茉莉酸甲酯对橡胶树花药愈伤乳管细胞分化的影响
2023-05-30郭子晗谭德冠黄东益付莉莉孙雪飘张家明
郭子晗 谭德冠 黄东益 付莉莉 孙雪飘 张家明


关键词:橡胶树;愈伤组织;乳管细胞分化;茉莉酸甲酯;基因表达
中图分类号:S794.1 文献标识码:A
橡胶树(Hevea brasiliensis Muell. Arg.)属于大戟科橡胶属,是重要的热带经济作物,在我国主要分布于海南、云南、广东等热带北缘地区[1]。乳管是橡胶树胶乳生物合成和贮存的唯一场所,是由无数个乳管细胞所组成的网状结构器官[2-3]。橡胶树树皮被定期开割,树皮中被割开的乳管中会排出奶汁状的胶乳(细胞质),胶乳经收集和加工后,成为重要的工业原料——天然橡胶(顺-1,4-聚异戊二烯)。世界上99%的商品天然橡胶来源于橡胶树[4]。
由于树皮中乳管的数量是决定天然橡胶产量的最重要因素[5-6],为此许多学者聚焦于橡胶树乳管分化研究。大部分学者用大田植株枝条或树干的树皮作为研究对象,通过外界物理或化学刺激来研究树皮乳管分化情况[7-9]。近期有学者以橡胶树愈伤组织为研究对象,通过在培养基中添加化学物质来研究愈伤乳管分化情况[10]。
茉莉酸(JA)具有促进橡胶树乳管分化功能的这一发现是橡胶树乳管分化研究中取得的突破性进展。最初HAO 等[7]發现外源JA 能促进橡胶树枝条树皮乳管分化;TAN 等[10]也发现培养基中添加一定浓度的外源JA 能促进橡胶树愈伤组织中乳管细胞分化。当前,科研人员围绕JA 调控橡胶树乳管分化的信号分子开展一系列研究并取得一定进展。HbSKP1、HbCOI1、HbJA1、HbLAP1、HblMYC1、HblMYC2、HbEREBP1 等与茉莉酸信号途径相关的基因被克隆出来[11-14];发现橡胶树乳管细胞存在特定的茉莉酸信号转导模块COI1-JAZ3-MYC2,其通过上调法尼基焦磷酸合酶(FPS)基因和小橡胶粒子蛋白(SRPP)基因的表达来增强橡胶生物合成[15]。
茉莉酸甲酯(MeJA)与JA 同属茉莉酸类物质(JAs),但MeJA 对橡胶树愈伤乳管细胞分化的效果尚不清楚。本研究将应用橡胶树愈伤组织来系统研究MeJA 对乳管细胞分化的效果,并研究其所调控橡胶合成相关基因表达情况,对促进橡胶树愈伤组织在乳管分化研究中的应用,以及开发新型橡胶树增产刺激剂从而促进橡胶树天然橡胶的生产具有一定意义。
1 材料与方法
1.1 材料
于2021 年3 月在中国热带农业科学院试验农场三队采集橡胶树品种‘热研7-33-97的春花,将采集好的花枝立刻置于4℃冰箱中保存24~48 h,备用。
1.2 方法
1.2.1 橡胶树花药愈伤组织的诱导 选取已低温处理过的处于单核期的橡胶树雄花,包裹于纱布中,于75%(V/V)酒精中浸泡30 s,然后置于0.15%(W/V)升汞中浸泡8 min,无菌水清洗5遍。于无菌条件下从雄花中剥离出雄蕊,接种于相应的培养基中。
诱导花药愈伤组织培养基配方参考文献[16],具体为改良MS+2,4-D 1 mg/L+ KT 1 mg/L+NAA1 mg/L+8%(W/V)蔗糖+2.5 g/L Phytagel。为了研究MeJA 对花药愈伤组织乳管细胞分化的效果,在诱导花药愈伤组织培养基中分别添加0、1、2、3 mg/L 的MeJA。上述的植物激素和Phytagel均购自Sigma 公司。培养基pH 为6.2,培养条件为25℃黑暗条件。
1.2.2 橡胶树花药愈伤组织鲜重测量 取培养至第65 天的不同MeJA 浓度处理的愈伤组织块于电子天平中称重,每个处理重复20 次。
1.2.3 组织化学法鉴定愈伤组织乳管细胞 组织化学法参照田维敏等[17]的方法进行。取培养至第65 天的不同MeJA 浓度处理的愈伤组织于70%(V/V)酒精中固定48 h,酒精系列脱水,冰醋酸过渡处理4 h,于60℃下溴-碘染色液处理48 h。冰醋酸浸洗3 次,每次2 h;无水酒精脱水3 次,每次1 h;正丁醇透明3 次,每次2 h。材料被渗蜡、包埋后,经莱卡滑走切片机切成10 μm 厚的薄片。切片经二甲苯脱蜡、固绿染色、无水酒精脱水、二甲苯透明处理后,再用中性树胶封片。光学显微镜下观察制备好的切片。
1.2.4 愈伤组织乳管细胞分化频率 光学显微镜下观察不同MeJA 浓度处理的花药愈伤组织的组织化学切片,每处理随机观察30 个视野,统计每个视野中乳管细胞的分化频率。愈伤组织乳管细胞分化频率=(乳管细胞数量/总细胞数量)×100%。
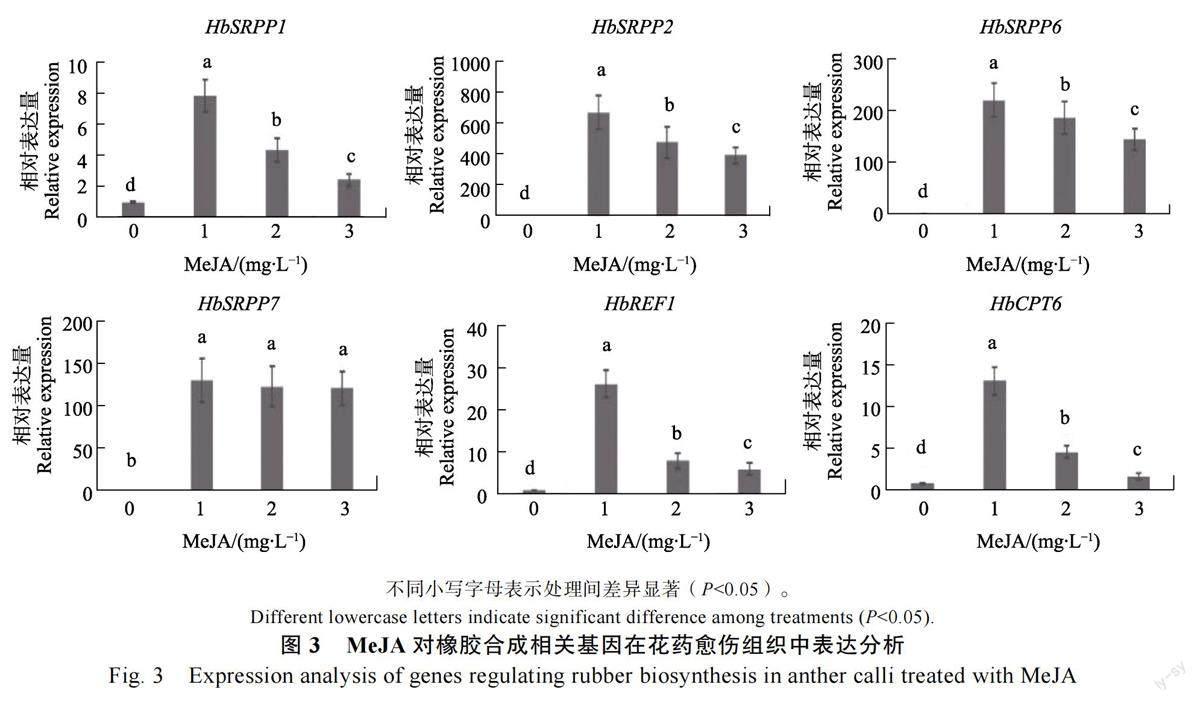
1.2.5 荧光定量PCR 分别取培养至第65 天的不同MeJA 浓度处理的愈伤组织,采用北京天根生化科技有限公司的RNAprep Pure 植物总RNA提取试剂盒(产品号:DP432)提取愈伤组织RNA,再用Thermo Scientific 公司的RevertAid FirstStrand cDNA Synthesis Kit 试剂盒(产品号:#K1622)将愈伤组织总RNA 反转录为第一链cDNA。根据TANG 等[18]报道的调控橡胶树橡胶生物合成的重要基因序列信息,选取与橡胶合成密切相关的小橡胶粒子蛋白(small rubber particleprotein,SRPP)基因的4 个家族成员(HbSRPP1、HbSRPP2、HbSRPP6、HbSRPP7)、橡胶延伸因子(rubber elongation factor, REF)基因的家族成员HbREF1、橡胶转移酶(cis-Prenyl transferase,CPT)基因的家族成员HbCPT6 的序列设计引物(表1),用于荧光定量PCR 反应。以在橡胶树胶乳中稳定表达的eIf2 基因[19]作为参考基因。利用宝生物公司的SYBR Premix Ex TaqTM II 试剂盒(产品号:RR820A)进行荧光定量PCR 反应,反应条件为95℃预变性30 s,95℃变性15 s,58℃退火15 s,72℃延伸15 s,反应共进行40 个循环。每处理含有3 个生物学重复。采用2?ΔΔCT 法计算基因相对表达量。
1.2.6 电镜制片 电镜超薄制片参考照田维敏等[17]的方法进行。取培养至第65 天的1 mg/L MeJA 处理的愈伤组织置于预冷的4%戊二醛溶液中4℃下固定24 h,0.2 mol/L PBS 缓冲液清洗2 次,2%锇酸4℃下再固定10 h,乙醇系列脱水,环氧丙烷渗透,环氧树脂包埋,切成50~70 nm 薄片,醋酸铀和柠檬酸铅染色,透射电子显微镜(HT-7700)下观察。
2 结果与分析
2.1 MeJA 对橡胶树花药愈伤组织生长的影响
在诱导橡胶树花药愈伤组织培养基中添加不同浓度MeJA,会明显影响到愈伤组织块的生长。培养至65 d 时,各处理的愈伤组织均呈现鲜黄色,表面凹凸不平(图1A~图1D)。MeJA 浓度为1 mg/L 的处理其愈伤块平均鲜重(FW)为0.23 g,虽然略低于对照(0.24 g),但统计学上二者无显著差异,说明MeJA 在0~1 mg/L 范围内对愈伤组织的生长影响不明显。MeJA 浓度为2、3 mg/L的处理愈伤块平均FW 分别为0.18 g 和0.13 g,均显著低于对照和1 mg/L MeJA 的处理(图1E),说明MeJA 在2~3 mg/L 范围内会抑制愈伤组织生长。对MeJA 浓度与愈伤FW 进行相关性分析(图1F),相关系数R 为-0.971,表明二者负相关,说明随着培养基中MeJA 浓度的增加,抑制花药愈伤组织生长的程度会加大,愈伤组织FW 呈现下降趋势。上述结果表明低浓度的MeJA(0~1 mg/L)对花药愈伤组织的影响不明显,而高浓度的MeJA(2~3 mg/L)会抑制愈伤组织生长,且MeJA 浓度与愈伤组织生长呈负相关关系。
2.2 MeJA 对橡胶树花药愈伤组织乳管细胞分化的影响
溴-碘染色法是一种经典可靠的鉴别乳管细胞方法[20]。溴-碘染色的石蜡制片显示培养至65 d的花药愈伤组织中存在被染成棕色的细胞,是由于该细胞中的橡胶内含物被溴-碘溶液染成棕色,这些细胞为乳管细胞(图2A~图2D)。切片中还存在一些黑褐色单宁细胞,该类细胞的单宁内含物被染成黑褐色。此外,愈伤组织中还存在一些淀粉细胞,这些细胞内充满颗粒状的淀粉粒。
培养基中添加一定浓度的MeJA 会促进橡胶树花药愈伤乳管细胞分化。当培养基中MeJA 浓度为1 mg/L 时,愈伤乳管细胞发生频率上升至15.8%,显著高于其他处理,比对照(5.7%)增加近2 倍(图2A、图2B、图2E)。当培养基中MeJA 浓度为2 mg/L 时,愈伤乳管细胞发生频率为13.0%,虽然显著高于对照,但却显著低于1 mg/L MeJA 的处理(图2C、图2E)。当培养基中MeJA 浓度为3 mg/L 时,愈伤乳管细胞发生频率下降至9.7%,虽然也显著高于对照,但显著低于1、2 mg/L MeJA 的處理(图2D、图2E)。上述结果说明培养基中添加低浓度的MeJA 能促进愈伤组织乳管细胞分化,而高浓度的MeJA 会抑制其分化,其中MeJA 浓度为1 mg/L 效果最佳。
2.3 胶乳合成相关基因在MeJA 处理的花药愈伤组织中表达分析
SRPP、REF、CPT 是存在于小橡胶粒子膜的蛋白质,是参与橡胶生物合成的重要酶和调控因子,它们所编码的基因在乳管细胞中特异表达[21-24]。笔者检测橡胶树SRPP、REF、CPT 基因在MeJA 处理的花药愈伤组织中的表达情况,发现它们表达量的变化趋势与愈伤乳管细胞发生频率的变化趋势基本一致。荧光定量PCR 结果显示,与对照相比,培养基中添加1~3 mg/L MeJA 后均能显著促进HbSRPP1、HbSRPP2、HbSRPP6、HbSRPP7、HbREF1、HbCPT6 的表达(图3)。这是由于添加MeJA 后愈伤组织中乳管细胞数量增加所致。当培养基中添加1mg/L MeJA 时愈伤乳管细胞发生频率最高,此时HbSRPP1、HbSRPP2、HbSRPP6、HbSRPP7、HbREF1、HbCPT6 表达量最高,分别为对照的7.8、676.6、222.3、130.7、26.1、13.0 倍。当培养基中添加2、3 mg/LMeJA 时愈伤乳管细胞发生频率呈下降趋势,此时除了HbSRPP7 外,其他基因表达量均随着MeJA 浓度的升高而下降。这些结果进一步证实MeJA 能促进橡胶树愈伤组织中乳管细胞分化。
2.4 橡胶树花药愈伤乳管细胞的超微结构
虽然通过溴-碘染色法能确切证实愈伤组织中存在乳管细胞,但愈伤乳管细胞的超微结构尚不清楚。橡胶树树皮乳管细胞的大尺寸橡胶粒子直径为0.40~0.75 μm,中等尺寸橡胶粒子直径为0.25~0.35 μm, 小尺寸橡胶粒子直径为0.08~0.20 μm[21]。本研究采用透射电镜技术来研究花药愈伤组织中乳管细胞的超微结构,发现愈伤乳管细胞中也存在大、中、小尺寸橡胶粒子,其中大尺寸橡胶粒子直径为0.20~0.30 μm,中等尺寸粒子直径为0.10~0.18 μm,小尺寸粒子直径为0.04~0.08 μm(图4)。与树皮乳管细胞的橡胶粒子相比较,这些愈伤乳管细胞的橡胶粒子尺寸均较小,其原因可能是它们都处于发育早期,也可能是由于乳管细胞来源不同所致。愈伤乳管细胞内还含有黄色体,它们呈球形囊泡状,内部含高电子致密物质(图4B)。
3 讨论
已有研究表明JAs 具有能够调节植物抗逆和抗病虫害的防御反应,同时还具有调节植物生长发育的功能[25-26]。橡胶树乳管细胞中的胶乳主要起防御功能[27-28],除了含有橡胶烃外,还含有一些有毒物质,如羟基腈水解酶,用来催化腈基类化合物水解,释放氢氰酸,抵御害虫和病原菌的侵害[29]。在本研究中,培养基中添加MeJA 具有促进橡胶树花药愈伤组织中乳管细胞分化的功能;而随着愈伤组织中乳管细胞数量增加,所含的胶乳也会增多,因此愈伤组织中含有毒物质的浓度可能会增高,这些有毒物质可能会抑制愈伤组织生长,这可能是MeJA 影响愈伤组织生长的主要原因。
在本研究中,培养基中MeJA 浓度为1 mg/L时为橡胶树花药愈伤组织乳管细胞分化的最佳浓度。而TAN 等[10]发现培养基中JA 浓度为2 mg/L时为橡胶树花药愈伤组织乳管细胞分化的最佳浓度。但在本研究中高浓度MeJA 处理其愈伤乳管细胞分化的变化趋势与TAN 等[10]报道的相一致。这些结果表明MeJA 与JA 诱导愈伤乳管细胞分化的最佳浓度虽存在一定差异,但二者调控愈伤组织乳管细胞分化的效果相似,说明MeJA 可代替JA 进行乳管细胞分化研究。
MeJA 与JA 同属茉莉酸类物质(JAs),但二者市场价格差异巨大,前者价格低廉,根据Sigma公司网站的报价,后者价格约为前者的140 倍(https://www.sigmaaldrich.cn/CN/zh)。由于JA 价格过于昂贵,限制其在橡胶树大田生产试验与应用。JA 应用于橡胶树乳管分化的研究主要集中于枝条树皮,发现JA 能促进枝条树皮分化次生乳管[7-8]。于俊红等[9]开展了JA 对成龄橡胶树次生乳管分化的实验,发现JA 对成龄橡胶树次生乳管分化具有促进效果,但它们仅涂1 g 含1% JA的羊毛脂于面积2 cm×2 cm 的用刀片轻刮至绿色的橡胶树皮层,1 个月后取处理部位树皮进行组织化学鉴定。目前尚未能像橡胶树增产刺激剂乙烯利那样涂抹于成龄橡胶树割口处来开展JA 能否促进橡胶树增产的大田实验。本研究发现,MeJA 能促进愈伤组织乳管细胞分化,表明其可代替JA 进行乳管细胞分化研究。因此,在橡胶树的大田生产试验中,将来可用价格低廉的MeJA来代替价格昂贵的JA 进行,这对开发新型橡胶树增产刺激剂,促进橡胶树生产具有一定意义。
