微管调节试剂对白菜型冬油菜萌动种子内源激素及抗寒基因表达的影响
2023-05-27田海燕冯秀英徐明霞董小云郑国强魏家萍武泽峰崔俊美谭雪莲刘自刚
田海燕,冯秀英,徐明霞,董小云,郑国强,李 辉,王 莹,魏家萍,武泽峰,崔俊美,谭雪莲,刘自刚
(1.甘肃农业大学农学院/甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室,甘肃 兰州 730070;2.武山县农业技术推广中心,甘肃 天水 741300;3.甘肃省农业科学院旱地农业研究所,甘肃 兰州 730070)
油菜是我国重要的油料作物,我国50%以上的食用油来源于油菜,同时油菜还是动物饲料、蔬菜、绿肥和生物柴油等的重要来源[1]。长期以来,低温胁迫是限制我国油菜产区北移和高产的主要非生物因素,严重影响我国油菜产业的发展。当植物受到胁迫时,细胞膜上的感受器会首先感知到外界低温,植物接收到冷的信号后迅速调节植物体内的抗寒物质,启动抗寒基因的表达,进而激活相应的代谢调控途径,对植物产生保护屏障[2]。环境低温信号与转换是植物冷应答的关键环节,细胞膜是植物感受低温最敏感的部位,由中间磷脂双分子层构成基本骨架,物质通过细胞膜进出,冷应激均会引起细胞骨架重排[3]。细胞骨架是一类蛋白纤维网架体系,由微管、微丝及中间纤维组成,细胞骨架调节细胞膜的物质运输,同时细胞膜骨架整体结构还影响细胞膜上不同通道的离子活性[4]。细胞骨架与跨质膜的细胞外基质互相联结,低温作用于跨膜的胞外受体,然后将冷信号传递给细胞骨架,由细胞外信号传递给生命活动的控制中心,进而对下游相关基因进行调控,形成了一个统一协调的调控网络。
细胞骨架重组对冷诱导的胞质Ca2+浓度的波动 (钙信号) 非常重要,低温胁迫后,质膜生理状态、Ca2+内渗、相关质膜蛋白等在冷信号感受和传递的上游途径中发挥重要作用[5]。在冷胁迫初期,Ca2 +通道和质膜蛋白CRLK1 / 2被激活,随后Ca2 +信号和MAPK级联途径被激活,形成由胞外钙库向胞内的跨膜Ca2+流。Ca2+流作为第二信使,通过MEKK-MPK激酶途径和14-3-3s穿梭途径,分别多泛素化ICE1和CBFs转录因子,从而调控冷信号通路核心位置ICE1-CBF-COR的途径[6]。ICE1被MAPK级联中的MPK3/6磷酸化,导致ICE1被降解,同时,CRPK1磷酸化的14-3-3蛋白从胞浆穿梭到细胞核,促进CBFs的降解,从而调节冷防御反应的持续时间,参与植物耐冷性通路[7]。CBF转录因子在低温应答调控中起关键作用[8]。拟南芥中有三个CBF基因,即CBF1、CBF2和CBF3,其中CBF1和CBF3直接感应冷信号,调控通路上的抗寒基因,低温诱导OST1激酶的活性,ICE1和激活状态的OST1互作使ICE1蛋白磷酸化,提高ICE1蛋白的转录活性,从而提高植物的耐冷性[9]。在冬油菜中抗冻蛋白(AFPs)具有抑制冰晶结合和生长能力,受ICE1-CBF冷信号通路的调控[4]。
秋水仙碱(Colchicine,Col)作为一种微管聚合抑制剂[10],可以改变细胞的发育进程,阻断染色体,使染色体结构发生变异,当Col与正在进行有丝分裂的细胞相遇时,秋水仙碱会与微管蛋白的特定位点结合,进而导致细胞的α微管蛋白结构和β微管蛋白二聚体变形,微管蛋白组装成微管的过程则会被明显阻断,使细胞停止于有丝分裂中期,从而导致细胞死亡[11]。紫杉醇(Taxol,Tax)是一种微管促进和稳定已聚合的微管剂,当细胞与Tax接触后,会在细胞体内积累大量的微管,微管的积累会阻断细胞分裂[12],进而促进或诱导微管蛋白聚合、防止解聚,从而能稳定细胞微管的稳定性[13]。因此,本研究利用微管调节试剂,即微管聚合抑制剂Col和微管聚合促进剂Tax处理白菜型冬油菜萌动种子,分析微管试剂对不同白菜型冬油菜萌动种子相关内源激素和抗寒基因的变化,为油菜出苗的抗寒防冻提供理论基础。
1 材料与方法
1.1 供试材料
以白菜型冬油菜‘陇油7号’和‘天油4号’为试验材料,‘陇油7号’种子由甘肃省干旱生境作物学国家重点实验室油菜课题组提供,‘天油4号’种子由天水市农业科学研究所提供。
1.2 试验设计
挑选籽粒饱满、大小均一的种子,先用75%的酒精消毒5 min,再用ddH2O冲洗种子3次,灭菌滤纸吸干附着水,发芽盒(12 cm×12 cm)中萌发种子,每盒播种100粒。播后将发芽盒置于SPX-GB型植物光照培养箱(光照强度600 μmol·s-1·m-2),每天光照16 h,昼/夜温度(20±1)/(20±1)℃,相对湿度40%,每天浇2 mL 50%浓度的霍格兰营养液。种子萌发5 d后,添加400 μmol·L-1的Col处理6 h,随后用ddH2O水冲洗发芽盒3~5次,去除残留Col溶液。然后将油菜萌动种子放置于20℃和0℃温度下分别处理12 h和24 h后采样。对照添加等体积ddH2O,不加试剂处理。Tax处理时,添加试剂为50 μmol·L-1的Tax处理,其他条件同Col处理。
1.3 内源激素含量测定
采用液相色谱质谱联用仪(Agilent LC-MS,安捷伦公司,美国)测定试样中赤霉素(gibberellin,GA3)、生长素(auxin,IAA)、脱落酸(abscisic acid,ABA)和玉米素(zeatin,ZT)含量。
1.4 白菜型冬油菜中与细胞骨架相关抗寒基因的表达验证
使用TIANGEN公司的植物总RNA提取试剂盒提取白菜型冬油菜萌动种子总RNA,用超微量紫外分光光度计检测总RNA的浓度和质量,取OD260/280值介于1.9~2.1的RNA样品,按TIANGEN FastQuant RT反转录试剂盒反应体系比例要求将RNA反转为cDNA,测定浓度后稀释至50 ng于-20℃保存备用。从GeneBank中挑选出3个拟南芥中与细胞骨架相关的抗寒基因,即MKK2、α-tubulin、β-tubuli,根据这些抗寒基因在拟南芥序列,在NCBI数据库(National Center for Biotechnology Information,https://www.ncbi.nlm.nih.gov/)中进行BLAST-X(https://cipptato.org/genebankcip/)序列同源比对,找到在白菜型油菜中高度同源的相关基因。除上述3个抗寒基因外,同时在白菜型油菜中找到冷调控通路上的关键基因(ICE1、CBF1)和抑制冰核形成抗冻蛋白编码基因(BrAFP1)的同源序列。采用Primer 3 Input(http:/bioinfo.ut.ee/primer3-0.4.0/)在线软件设计real-time PCR引物(表1)。以Actin基因作为内参基因,引物由上海Sangon Biotech有限公司合成。扩增程序为:95℃预变性30 s;95℃ 5 s,60℃ 30 s,95℃ 15 s,40个循环。待扩增完成后,根据扩增曲线和熔解曲线检测其特异性,并利用2-ΔΔCt法对基因表达情况进行分析。
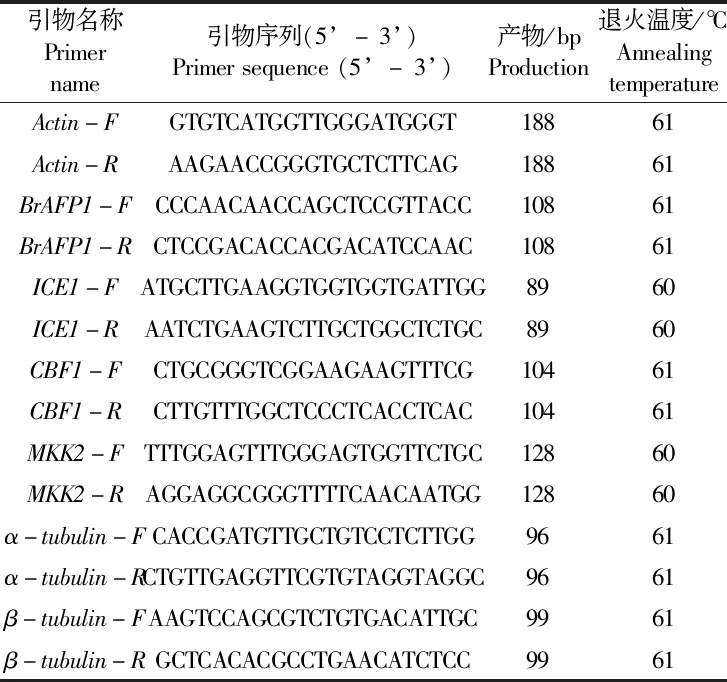
表1 实时PCR定量分析相关基因的引物序列Table 1 The primer sequence of corresponding genes that were used to real-time PCR analysis
1.5 数据处理与分析
所有试验数据均采用Microsoft Excel 2016和SPSS 19.0进行处理和统计分析,数据采用“平均值±标准差”表示,不同试验处理下相应生理性状分别采用GLM(generalized linear model)模型进行联合方差分析。
2 结果与分析
2.1 微管调节试剂对白菜型冬油菜萌动种子ABA含量的影响
由图1可知,与20℃处理相比,0℃处理使‘陇油7号’和‘天油4号’萌动种子ABA明显上升,而Col处理具有降低萌动种子ABA含量的作用,在0℃下更加明显。在20℃处理下,与CK相比,Col处理对不同品种白菜型冬油菜萌动种子ABA含量的影响存在差异,在20℃处理12 h和24 h下,Col处理使‘陇油7号’萌动种子ABA含量分别降低了5.26%和1.29%,而在12 h时Col处理使‘天油4号’ABA含量升高了4.19%,在24 h时Col处理使ABA含量下降了23.48%。在0℃处理下,Col处理能使‘陇油7号’和‘天油4号’萌动种子ABA含量显著下降,0℃处理12 h和24 h下,‘陇油7号’和‘天油4号’萌动种子ABA含量分别降低了55.68%、61.16%和11.22%、52.20%(图1A)。
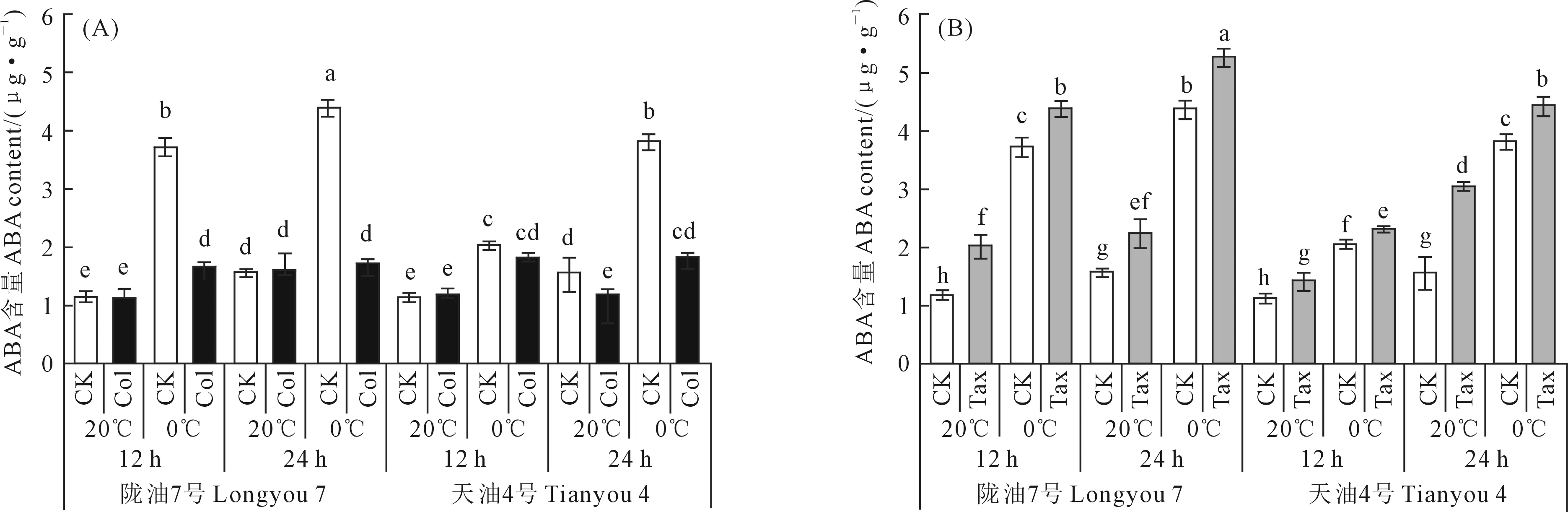
注:不同小写字母表示在0.05水平下差异显著。下同。Note:Different lowercase letters indicate significant difference at 0.05 level.The same below.图1 不同白菜型油菜品种在不同时长、温度和Col/Tax浓度处理下ABA含量Fig.1 The ABA content at different times,temperatures,and Col/Tax concentration treatments in different Brassica rape
Tax处理能使白菜型冬油菜萌动种子ABA含量明显上升;20℃处理12 h和24 h下,与CK相比,Tax处理使‘陇油7号’和‘天油4号’ABA含量分别增加了76.02%、42.49%和26.35%、97.39%。在0℃处理12 h和24 h下,与CK相比,Tax处理使ABA含量分别增加了17.84%、20.34%和13.53%、16.52%(图1B)。综上所述,低温胁迫能使白菜型冬油菜萌动种子ABA含量明显上升,Tax能够促进ABA的上升,Col能够减缓萌动种子ABA。
2.2 微管调节试剂对白菜型冬油菜萌动种子IAA含量的影响
由图2可知,与20℃处理相比,0℃处理使‘陇油7号’和‘天油4号’萌动种子IAA总体上呈现上升趋势,与‘天油4号’相比,‘陇油7号’的变化趋势更加明显(图2A);与CK相比,Col处理使萌动种子IAA含量明显降低。在20℃处理12 h和24 h下,与CK相比,Col处理能使‘陇油7号’和‘天油4号’萌动种子IAA含量显著下降,分别降低了31.17%、83.68%和47.02%、24.69%;在0℃处理12 h和24 h下,与CK相比,Col处理使‘陇油7号’和‘天油4号’萌动种子IAA含量分别降低了65.56%、78.76%和61.28%、35.70%。
在Tax处理下,除‘陇油7号’在20℃处理12 h时IAA含量降低了37.01%,其他处理下两个油菜品种萌动种子的IAA含量都表现出不同程度的升高(图2B)。在0℃处理12 h和24 h下,与CK相比,Tax处理使‘陇油7号’和‘天油4号’IAA含量分别上升了55.11%、33.30%和64.13%、421.75%。在20℃处理12 h下,与CK相比,Tax处理使‘天油4号’IAA含量上升了65.61%;在20℃处理24 h下,Tax处理使‘陇油7号’和‘天油4号’IAA含量分别上升了75.52%和241.98%。综上所述,低温胁迫能使白菜型冬油菜萌动种子IAA含量明显上升,Tax能够促进萌动种子体内IAA含量增加,Col能够减缓萌动种子IAA的产生。
2.3 微管试剂处理对白菜型冬油菜萌动种子GA3含量影响
由图3可知,与20℃处理相比,0℃处理后,‘陇油7号’和‘天油4号’萌动种子GA3含量均呈现下降趋势,且‘天油4号’的下降趋势明显强于‘陇油7号’;与CK相比,Col处理能使‘陇油7号’和‘天油4号’萌动种子GA3含量明显下降。在20℃处理12 h和24 h下,与CK相比,Col处理使‘陇油7号’和‘天油4号’GA3含量分别降低了34.54%、43.74%和14.61%、30.59%;在0℃处理12 h和24 h下,Col处理使‘陇油7号’和‘天油4号’GA3含量分别降低了47.01%、53.49%和31.95%、26.82%(图3A)。
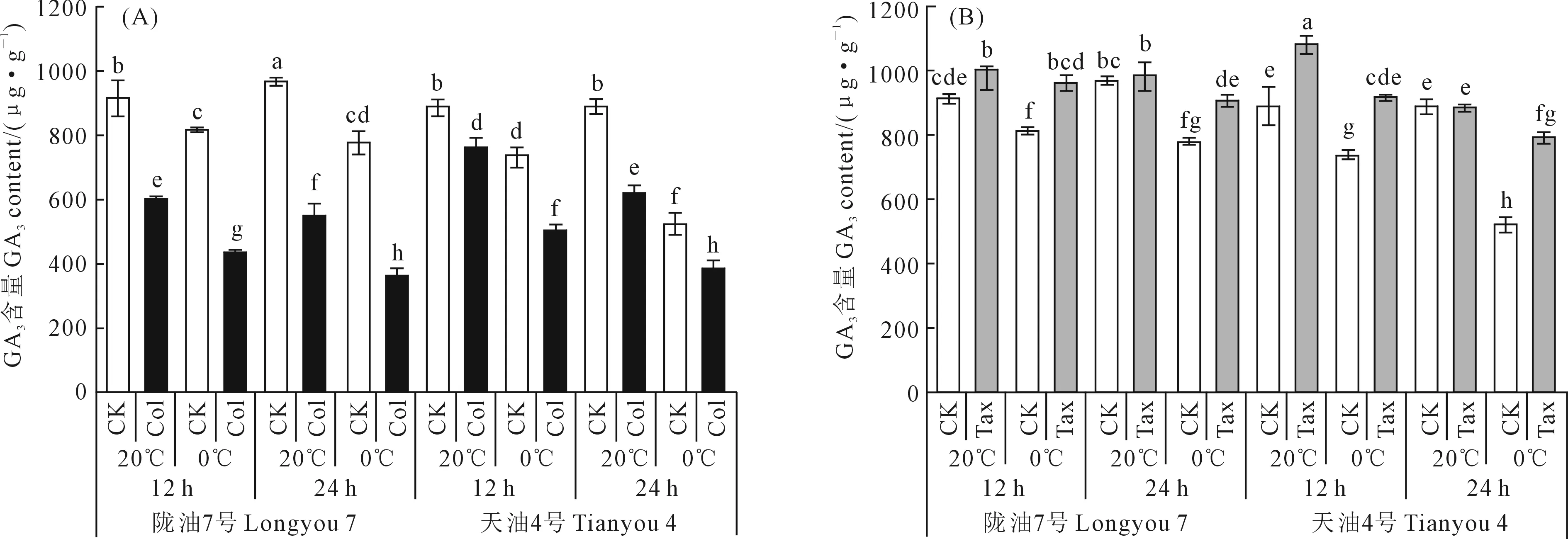
图3 不同白菜型油菜品种在不同时长、温度和Col/Tax浓度处理下GA3含量Fig.3 The GA3 content at different times,temperatures,and Col/Tax concentration treatments in different Brassica rape
与CK相比,Tax处理能使‘陇油7号’和‘天油4号’萌动种子GA3含量明显上升(图3B)。在0℃处理12 h和24 h下,Tax处理使‘陇油7号’和‘天油4号’GA3含量分别上升了18.26%、16.84%和24.97%、51.74%。在20℃处理12 h下,Tax处理使‘陇油7号’和‘天油4号’的GA3含量明显上升,分别增加了9.71%和21.66%。而在20℃的处理24 h下,‘陇油7号’和‘天油4号’萌动种子的GA3含量在处理前后基本无变化。综上所述,在低温胁迫下使白菜型冬油菜萌动种子GA3含量明显下降,Tax处理对GA3产生具有显著的促进作用,而Col能够减缓萌动种子GA3的产生。
2.4 微管试剂处理对白菜型冬油菜萌动种子ZT含量影响
由图4可知,与20℃处理相比,0℃处理使‘陇油7号’和‘天油4号’萌动种子ZT呈现显著上升趋势,而Col处理能够减缓ZT的产生(图4A)。在20℃处理12 h和24 h下,与CK相比,Col处理使‘陇油7号’和‘天油4号’萌动种子ZT含量分别降低了8.57%、70.72%和5.01%、8.57%。在0℃处理12 h和24 h下,Col处理使‘陇油7号’和‘天油4号’ZT含量分别降低了39.35%、86.80%和49.20%、85.77%。
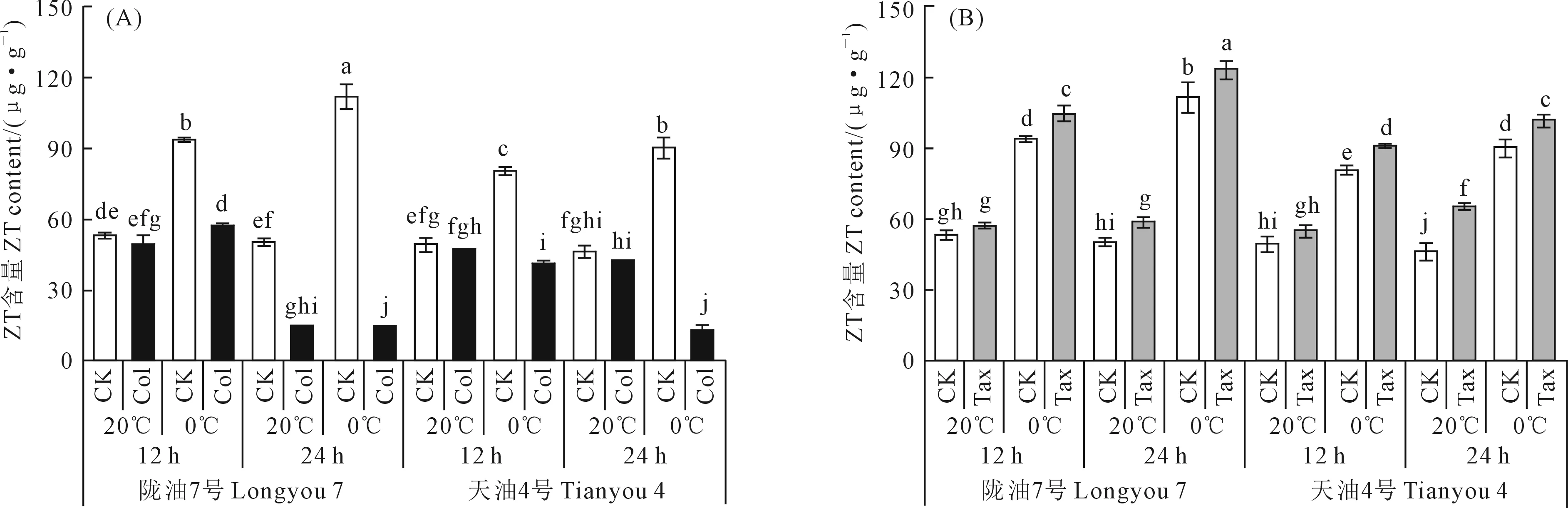
图4 不同白菜型油菜品种在不同时长、温度和Col/Tax浓度处理下ZT含量Fig.4 The ZT content at different times,temperatures,and Col/Tax concentration treatments in different Brassica rape
与CK相比,Tax处理能使白菜型冬油菜萌动种子ZT含量明显上升(图4B)。在20℃处理12 h和24 h下,与CK相比,Tax处理使‘陇油7号’和‘天油4号’ZT含量分别增加了6.89%、17.28%和11.15%、41.56%。在0℃处理12 h和24 h时,与CK相比,Tax处理使‘陇油7号’和‘天油4号’ZT含量分别增加了11.24%、10.30%和12.79%、12.80%。表明低温胁迫能够促进白菜型冬油菜萌动种子ZT的产生,Tax能够促进ZT含量的上升,Col能够减缓萌动种子ZT的产生。
2.5 微管调节试剂处理对白菜型冬油菜抗寒基因表达的影响
由图5可知,在0℃与20℃处理下,Col试剂处理使‘陇油7号’和‘天油4号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1基因的表达量显著高于Tax试剂处理。不同品种间比较发现,添加Col试剂后,与0℃处理12 h相比,0℃处理24 h时,‘陇油7号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量分别上升了102.7%、248.1%、195.4%、72.98%、11.00%、274.4%;‘天油4号’α-tubulin、β-tubulin、CBF1的表达量分别上升了39.31%、17.21%、39.69%,MKK2、ICE1、BrAFP1的表达量分别下降了17.80%、52.68%、92.48%。添加Tax试剂后,与0℃处理12 h相比,0℃处理24 h时,‘陇油7号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量分别下降了100%、99.50%、99.90%、99.37%、32.47%、92.05%;‘天油4号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量分别上升了79.99%、208.8%、169.5%、94.05%、44.79%、78.62%。添加Col试剂后,与20℃处理12 h相比,20℃处理24 h时,‘陇油7号’和‘天油4号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量分别上升了601.9%、329.3%、303.5%、228.6%、24.84%、440.6%和79.77%、3.28%、1.18%、12.72%、10.10%、168.0%;添加Tax试剂后,与20℃处理12 h相比,20℃处理24 h时,‘陇油7号’和‘天油4号’MKK2、α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量分别下降了54.46%、32.85%、31.50%、47.76%、22.61%、11.77%和54.39%、54.45%、46.96%、75.59%、31.91%、76.13%。
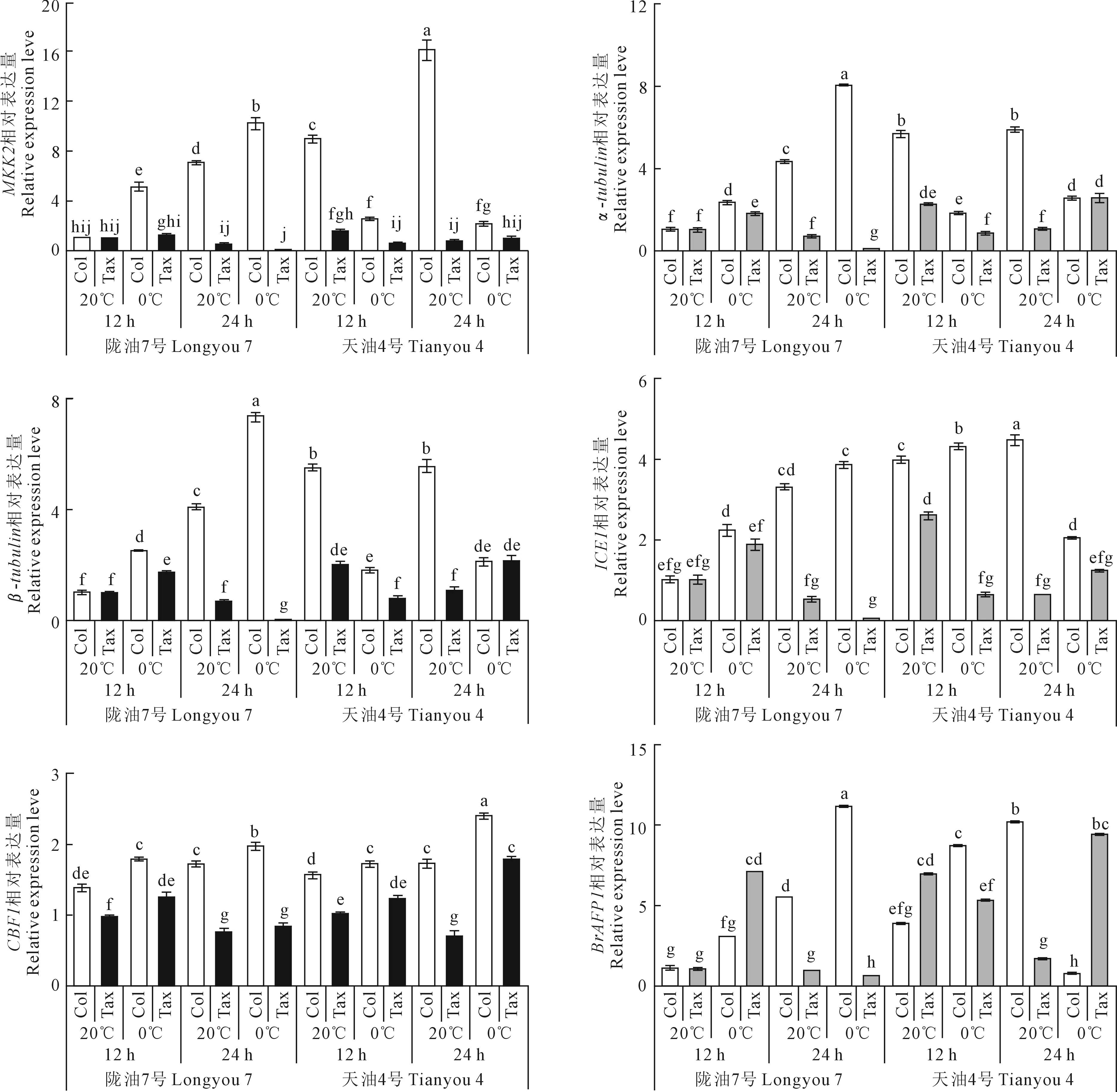
图5 微管调节试剂处理对白菜型冬油菜萌动种子抗寒基因表达的影响Fig.5 Effect of microtubule reagent treatment on cold resistance gene expression in seedling of winter rapeseed
3 讨 论
3.1 微管调节试剂Col和Tax对白菜型冬油菜内源激素的影响
植物内源激素对植物的生长发育具有重要的调控作用,当植物在受到低温胁迫时,会受到多种激素的共同调节,增强植物对胁迫的抵抗能力[14]。研究表明,ABA在冷胁迫中可以促进植物体内渗透调节物质含量的增加[15],从而稳定细胞膜结构,通过ICE1-CBF冷信号通路诱导下游抗寒基因表达等作用,从而提高植物对低温的抵御能力[16]。前人在小麦[17]、茶树[18]及玉米[19]等作物中研究表明,ABA可以提高植物的抗寒性。GA3能降低自由水含量,维持细胞膜的稳定性,GA3含量升高可以提高植物的抗寒性[20]。IAA的代谢也会影响植物的抗寒性[21],在一定的低温处理下,IAA含量升高可以提高油菜的抗寒能力[22]。ZT含量增高会加速植物新陈代谢和蛋白质的合成,使有机体迅速增长,从而提高植物抗寒能力,减轻低温对植物的伤害[23]。本研究发现,内源激素在不同油菜品种、不同微管调节试剂、不同温度和处理下均存在一定差异。施加Tax试剂后,在0℃与20℃处理下会引起‘陇油7号’和‘天油4号’ABA、IAA、GA3和ZT含量显著上升,施加Col试剂则使‘陇油7号’和‘天油4号’ABA、IAA、GA3和ZT含量下降。说明施加Col和Tax试剂会影响白菜型油菜萌动种子内源激素含量。
3.2 微管调节试剂Col和Tax对白菜型冬油菜抗寒基因表达量的影响
在低温胁迫下MAPK级联起着重要的作用,典型的MAPK级联包含三种蛋白激酶(MAPKKK、MAP3K或MEKK)、MAPK激酶(MAP2K、MKK或MEK)和MAP激酶(MAPK),其中MKK2与MPK4(MAPK4)和MPK6(MAPK6)相互作用来调节MKK2的活性,同时MKK2还诱导CoR基因的表达[24]。ICE1与CBF基因的启动子区结合激活表达,CBF结合到其下游目的基因启动子的DRE序列上,诱导COR的表达,从而提高植株的抗寒性[25]。α-微管蛋白和β-微管蛋白是植物微管蛋白的主要组成部分,也是细胞骨架的重要组成部分,植物微管蛋白基因会受温度调控[26]。低温下α-微管蛋白表达上调会维持细胞骨架的稳定性[27]。微管在抗寒中的作用与植物的部位以及温度有很大的关系[28]。AFPs具有修饰冰晶形态,可以抑制冰晶的生长和重结晶等作用,在抵御外界低温冻害中具有重要作用,可以提高植物的抗寒性[29]。本研究发现,抗寒基因在不同油菜品种、不同微管调节试剂、不同温度和处理下均存在差异。添加Col后,0℃处理24 h时,‘陇油7号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量明显升高,‘天油4号’α-tubulin、β-tubulin、CBF1的表达量上升,但MKK2、ICE1、BrAFP1的表达量呈现下降趋势;20℃处理24 h时,‘陇油7号’和‘天油4号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量均呈上调趋势。添加Tax后,0℃处理24 h时,‘陇油7号’α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量被下调,‘天油4号’α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量呈现上升趋势;20℃处理24 h时,‘陇油7号’和‘天油4号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量均呈下降趋势。说明施加Col和Tax试剂会影响白菜型油菜萌动种子抗寒基因的表达。
4 结 论
本研究发现,Col能够降低‘陇油7号’和‘天油4号’萌动种子的ABA、IAA、GA3和ZT含量,而Tax能够增加‘陇油7号’和‘天油4号’萌动种子的ABA、IAA、GA3和ZT的含量。添加Col后,与0℃处理12 h相比,0℃处理24 h时,‘陇油7号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量上调,‘天油4号’α-tubulin、β-tubulin、CBF1表达量上调,但MKK2、ICE1、BrAFP1的表达量呈现下调;与20℃处理12 h相比,20℃处理24 h时,‘陇油7号’和‘天油4号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量均上调。添加Tax后,与0℃处理12 h相比,0℃处理24 h时,‘陇油7号’α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量均下调,‘天油4号’α-tubulin、β-tubulin、ICE1、CBF1、BrAFP1的表达量均上调;与20℃处理12 h相比,20℃处理24 h时,‘陇油7号’和‘天油4号’α-tubulin、β-tubulin、MKK2、ICE1、CBF1、BrAFP1的表达量均下调。表明Col和Tax能使白菜型冬油菜细胞骨架结构发生变化,影响白菜型冬油菜萌动种子内源激素的含量及抗寒基因的表达。
