小麦抗旱性研究进展与展望
2023-05-27毛新国王景一李超男柳玉平景蕊莲
李 龙,毛新国,王景一,李超男,柳玉平,景蕊莲
(中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程,北京 100081)
小麦是全人类的主要粮食作物,2020年全球总产量约7.16×108t,养活了近1/5的人口(FAOSTAT,https://www.fao.org/faostat/en/#data)。然而,为了应对人口增长、疾病侵袭和战争动荡等持久性或突发性问题,全球小麦产量仍需大幅提高[1]。随着气候变化以及耕地面积减少的形势日益严峻,实现这一目标将主要依赖于提高小麦自身生产力[2]。小麦主要分布在干旱和半干旱地区,干旱缺水是影响小麦生产力的主要环境因素。因此,培育并选用抗旱高产小麦品种是抵御干旱、保障粮食安全的重要途径。科学地提高小麦抗旱性需要深入认识以下问题:干旱胁迫对小麦生产力的危害机理是什么?如何从大量的种质资源中鉴定抗旱种质?抗旱种质中蕴含哪些优异基因资源?如何利用抗旱基因资源高效创新小麦种质?这些问题涉及小麦抗旱性改良的物质基础和有效手段,也是长期以来小麦研究的核心问题。本文系统阐述了上述问题的研究现状,并探讨今后有待加强和扩展的重点研究内容,以期为加快培育突破性抗旱高产小麦新品种提供理论参考。
1 干旱对小麦生产的影响
全球大约50%的小麦产区遭受周期性干旱胁迫[3],我国受旱地区也十分广泛。国家统计局最新数据(https://data.stats.gov.cn/%20easyquery.htm?cn=C01)显示,2021年我国旱灾受灾面积为342.6万hm2,约占总受灾面积的29%,旱灾成灾面积140.7万hm2,约占总成灾面积的30%。据农业农村部报道,干旱年份中河南、山东和河北等8省小麦受旱面积约占其冬小麦种植面积的40%;另外,随着全球气候变化,我国黄淮冬麦区和北部冬麦区的干旱形势仍将持续加剧[4]。干旱缺水严重影响小麦产量形成,Daryanto等[5]对1980—2015年发表的144项小麦抗旱性研究结果进行荟萃分析(meta-analysis),发现干旱胁迫导致小麦产量平均下降21%。Zhang等[6]通过分析1980—2017年期间发表的60项研究结果认为,干旱胁迫使得小麦产量平均下降28%,而持续重度干旱造成小麦产量下降幅度可高达92%[7];另外,穗粒数、千粒重、株高等重要产量相关农艺性状下降幅度分别约为30%~50%、15%~20%、10%~35%[8],叶片含水量、光合速率和叶绿素含量等关键生理性状下降幅度分别可达70%、30%和20%[9]。
干旱胁迫可能发生在小麦的各个生长发育阶段(如表1所示),其危害的主要生理途径表现为以下方面:一是改变细胞膜的结构与透性,细胞膜在干旱缺水条件下失去半透性,从而引起胞内氨基酸、糖类物质外渗[18];二是破坏正常代谢过程,如光合作用显著下降或停止[19],呼吸作用增强,使得氧化磷酸化解偶联,能量以热能的形式耗散,使得蛋白质合成过程削弱,分解速率加剧,进而影响核酸代谢,DNA和RNA合成受阻,导致遗传调控紊乱[20];三是水分分配异常,蒸腾作用强烈的功能叶向分生组织或幼嫩组织夺取水分,导致幼嫩组织严重失水,发育不良[21];四是原生质体的机械损伤,干旱胁迫使细胞脱水,向内收缩,破坏原生质体的结构,骤然复水则会引起细胞质与细胞壁的不协调膨胀,从而撕裂原生质,造成组织或器官死亡[22]。此外,不同生育期缺水对小麦产量形成的危害程度不同,开花期和灌浆期对于水分亏缺尤为敏感,主要是由于干旱导致该时期小麦叶片衰老加速,光同化系统受到氧化损伤,碳固定和同化物转运效率降低,花粉不育,胚珠败育,以及库容量下降等[23]。
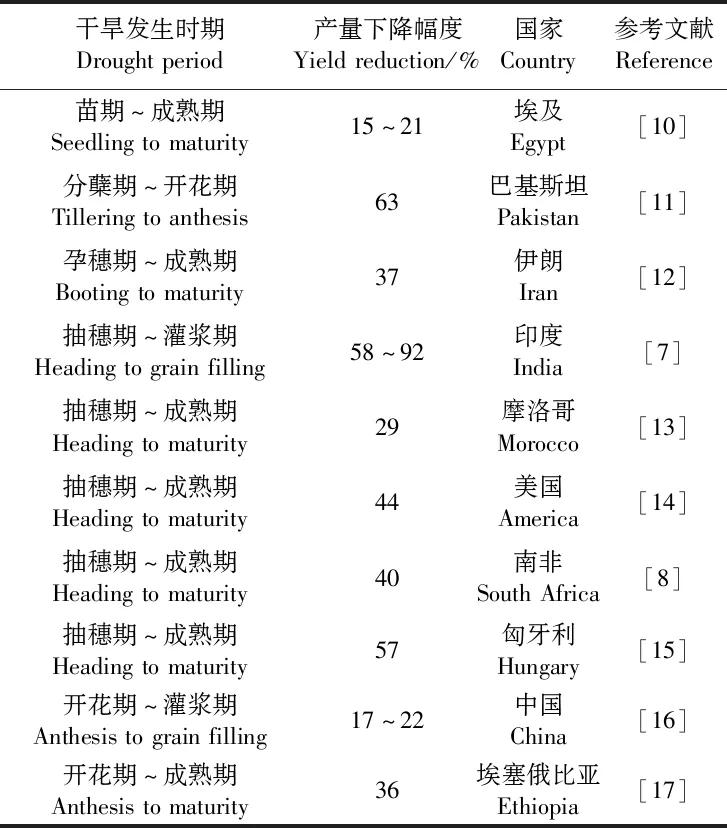
表1 干旱导致小麦籽粒产量的下降幅度Table 1 Reductions in grain yield in wheat caused by drought stress
2 小麦抗旱种质资源鉴定评价
小麦在进化和驯化过程中不断适应环境,形成了形态、生理、遗传特性迥异的多样性种质,其中一些种质表现出强抗旱性,这些种质资源是改良小麦抗旱性的物质基础。多年来国内外学者不断探索,总结出了适用于小麦不同生育期的抗旱性鉴定评价技术体系。芽期主要采用高渗溶液法,通常使用PEG-6000和甘露醇等高渗透势溶液模拟干旱环境,检测发芽率、发芽势、发芽指数及活力指数等指标来比较不同材料的萌发能力。任毅等[24]利用该方法从301份冬小麦种质中筛选到‘川麦44’、‘皖麦33’及‘藁城8901’等芽期抗旱种质。该方法具有重复性好、操作简便等优点,但是鉴定结果难以准确反映小麦在复杂田间环境下的抗旱性表现。苗期主要采用反复干旱法,测定指标为反复干旱存活率,即多次干旱处理后幼苗存活率的平均值。早在20世纪前叶,前苏联研究人员就利用该方法鉴定了上万份小麦种质,并建立了小麦抗旱基因库[25]。通过实施反复干旱,可以使不同种质的细胞膜透性、叶片水势及酶活性等性状差异凸显,从而有效考量小麦种质在不同干旱胁迫下的存活能力。该方法也具有较好的重复性,通过合理的土壤组分配比及试验设计能够有效控制环境效应,目前已经广泛应用于各类粮食及经济作物[26-27],但该方法需要旱棚等特殊设施支持。成株期主要采用田间鉴定法,通过设置水、旱两种处理,重点考量逆境条件下不同种质最终的生产能力,测定指标一般为产量及其构成因素。田间鉴定在降水量较低的地区进行,旱处理一般为雨养条件,水处理进行灌溉,优点是鉴定条件接近生产实际,不过该方法受环境影响较大,尤其是不同年际间的降水量重复性较低,实施前应注重试验地点的选择和试验设计的合理性[28]。2007年颁布的国家标准《小麦抗旱性鉴定评价技术规范》(GB/T 21127-2007)中明确了相关试验的条件要求,为种质资源评价和遗传改良提供了科学、准确、可操作性强的规范化标准,已被国家农作物品种审定委员会和育种单位广泛采用,加速了全国作物抗旱性鉴定评价的标准化进程,提高了优异种质资源的共享水平和育种效率。
随着作物抗旱性研究仪器设备和分析方法的不断升级换代,目前能够检测的性状种类更加丰富,因此,抗旱性评价指标的选择也引起了广泛的关注。早期主要是基于发芽率、存活率或产量,通过抗旱系数或抗旱指数进行单指标评价,随后引入了隶属函数法,而后基于主成分及灰色关联度分析方法形成更为复杂的综合评价方法,旨在提高种质资源鉴定的客观性和准确性[29]。祁旭升等[30]发现抗旱性综合评价值与单指标评价值高度相关,但少量种质出现了跃级的现象,此外,从汪灿等[31]的鉴定结果来看,不同指标抗旱系数与综合评价值存在较大差异。本研究组对单指标评价方法与综合评价方法的鉴定结果进行比较分析,认为在育种工作中,如果仅通过单指标筛选亲本,可能导致育种后代产量提升潜力较小,而根据综合指标筛选优异种质进行杂交,有望聚合更多的优异基因,从而提高育种效率[32]。因此,在实际应用中,应综合考虑鉴定评价的目的以及不同鉴定方法的准确性和工作量,选择合适的鉴定评价方法和指标,例如在进行大规模种质抗旱性评价工作中,可采用单指标评价方法,而针对优异亲本材料或品种的遴选工作,应将单指标分析与综合指标分析方法结合运用。
3 小麦抗旱种质创新
基于抗旱性鉴定评价结果,选择优异抗旱种质作为亲本进行杂交,聚合抗旱优异等位基因,是培育抗旱新品种的基本策略。国际玉米小麦改良中心(CIMMYT)长期致力于小麦抗旱种质创新和品种选育,利用野生近缘种、地方品种和育成品种中筛选到的‘Aragon03’、‘Pastor’、‘CATBIRD’、‘Wariquam’、‘KALYANSONA’等优异种质进行杂交,相继培育出‘Babax’、‘SeriM82’、‘Borlaug100’、‘Opata M85’、‘Gladius’等一批突破性的抗旱高产小麦品种,并分享给不同国家和地区种植或育种利用,不仅提高了世界干旱半干旱地区的小麦生产力,同时也显著提高了全球旱地小麦育种水平[33-34]。此外,CIMMYT的研究人员利用四倍体小麦(TriticumturgidumL.)与二倍体粗山羊草(AegilopstauschiiCoss.)杂交创制了1577份人工合成六倍体小麦,极大地拓宽了现代小麦的遗传多样性,为利用小麦野生祖先种的优良基因搭建了桥梁[35]。目前,利用这些人工合成小麦种质与来源于20多个国家的小麦育成品种进行杂交,已经相继育成86个高产耐逆小麦新品种,为全球旱地小麦生产贡献了30%的产量增益[36]。
我国也始终将抗旱性改良作为小麦育种工作的中心任务之一,并取得了丰硕成果。例如,四川省农科院从CIMMYT引进大量人工合成六倍体小麦,通过与本地品种杂交,培育出了‘川麦42’、‘川麦43’、‘川麦48’等一系列高产广适小麦品种[37]。中国农业科学院牵头带动全国小麦工作者历时近40年对2.4万份小麦种质进行了抗旱性鉴定,遴选出强抗旱种质110多份,利用这些种质培育出了‘晋麦47’、‘长6878’及‘洛旱7号’等一批抗旱节水小麦新品种[38],其中,‘晋麦47’具有适应性广、稳产高产、低投入高产出等优点,被连续选为国家黄淮冬麦区旱地区域试验对照品种长达22年(1998-2020年)[39]。国家黄淮冬麦区旱地区域试验是我国优选抗旱高产小麦品种的主要平台,于2005年被进一步细分为黄淮旱肥、旱薄两组区域试验,旱薄组区域试验主要面向抗旱耐瘠薄小麦品种选育。截至目前,国家黄淮冬麦区旱薄组区域试验已推选出‘洛旱13’、‘西农219’和‘中麦36’等22个抗旱耐瘠高产小麦品种(表2),这些品种的推广种植大幅提高了我国小麦中低产田的生产水平。

表2 2005—2022年国家区域试验黄淮冬麦区旱薄组推选品种Table 2 Varieties approved by the national trial of arid and infertile group in Huang-Huai River Valleys facultative wheat zone from 2005 to 2022
‘中麦36’是以‘晋麦47’为受体亲本,遗传远缘的抗逆稳产品种‘陕优225’为供体亲本,通过3次回交和自交,在多生态环境中选择出的抗旱高产稳产广适小麦新品种,与山西省农业科学院棉花研究所合作育成[40]。该品种自2017年参加生产试验以来,连续4年(2017—2020年)被农业部推介为黄淮冬麦区旱地展示示范新品种,并于2020年接替‘晋麦47’成为国家黄淮冬麦区旱薄组区域试验对照品种,为我国旱地小麦品种评价利用树立了新标杆。‘中麦36’生产试验平均产量5 563.5 kg·hm-2,比对照品种‘晋麦47’增产12.6%,株高降低12 cm左右,茎秆基部节间较短,穗下节较长(约占株高的1/3),重心下移,使得抗倒性显著增强(图1a、b);在干旱条件下,‘中麦36’有效穗数和穗粒数的稳定性表现尤为突出,有效穗数较对照品种增加3.7万,同时穗粒数和千粒重与对照品种相当;此外,分蘖期和拔节期的根深均显著大于‘晋麦47’(图1c、d)。从基因组水平来看,‘中麦36’的优良性状得益于‘晋麦47’和‘陕优225’优异等位基因的互补与聚合,‘中麦36’不仅保留了其轮回亲本‘晋麦47’中绝大多数优异等位基因,同时还导入了59个‘陕优225’基因组中与株高(18个)、有效穗数(4个)、千粒重(7个)、小区产量(8个)、冠层温度(9个)、根深(13个)相关的优异等位基因,这些优异等位基因在小麦表型塑造中具有显著的累加效应,这意味着通过分子育种策略科学选择和聚合优异等位基因是进一步加快小麦抗旱性遗传改良的有效途径。

图1 ‘晋麦47’与‘中麦36’表型比较Fig.1 Comparison of phenotypes between ‘Jinmai 47’ and ‘Zhongmai 36’
4 小麦抗旱基因资源发掘与利用
长期以来,小麦优异等位基因的发掘效率远远滞后于水稻、玉米等主要农作物,其主要原因在于小麦为异源六倍体作物,基因组庞大(15.4~15.8 Gb)且结构复杂,序列注释难度高[41-42]。随着小麦参考基因组信息的日趋完善,以及表型与基因型鉴定技术的快速发展,小麦优异基因资源发掘和分子标记开发效率大幅提升[43-44]。目前,国内外研究者已发掘到一批小麦抗旱相关基因,并对其作用机理进行深入解析(表3)。例如,Mega等[45]发现ABA受体基因TaPYL4能够提高小麦对CO2的固定能力和羧化速率,从而加强光合活性和抗旱性。Bi等[46]克隆到1个AP2/ERF家族基因TaSHN1,该基因通过降低叶片的气孔密度和失水率,增加表皮蜡质中的烷烃含量,从而提高小麦抗旱性。在我国,康振生团队利用不同地理来源的小麦自然群体,通过全基因组关联分析发掘出小麦抗旱基因TaNAC071-A,并发现MYB转录因子TaMYBL1可结合并调控TaNAC071-A基因表达,从而导致TaNAC071-A表达量变化和小麦抗旱性差异[47]。马有志团队从强抗逆大豆品种‘铁丰8号’中克隆了转录因子基因GmTDN1,发现该基因可协同提高小麦的抗旱性和氮肥利用效率,为同步改良小麦抗旱节水和氮肥利用效率开辟了新途径[48]。本课题组通过结合全基因组关联分析、连锁作图及集群分离分析等方法鉴定到TaSnRK2.10、TaMYB5和TaERF73等关键小麦抗旱基因,其中,TaSnRK2.10能够磷酸化TaERD15和TaENO1,调控DREB2s、HSFs、HSPs、Dehydrins、LEAs等干旱应答基因的表达模式,参与协调小麦在干旱条件下维持较低的基础代谢水平,从而提高抗旱性[49-51]。目前我们已基于这些基因的启动子或编码区序列变异开发了多种类型的分子标记(CAPS、dCAPS、KASP)[52-54],利用这些标记可以对根系、农艺、生理等抗旱相关性状的优异单倍型或等位基因进行精准选择,从而有效提高小麦抗旱基因型鉴定的准确性。
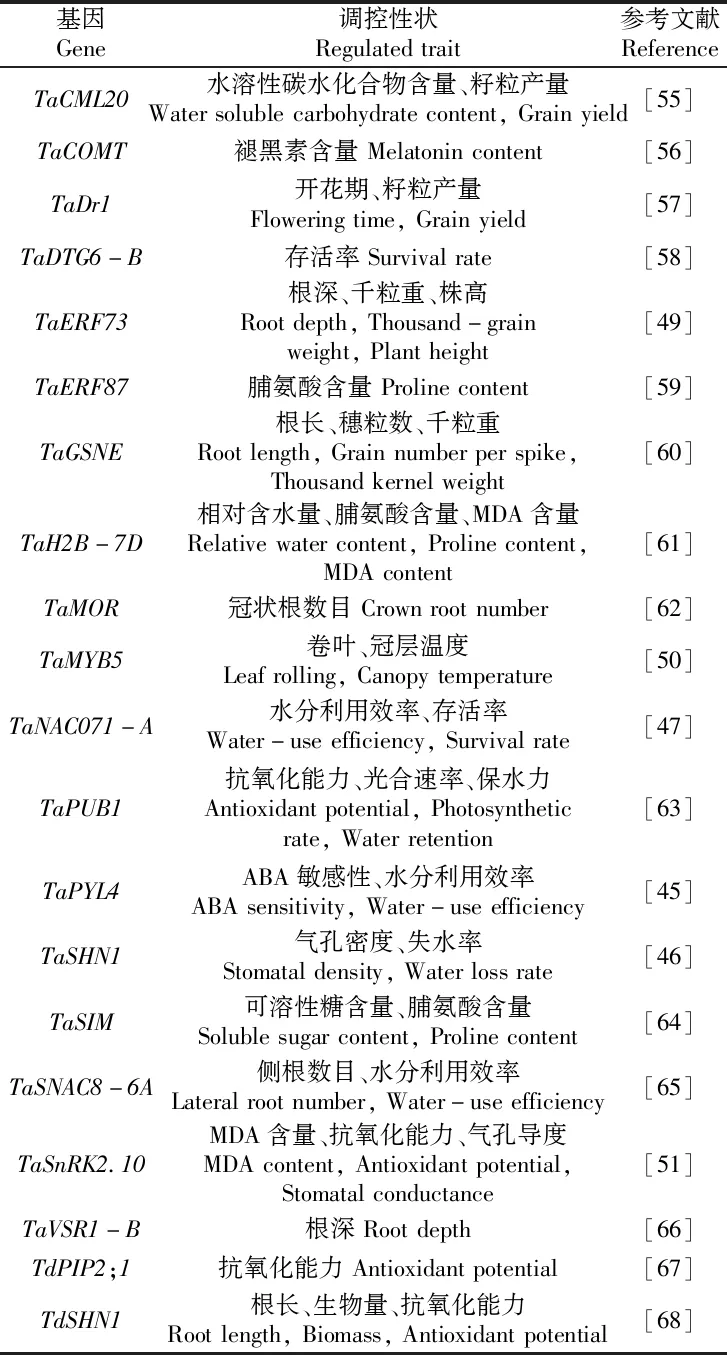
表3 近5年报道的小麦抗旱相关基因Table 3 Drought-resistant genesin wheat reported in the last five years
小麦抗旱性是复杂的数量性状,由分布在庞大基因组上的众多基因协同调控,如果不考虑环境影响以及基因间的连锁与互作效应,仅仅对单个或少数优异等位基因进行选择,难以高效改良小麦抗旱性,这也是目前抗旱基因理论研究与育种实践相脱节的主要问题[69]。因此,如何有效地应用优异抗旱基因资源是目前小麦育种面临的重大科学问题和难点。多组学(基因组、转录组、代谢组、表型组)技术与机器学习的深度融合为抗旱基因资源的高效利用提供了新的契机[70]。例如,王向峰团队与马闯团队利用玉米遗传材料的系谱关系和表型数据模拟育种过程,提出“基因组优化设计”策略,即通过模型算法模拟出理论上尽可能多的聚合了优异等位基因的虚拟基因组,育种家可根据自交系中含有的虚拟基因组中优异等位基因个数选择优良品系,也可以参考虚拟基因组信息选择优异等位基因互补的自交系进行杂交,从而利用最少的遗传材料、最短的育种周期,最大限度地将优异等位基因聚合于后代材料中[71]。李慧慧团队提出了利用作物海量多组学数据进行全基因组选择的深度学习策略(DNNGP),即通过构建预测模型,根据基因组估计育种值进行早期个体的预测和选择,从而缩短育种世代间隔,该策略能够针对具有复杂结构的多组学海量数据进行数学建模,实现了育种大数据的高效整合,为深度利用基因资源和表型信息实现“经验育种”到“精确育种”的转化提供了有效工具[72]。目前,这些新的分子育种策略已经逐步由理论研究阶段转向实际应用阶段,也必将有效提高优异抗旱基因资源的应用水平。
5 问题与展望
过去几十年,我国针对小麦抗旱性的科学研究沿着从种质资源评价到基因发掘、再到种质创新和利用的思路不断推进,助力我国旱地小麦连年增产,已取得显著成效[73]。然而在面对人口增长和环境变化的双重压力下,我们仍需加速提升旱地小麦生产力水平,向着更高的育种目标奋进,这将有赖于优异种质、基因资源、育种策略三方面的创新发展(图2)。
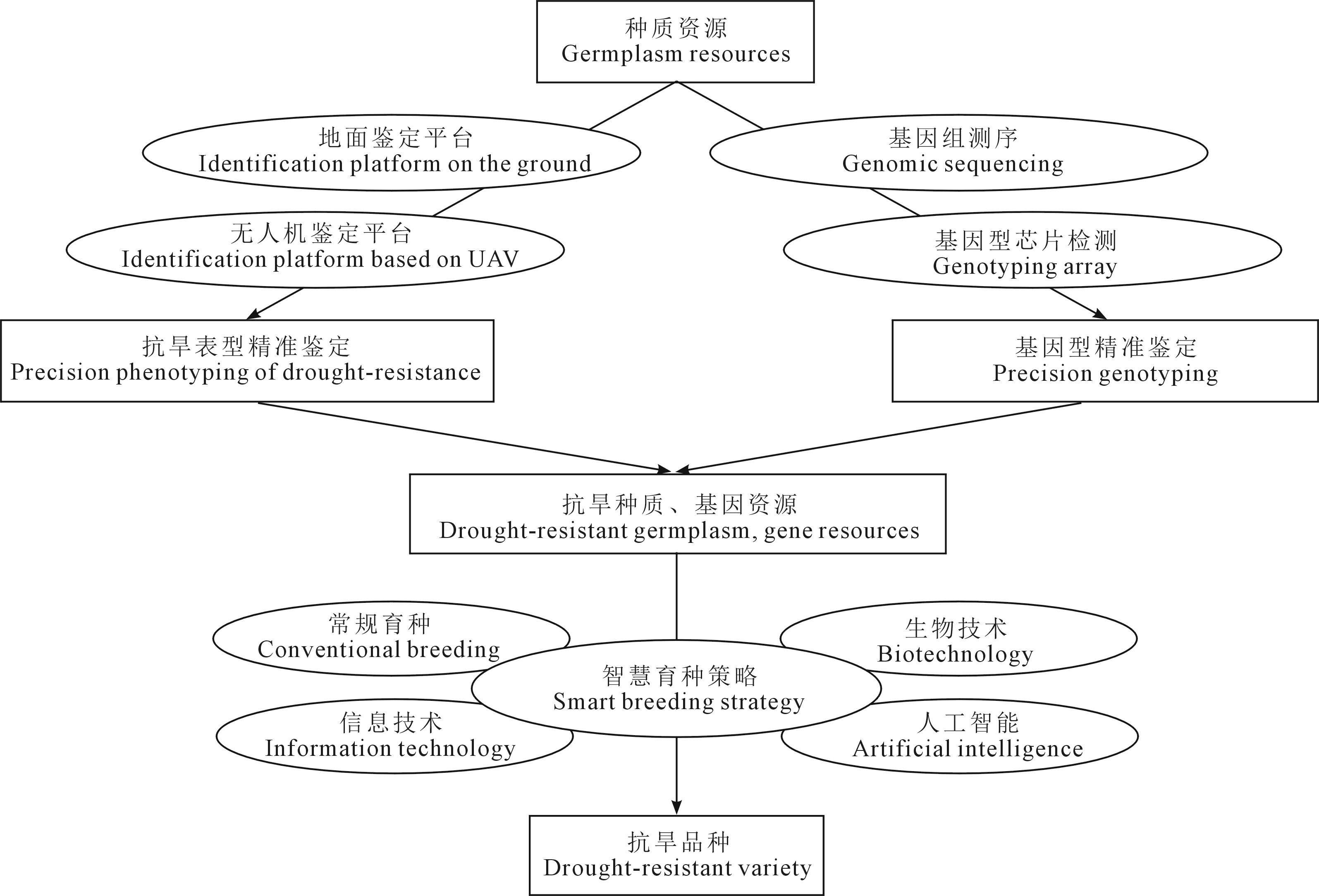
图2 小麦抗旱性改良策略框架图Fig.2 Framework of drought resistance improvement strategy in wheat
5.1 建立基于高通量表型鉴定的抗旱性综合评价技术体系
在抗旱种质鉴定评价方面,需同步提升鉴定效率和精度,即实现“综合鉴定”和“高效评价”,这就要求我们研发高通量表型鉴定技术及其配套条件。一方面应建立地面高通量表型检测平台,将光电技术、自动化控制技术和计算机图形处理技术集于一体,通过光、温、水(湿度)、CO2等不同的环境感应器实现作物抗旱性表型参数全自动、无损伤、高通量准确提取,例如法国农业科学院、澳大利亚植物功能基因组中心,以及华中农业大学作物表型中心均建立了高通量表型观测温室系统,通过该系统已经完成了在人工控制环境条件下大量作物种质的抗旱性鉴定,发掘出了一批强抗旱种质[74]。另一方面是发展无人机表型观测平台,通过无人机搭载可见光成像传感器、热红外成像传感器、多光谱成像传感器、高光谱成像传感器、激光雷达成像传感器等新型光温图像采集设备,快速获取田间的作物表型参数,减少环境和人工检测误差的影响。同时,应进一步加强地面和无人机表型鉴定平台技术与数据的有机整合,从不同视角尽可能多的获取不同水分环境下的小麦形态和生理指标,利用大数据分析方法建立综合评价体系,全面高效准确地评价小麦种质的抗旱性[75]。
5.2 建立基于高通量基因型鉴定的抗旱基因资源发掘平台
自1977年Sanger发明双脱氧法测序至今,基因测序技术经历了三次大变革,目前已经能够对规模化种质全基因组进行低成本、高通量、高精度测序,并构建DNA指纹图谱、单倍型图谱和泛基因组[76]。例如,通过对3010份亚洲栽培稻基因组测序,剖析了水稻核心种质资源的基因组遗传多样性,推动了水稻规模化基因发掘和复杂性状的分子改良,提升了全球水稻分子育种水平[77]。对2898份大豆基因组测序以及26份代表性大豆品种高质量基因组组装,构建了大豆泛基因组图谱,极大地促进了大豆功能基因组学研究[78]。目前在小麦中也完成了‘中国春’、‘藏1817’和‘Fielder’等小麦品种的基因组测序与组装,获得了多个高质量参考基因组,同时对多个小麦群体进行了简化基因组测序,初步建立了小麦基因组变异数据库,为高效发掘小麦优异基因提供了丰富的基因组信息[41,79-81]。基于此,我国的小麦研究工作者已经创建了小麦基因资源发掘平台,如小麦基因定位与基因组研究平台整合了超过3500份小麦的全基因组、外显子组和转录组数据,兼容四种集群分离分析模型,有助于研究者利用集群分离分析方法定位并克隆小麦抗旱及其它复杂性状调控基因[82]。未来随着小麦基因组变异信息的不断扩充以及泛基因组的构建完成,高通量小麦基因资源发掘平台建设将更加完善,通过搭载更多的抗旱相关表型与基因型关联分析模型,将推动小麦抗旱基因资源高效发掘。
5.3 创建基于综合运用多学科技能的智慧育种策略
先进的育种策略是将种质、基因和表型数据信息资源有机整合,为作物生产力的提升提供重要理论支撑。进入21世纪以来,随着互联网、机器学习、人工智能等现代信息技术的迅猛发展,全球已迈入大数据时代[83]。信息科学与生命科学的交叉融合为育种策略的创新发展提供了新的契机。“一把尺子一杆秤、用牙咬、用眼瞪”的常规育种体系正在大跨步走向“常规育种+生物技术+信息技术+人工智能”的智慧育种模式[84]。2018年,美国玉米遗传育种学家Buckler教授提出“育种4.0”概念,我国水稻遗传育种学家张启发教授也提出了“5G育种”概念,其本质都是育种策略的智能化、工程化升级[85-86]。实际上,美国依据过去几十年所积累的海量育种数据,已经可以进行作物表型的模拟,每一个育种环节都能够通过大数据驱动,辅助育种家筛选材料进行杂交组配,可以说基本进入了“育种4.0”时代。虽然我国整体育种模式与之相比仍有差距,但在生物技术和数据模型分析等科学研究方面已跻身世界先进行列。例如,高彩霞团队在基因编辑方面取得了一系列重大突破,其中多重“基因剪刀”实现了对小麦基因序列的精准操控[87]。王向峰团队与严建兵团队合作研发的LightGBM工具箱整合了育种材料遗传结构解析、标记筛选与模块设计、全基因组选择模型、数据可视化等功能模块,可以为作物基因组设计育种提供一站式理论指导[88]。因此,我们今后应着重加强生物信息学、群体遗传学、基因编辑、大数据、智能计算等多学科的综合应用,创建系统实用的智慧育种策略,为未来小麦抗旱性等复杂性状的遗传改良提供技术支撑。
总之,通过对种质资源的精准鉴定评价,基因资源的高效发掘利用,以及育种方法的集成创新,将整体推进小麦抗旱性理论研究向育种实践的转变,进一步发挥小麦的抗旱增产潜力,为在全球气候变化背景下保障国家粮食安全做出新贡献。
