不同林龄光皮桦林土壤养分含量与酶活性相关关系
2023-06-04余春和韦秋思刘世男
余春和,曾 珠,韦秋思,刘世男,杨 梅
(1.广西壮族自治区国有雅长林场,广西百色 533209;2.广西大学林学院,南宁 530004)
土壤养分和土壤酶是森林生态系统的重要组成成分[1],在维持植物生长过程中作用显著。土壤养分是存在于土壤中的矿质营养成分,是植物生长必不可少的成分。土壤养分含量大小直接影响林木生长速度和质量[2]。土壤中氮、磷、钾等元素是林木生长发育所需的大量营养元素,驱动着其他元素的循环与转化,是养分循环的核心[3]。土壤酶是起催化作用的蛋白质总称,在土壤能量转化和物质循环过程中起着不可忽视的作用,可加快有机质的降解速度。土壤酶活性是土壤对植物代谢需求和有效养分的直接表达[4],其高低可以反映土壤养分(尤其是氮、磷)周转的快慢[5],被认为是评价和衡量土壤肥力的重要指标[6]。陈彩虹等[7]认为土壤酶活性与土壤养分因子间存在极显著或显著的相关关系。土壤脲酶及蛋白酶活性与土壤中磷、钾的循环呈负相关,碱解氮和全磷分别是蛋白酶和磷酸酶活性的主要影响因子[8,9]。然而,在植物生长和代谢过程中,土壤中酶活性、有机质和氮、磷、钾的高低均与树种、树龄有一定的相互性。相关研究表明,人工林随着种植年限的增加,其土壤酶活性会产生显著变化[10]。土壤脲酶、酸性磷酸酶、蔗糖酶均与人工林土壤质量密切相关,其活性能迅速对土壤养分变化产生响应[11]。目前对森林土壤中酶活性与土壤养分的研究,多集中在混交林分、间伐强度、林下植被等方面,而对于不同林龄人工林的土壤养分含量与酶活性之间的关系鲜有报道。
光皮桦(Betula luminifera)属于桦木科(Betulaceae),是优良速生阔叶树种[12]。在火烧迹地、采伐迹地或森林破坏后的荒山上常与松、栎等喜光树种组成次生混交林,是一种值得推广的荒山造林先锋树种。目前,已有一些对光皮桦种子园建设[13]、基因功能[14]、育苗技术[15]、营林技术[16]、群落特征[17]、养分循环[18]、木材特性[19]等方面的研究报道,这些已有研究对光皮桦的栽植培育、开发利用和经营管理起到良好的推动作用。本研究对土壤养分以及酶活性的研究,有利于进一步了解不同酶活性和养分循环在不同林龄光皮桦林中的变化特征,为光皮桦的经营管理及其土壤改良作用的发挥提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地位于广西壮族自治区西北部的雅长林场(106°08′—106°23′E,24°37′—25°00′N),地跨百色市的乐业县和田林县,属亚热带气候。研究区土壤主要为黄红壤或黄壤,该地区雨量充沛,冬短夏长,干湿季节明显,年平均气温为16.8 ℃,年无霜期为357 d;年降雨量为1 056 mm,年蒸发量最高达1 484.7 mm,相对湿度为82%,每年6—10 月为雨季,11 月至次年5 月为旱季[20],该地区自然条件较适合光皮桦的生长。
1.2 样地设置和土壤采样
试验地造林前为无林地,没有乔木及灌木,只有草本植物。造林时间分别为2007、2013、2017 年的5 月,初始造林密度为1 666 株/hm2,造林株行距为2 m×3 m,造林地未施基肥和追肥,造林后前3年进行坎草抚育。林下植被主要有盐肤木(Rhus chinensis)、糙叶榕(Ficus irisana)、野牡丹(Melastoma candidum)、路边青(Geum aleppicum)等灌木,铁芒箕(Dicranopteris dichotoma)、臂形草(Brachiaria eruciformis)、龙须草(Eulaliopsis binate)、五节芒(Miscanthus floridulus)等草本植物。
林分调查时间为2020 年10 月,选择地形要素及土壤类型等立地条件相近的林地,在3 个林龄的光皮桦林中设置20 m×20 m 标准样地各3 块,以无林地的标准样地为对照。在每个标准样地内,按照“S”取样法,选取5 个样点,在样点处分层(0~20 cm 和20~40 cm)采集土壤样品,各样点相同土层混合为1 个样本。将采集的土壤保存于无菌保鲜袋带回实验室。将土壤样品分为2 部分:一部分放在室内自然风干,去除杂质,研磨过2 mm 筛,用于土壤养分含量测定。另一部分过2 mm 筛,置于4 ℃冰箱冷藏保存,用于过氧化氢酶、蔗糖酶、脲酶、酸性磷酸酶活性的测定。光皮桦林分基本概况见表1。
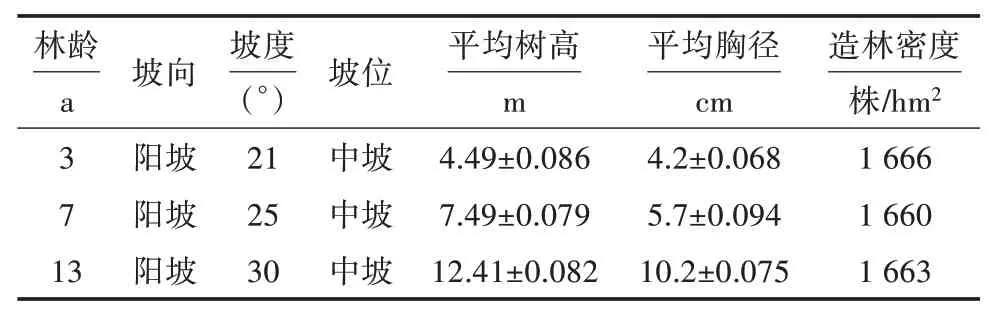
表1 不同林龄光皮桦林分基本信息
1.3 土壤养分和酶活性的测定
混合土样中氨态氮含量采用纳氏试剂分光光度法测定,速效钾含量采用乙酸铵浸提-火焰光度法测定,速效磷含量采用碳酸氢钠-钼锑抗比色法测定,全氮含量采用半微量凯氏蒸馏法测定,全钾含量采用氢氧化钠碱熔-火焰光度法测定,全磷含量采用浓硫酸-高氯酸消煮-钼锑抗比色法测定[21]。
过氧化氢酶活性的测定采用高锰酸钾滴定法,蔗糖酶活性的测定采用硫代硫酸钠滴定法,脲酶活性的测定采用比色法,土壤酸性磷酸酶活性的测定采用磷酸苯二钠比色法[22]。
1.4 数据处理
运用软件Excel 对数据进行整理、汇总,运用软件SPSS 20.0 对数据进行统计分析。显著性检验采用单因素方差分析和最小显著差异法,相关性分析采用Pearson 法。
2 结果与分析
2.1 光皮桦土壤养分特征
从表2 可知,在不同林龄光皮桦林中,土壤各养分含量表现出不同的变化趋势。0~20 cm 土层中氨氮含量与20~40 cm 土层中氨氮和全氮含量的变化趋势大致相同,含量依次为13 年生>无林地>3 年生>7 年生,且在0~20 cm 土层中,氨氮含量在不同林龄光皮桦林间呈显著差异(P<0.05)。0~20 cm 土层和20~40 cm 土层中,全钾、速效钾和速效磷含量在7 年生光皮桦林均最低。0~20 cm 土层和20~40 cm 土层中,土壤中全磷含量依次为3 年生>7 年生>13 年生,表现为随林龄增大呈逐渐减少的趋势;7 年生光皮桦林土壤中氨氮、全氮、全钾、速效钾、速效磷和全磷含量较3 年生光皮桦林的含量均有所下降,但氨氮、速效钾、速效磷、全氮和全钾含量在林龄为13 年时均达到最大。在垂直方向上,随着土壤深度的增加土壤中各养分含量均呈下降趋势。
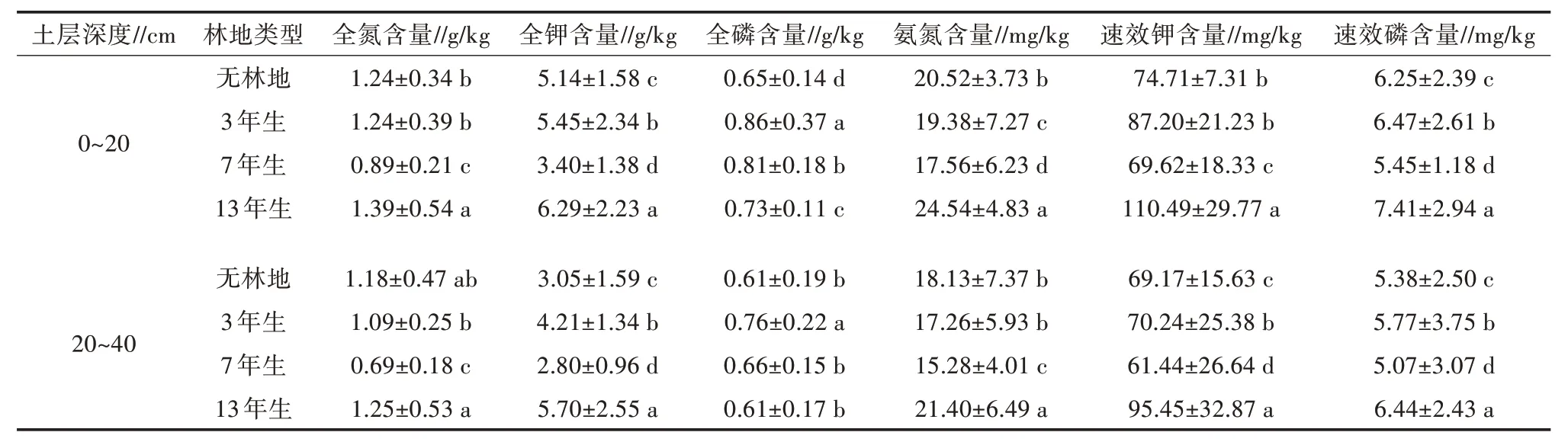
表2 光皮桦林地土壤养分含量
2.2 光皮桦土壤酶活性特征
不同种类的土壤酶活性随林龄的变化,表现出不同的变化规律,如图1 所示,在土壤的垂直方向上,过氧化氢酶、脲酶、蔗糖酶和酸性磷酸酶活性均随土层深度增加而降低,且同一林龄中不同土层酶活性之间差异显著(P<0.05),其变化趋势与土壤养分在土壤中的分布基本一致。这4 种土壤酶活性在3、7、13 年生林分中均显著高于无林地,仅过氧化氢酶活性在7 年生林分显著低于无林地。
在0~20 cm 土层和20~40 cm 土层中,蔗糖酶活性随着林龄的增大呈现持续上升的变化趋势,表现为13 年生>7 年生>3 年生>无林地,不同土层13 年生光皮桦林土壤中的蔗糖酶活性较3 年生分别增加了1.56 倍和2.15 倍;而过氧化氢酶、脲酶和酸性磷酸酶活性表现为13 年生>3 年生>7 年生>无林地,7 年生光皮桦林土壤中酶活性较3 年生有所下降;13 年生光皮桦林土壤中4 种土壤酶活性显著大于同层无林地对照;13 年生光皮桦林各土层土壤酶活性显著高于7 年生林地。综上,不同林龄光皮桦林土壤酶活性存在显著差异,随着林龄的增加,4 种酶的酶活性总体上呈波动上升的趋势。
2.3 土壤养分和酶活性的相关关系
分别对不同林龄光皮桦林0~20 cm、20~40 cm土壤养分指标和土壤酶活性进行相关性分析,如表3 所示。在0~20 cm、20~40 cm 土层中,过氧化氢酶和脲酶活性与土壤中全钾、氨氮、速效钾、速效磷含量均呈极显著正相关,与全氮含量虽为正相关关系,但仅在0~20 cm 土层中呈显著正相关;土壤全磷含量与4 种酶活性的相关性较小。在20~40 cm 土层中,蔗糖酶活性与全磷含量呈负相关,但相关性较小,为-0.300;蔗糖酶活性与全钾、速效钾含量呈极显著正相关。酸性磷酸酶活性与各土层中全氮、全钾、全磷、氨氮、速效钾、速效磷含量均为正相关关系。在0~20 cm、20~40 cm 土层中,速效钾含量均随酸性磷酸酶活性的增强而增加;速效磷含量与酸性磷酸酶活性在2 个土层中略有差异,在20~40 cm 土层呈极显著正相关,在0~20 cm 土层呈显著正相关。这4 种土壤酶活性均与速效磷和速效钾含量有较强的相关性。
3 小结与讨论
土壤氮、磷、钾元素作为植物必需的养分,它们的含量和分布影响植物的生长发育。土壤氮素供应量影响林木的生长状态及速度[23]。土壤磷元素影响植物生长、形态变化、养分利用效率、吸收利用机制等[24]。钾元素通过增强作物的抗病虫害、抗倒伏、抗旱和抗寒等抗逆能力,可促进作物生长发育[25]。本研究通过对不同林龄光皮桦林中土壤养分和土壤酶活性测定与研究发现,0~20 cm 土层中土壤养分含量和土壤酶活性高于20~40 cm 土层,这与表层土壤中全氮、全磷含量较高,随着土层的加深其含量减少的结果相似[26],有学者研究结果表明,土壤酶活性也呈现相似的规律[27,28]。土壤酶的主要来源是微生物、动植物残体及植物根系分泌物。由于表层土壤的通气性较强,微生物分解凋落物活动较强,养分和分泌的酶积累在土壤表层,随着土层加深,微生物活动减弱,因此土壤养分和酶活性呈下降趋势[29,30]。
在时间尺度上,土壤养分和酶活性的变化规律存在一定差异。3、7、13 年生光皮桦林中养分含量和酶活性普遍比无林地高。可能与人为活动影响有关,如在施肥等管理措施的干扰下,林下凋落物分解、根系分泌也会增加土壤养分和酶活性。本研究中,进入速生期后,7 年生光皮桦林土壤氨氮、全氮、全钾、全磷、速效磷和全磷含量均小于3 年生,可能是由于7 年生光皮桦林进入到速生期,林分郁闭度增大、林内光照减弱、林下植被开始减少、土壤微生物活性降低,从而使地表枯落物分解变慢、土壤接收养分含量减少,需要从土壤中吸收大量养分,酶活性随之降低。种植光皮桦林13 年后,光皮桦生长过了高峰期,进入稳定生长时期,此时自然稀疏量强烈,凋落物大量增加,土壤养分的吸收和回归趋于平衡,土壤条件得到改善,这时期土壤养分含量逐渐上升,酶活性增强,因此13 年生林分土壤养分含量较7 年生林分显著增加[31]。
土壤酶活性对土壤养分核心要素氮、磷、钾元素的转化和迁移具有重要作用。相关性分析表明,土壤氨氮、速效钾和速效磷含量和土壤酶活性相关性较大,说明有效养分含量对土壤酶活性的影响更强烈[32]。过氧化氢酶、脲酶、蔗糖酶和酸性磷酸酶活性与氮、磷、钾含量呈正相关,其中,过氧化氢酶和脲酶活性与全钾、氨氮、速效钾、速效磷含量呈极显著正相关,与罗亚进[33]的研究结果一致。当土壤中有可利用养分时,微生物生长较快[34],需分泌较多的酶以满足自身需求。土壤酶能够活化土壤中各元素的化合物,进而提升土壤有效养分、改善土壤质量。而严绍裕[10]研究发现,速效磷和速效钾含量与过氧化氢酶活性呈负相关。可能是因为本研究对0~20 cm 和20~40 cm 土层的酶活性和养分含量分开进行相关分析,土壤酶活性与土壤养分同样具有明显的空间异质性,不同土层、不同季节土壤酶活性差异明显[35]。为使光皮桦林可持续发展,光皮桦林生长到速生期时需加强水肥管理,采取合理的抚育措施,为光皮桦林土壤适当补充氮肥、磷肥和钾肥,促进光皮桦林生长,并为养分循环提供基本保障。
