物:L16750,5'-AGGACTACGGCTTGAAAAGC-3'[17];H1374,5'-TTTCACTGGGATGCGGATACT-3'[18]。PCR 反应体系(30 μL)包括Premix Taq〔宝生物工程(大连)有限公司〕15 μL,正反向引物(20 μmol/L)各0.3 μL,模板DNA 1 μL,灭菌ddH2O 13.4 μL。PCR 反应条件:94 ℃预变性4 min;94 ℃变性30 s、58 ℃退火50 s、72 ℃延伸80 s,35个循环;72 ℃延伸7 min。目标条带经1.5%琼脂糖凝胶电泳确认后,送生工生物工程(上海)股份有限公司进行双向测序。
1.3 序列分析
测序序列用软件Bioedit[19]进行人工校对,以红原鸡mtDNA D-loop 全序列(GenBank 登录号:NC_007235)作为参考序列,采用MEGA 7.0[20]进行序列比对后,使用DnaSP 6.12[21]定义单倍型,提取变异位点,计算单倍型多样性、核苷酸多样性以及进行中性检验(Tajima'sD和Fu'sFs检验)。通过NCBI BLAST 功能(https://blast.ncbi.nlm.nih.gov/Blast.cgi)搜索与麒麟鸡单倍型完全匹配的中国地方鸡D-loop 全序列。碱基组成分析和群体间的遗传距离(Kimura 2-parameter 模型K2P)计算应用MEGA 7.0[20]完成。应用软件MitoToolPy(http://mitotool.kiz.ac.cn/)获取序列变异信息,然后将序列划入特定的单倍型类群[18,22]。采用软件PopART 1.7[23]绘制麒麟鸡及其他8 个地方鸡品种的mtDNA D-loop 全序列中介网络图。
2 结果与分析
2.1 麒麟鸡遗传多样性分析
麒麟鸡 mtDNA D-loop 全序列为1 231~1 232 bp,供试样本中有72 个在核苷酸位置859 处缺失碱基C。碱基T、C、G、A 的平均含量分别为33.7%、26.5%、13.3%、26.5%,T+A 的平均含量(60.2%)高于C+G 的平均含量(39.8%)。通过90 份麒麟鸡样品共检测到45 个变异位点,占总分析位点的3.65%,其中多态位点43 个、简约信息位点29 个、单一位点突变14 个。以红原鸡mtDNA 序列NC_007235 作为参考,在麒麟鸡45个变异位点中定义了17 个单倍型,其中独享型单倍型9 个、共享型单倍型8 个(表2)。黄羽麒麟鸡的单倍型数量最多(12 个),白羽麒麟鸡次之(7 个),黑羽麒麟鸡最少(5 个)。在其他8个地方鸡品种中,共定义了19 个单倍型,其中河田鸡的单倍型数量最多(8 个),宁都黄鸡最少(3个),没有发现与麒麟鸡共享的单倍型。通过搜索中国家鸡D-loop 全序列发现,与麒麟鸡单倍型2、3、8 和14 序列完全一致的地方鸡品种,主要有金湖乌骨鸡、竹乡鸡、略阳乌鸡、绿耳乌鸡、仙居鸡、崇仁麻鸡、白耳黄鸡、江边鸡、平铺麻黄鸡、藏鸡等(表2)。
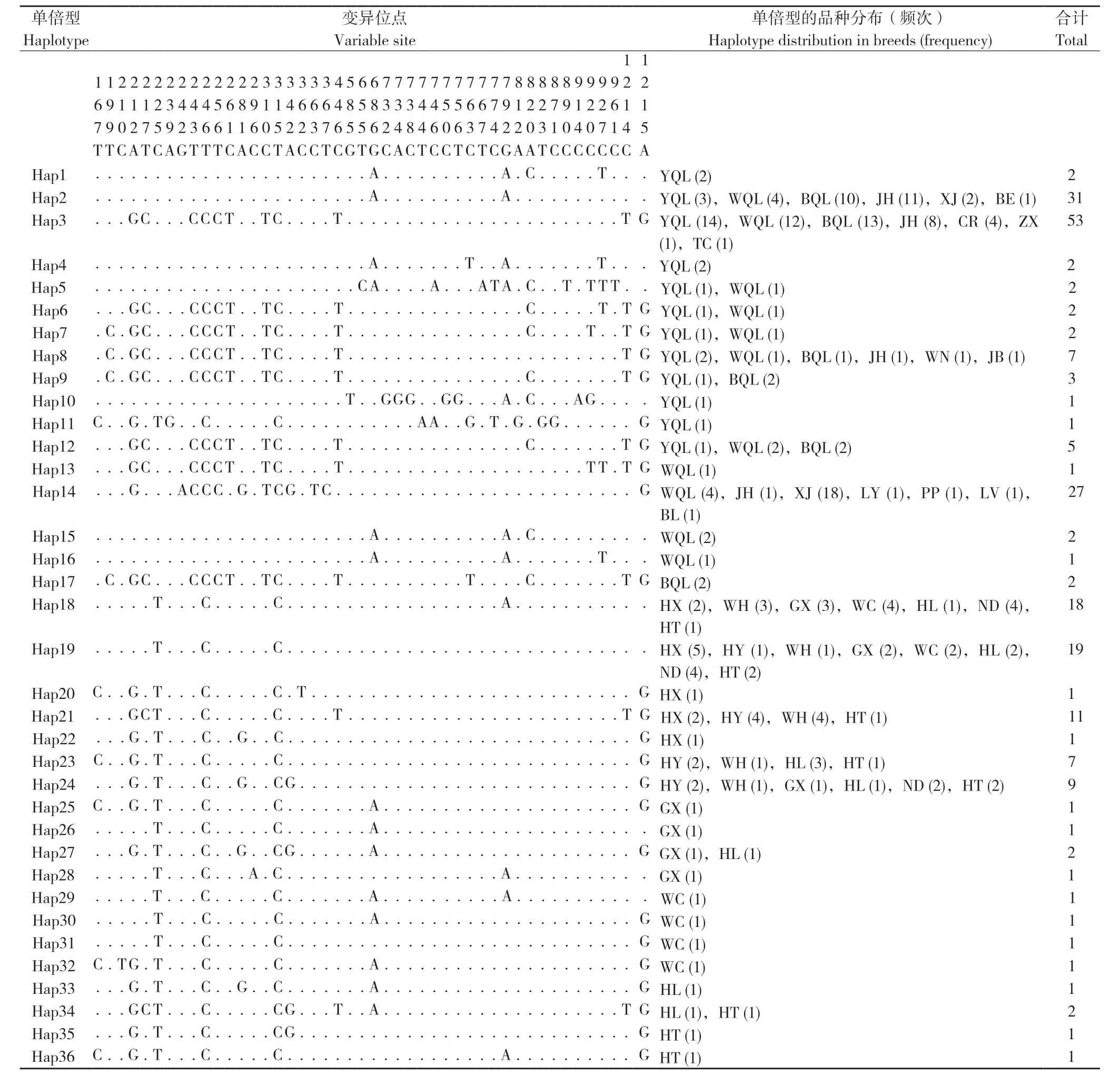
表2 麒麟鸡和8 个地方鸡品种的mtDNA D-loop 单倍型及其在不同群体的共享单倍型数量Table 2 mtDNA D-loop haplotypes and number of shared haplotypes in Qilin chickens and 8 indigenous chicken breeds
麒麟鸡总体单倍型多样性和核苷酸多样性分别为0.773±0.039 和0.00630±0.00054(表3)。3 个麒麟鸡群体中,黄羽麒麟鸡遗传多样性最高,黑羽麒麟鸡最低。在麒麟鸡周边8 个地方鸡品种中,河田鸡的遗传多样性最高,宁都黄鸡最低(表3)。然而,麒麟鸡的核苷酸多样性明显高于其他8 个地方鸡品种。中性检测结果显示,9 个鸡种的检测值差异均不显著,但黑羽麒麟鸡的Fu'sFs和Tajima'sD值均差异显著。
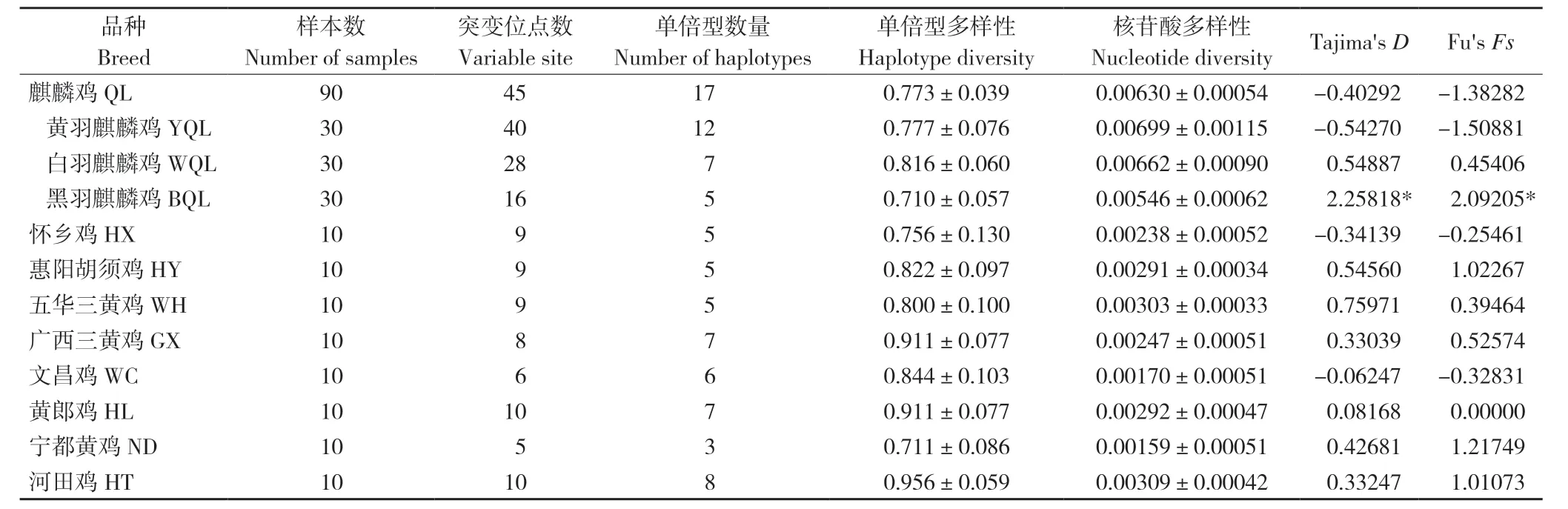
表3 麒麟鸡和8 个地方鸡品种的mtDNA D-loop 遗传多样性Table 3 Genetic diversity of mtDNA D-loop of Qilin chicken and 8 indigenous chicken breeds
9 个鸡品种间的K2P 遗传距离介于0.002~0.009 之间,麒麟鸡与其他8 个地方鸡品种的遗传距离均大于其他8 个地方鸡品种间的遗传距离;品种间的净遗传距离为0~0.005,麒麟鸡与8 个地方鸡品种的净遗传距离均大于其他8 个地方鸡品种间的净遗传距离(表4)。
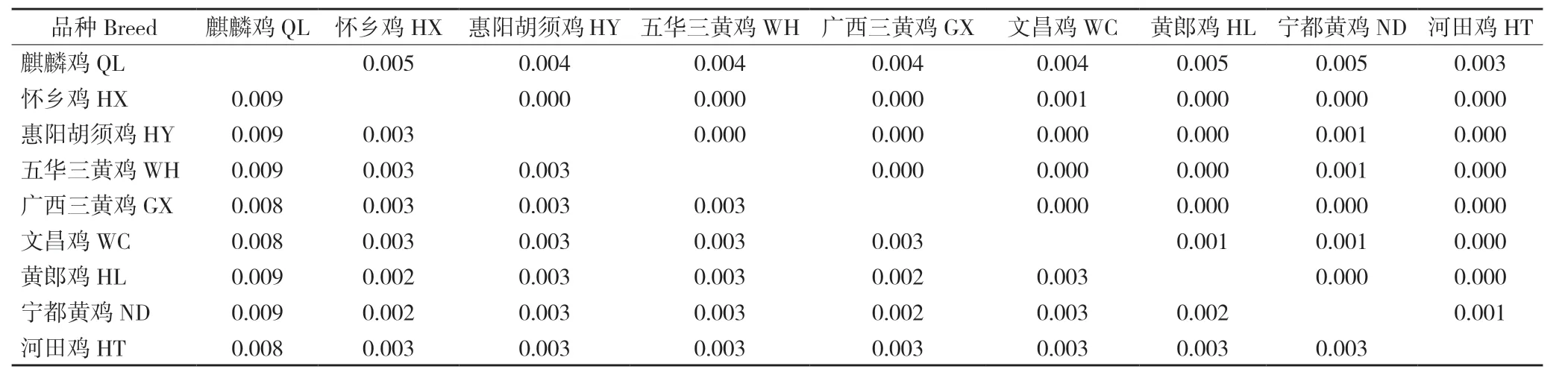
表4 麒麟鸡和8 个地方鸡品种的群体间遗传距离Table 4 Genetic distance among populations of Qilin Chicken and 8 indigenous chicken breeds
2.2 麒麟鸡遗传进化分析
以麒麟鸡和8 个地方鸡品种的mtDNA D-loop全序列构建中介网络图(图1),36 个单倍型可划分为单倍型类群A、B、C 和E,其中A、B、E是优势单倍型类群,C 只有1 个单倍型。麒麟鸡独享17 个单倍型,归属于单倍型类群A、B、C和E,频率分别为1.1%(1/90)、30.0%(27/90)、4.4%(4/90)和64.4%(58/90),B 和E 为优势单倍型类群;黄羽麒麟鸡和白羽麒麟鸡独享单倍型最多,各4 个。其他8 个地方鸡品种共有19 个单倍型,归属于单倍型类群A 和B,频率分别为75.0%(60/80)和25.0%(20/80),其中12 个为单个品种的独享单倍型,文昌鸡占4 个。
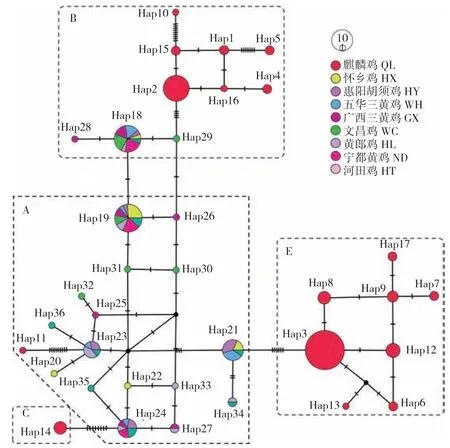
图1 麒麟鸡和8 个地方鸡品种mtDNA D-loop 单倍型的中介网络图Fig.1 Median network of mtDNA D-loop haplotypes of Qilin chicken and 8 indigenous chicken breeds
3 讨论
mtDNA D-loop 全序列分析表明,总体上麒麟鸡保持相对较高的遗传多样性水平,核苷酸多态性明显高于本研究供试的其他8 个地方鸡品种,也高于江西、浙江、山东等地方鸡的核苷酸多态性[7],这可能与其尚未开展系统选育有关。麒麟鸡有着1 000 多年的饲养历史,但长期未受到足够重视,2011 年起由广东海洋大学研究团队开始了较为系统的研究,同步开展了麒麟鸡提纯复壮和初步选育。在选育过程中,出现了部分白羽和黑羽的麒麟鸡后代,在此基础上逐步扩大白羽和黑羽麒麟鸡群体数量,形成了以黄羽麒麟鸡为主体、白羽麒麟鸡和黑羽麒麟鸡为特色的资源群体。麒麟鸡3 个资源群体的遗传多样性与其群体数量呈正相关,黑羽麒麟鸡的群体数量最少,因此单倍型数量和核苷酸多态性均最低。与2016 年有关麒麟鸡mtDNA D-loop 部分核苷酸序列(1~524)遗传多样性[16]的研究结果相比较发现,黄羽麒麟鸡的单倍型多样性、核苷酸多样性分别为0.628±0.063 和0.00859±0.00114,虽然单倍型多样性略有下降,但核苷酸多样性有所提高,说明黄羽麒麟鸡保种群体较为稳定。
地方品种的形成是自然环境和人文历史综合选择的结果[24]。虽然本研究供试的周边地方鸡品种样品较少,但麒麟鸡的17 个单倍型全部为独享型,不与其他8 个地方鸡品种共享,说明麒麟鸡具有相对独立的品种形成历史。麒麟鸡在粤西尤其在茂名一带备受民众喜爱,但在其他地区的销售量则有限。麒麟鸡的卷羽是不完全显性遗传性状[14-15],分为全卷和半卷两种类型,从外观上容易与片羽类型区分。从遗传距离可以看出,麒麟鸡与其他地方鸡的遗传交流较小,这可能是麒麟鸡保持独特性状和遗传特性的重要原因之一。在我国其他地区也有一些羽毛外翻的地方鸡,如贵州倒毛鸡[25]、湖南永顺倒毛鸡[26]、浙江开化翻毛鸡[27]、云南怒江翻毛鸡[28]等,均生长在我国南方地区,卷羽性状与其散热功能相吻合。这些卷毛鸡都是在本地鸡群体中发现少量卷羽个体后,经过扩群和本地选育而来,推测不同卷羽鸡品种的形成是独立事件。通过扩大比对范围发现,归属于单倍型类群B、C 和E 的4 个麒麟鸡单倍型与国内部分地方鸡品种一致[18,29],并且不少为乌骨鸡品种,这可能使部分麒麟鸡有乌骨性状,也说明了麒麟鸡是由本土品种选育而来。后续还需扩大样品收集范围,利用基因组重测序等分析技术以进一步研究麒麟鸡的品种形成历程。
根据线粒体基因组的划分方法,可将全球家鸡和原鸡划分为A~I 和V~Z[18,30-31]。特定的单倍型类群有特定的区域分布,如A、B 和C 在东亚家鸡是优势单倍型类群,D 主要在东南亚和太平洋地区分布,E 则全球广泛分布,G 主要在我国西南家鸡中分布[8,18,31]。mtDNA D-loop 全序列中介网络图将麒麟鸡划分为单倍型类群A、B、C和E,可认为麒麟鸡具有4 个母系来源。我国地方鸡品种主要以B 为优势单倍型类群[8],而麒麟鸡为E。值得注意的是,E 在中快速型肉鸡以及高产蛋鸡中是绝对优势单倍型类群[32-33]。鉴于麒麟鸡体型明显大于其他地方鸡品种,麒麟鸡在品种形成过程中是否有外来血源渗入仍有待进一步研究。
4 结论
本研究利用mtDNA D-loop 控制区全序列对麒麟鸡进行遗传多样性分析,黄羽麒麟鸡、白羽麒麟鸡和黑羽麒麟鸡的单倍型多样性分别为0.777、0.816、0.710,核苷酸多样性分别为0.00699、0.00662、0.00546,其中黄羽麒麟鸡多样性最高,且核苷酸多态性明显高于周边8 个地方鸡品种,表明保种效果良好。麒麟鸡有4 个单倍型类群来源,且B 和E 是优势单倍型类群,而8 个地方鸡品种单倍型类群主要为A 和B,表明麒麟鸡保持相对较高的遗传多样性水平。麒麟鸡17 个单倍型不与周边地方鸡品种共享,但4 个单倍型与国内乌骨鸡等品种共享,这一发现佐证了该品种的遗传独特性,为麒麟鸡的品种形成研究提供了新视角。
