白杨素衍生物通过DAF-16 调节线虫抵御细菌感染能力的研究
2023-05-19罗路陈梦婷陈映之周俊杰杨宝盈冯娜
罗路,陈梦婷,陈映之,周俊杰,杨宝盈,冯娜
(五邑大学 生物科技与大健康学院,广东 江门 529020)
白杨素(Chysin)是一种从紫葳科木蝴蝶种子里分离提纯得到的黄酮类化合物,具有抗氧化、抗菌、抗炎、抗焦虑、治疗糖尿病以及抗癌等广泛药理作用[1-4]. 白杨素是性能良好的抗氧化剂,能有效清除自由基[5]. Vaya 等[6]通过对白杨素的研究发现白杨素在体外能够清除AAPH 分解的阳离子型自由基,还能减少铜离子的氧化. 白杨素及其衍生物具有抑菌、免疫调节等活性. Patel 等[7]采用丁基链与哌嗪、吗啉和哌啶等取代合成的白杨素衍生物具有清除DPPH 和ABTS 自由基的能力,并且能抑制癌细胞的增殖. Zhu 等[8]合成的白杨素-β-D-吡喃半乳糖苷对H22 细胞系的活性抑制,和对羟基自由基、DPPH 自由基、超氧阴离子的清除能力,以及对细菌和真菌的抑制作用均比白杨素强.
近几十年,已有很多学者开始将秀丽隐杆线虫(Caenorhabditis elegans,以下简称线虫)作为宿主模型用于体内病原微生物的毒力及致病机制的研究. 在线虫中,天然免疫反应一方面是通过释放杀菌物质直接杀死病原微生物;另一方面是通过触发天然免疫信号通路,调节宿主体内生理平衡,保护自身免受病原微生物感染的危害. 最近的研究结果显示,活性氧(reactive oxygen species,ROS)参与了上述两方面[9]. ROS 是一类含有氧原子的自由基和非自由基的分子或离子化合物的总称[10].Malhotra 等[11]发现生物体内的ROS 水平与囊肿纤维化(CF)肺部疾病有关;Li 等[12]发现感染肠病毒71 型(EV71)会诱导不同细胞系的凋亡,同时细胞会产生大量ROS,而外源添加NAC 可降低EV71 感染细胞的凋亡和炎症水平,减少EV71 的增殖,这些结果表明ROS 水平与机体的感染和恢复有关.
白杨素及其衍生物的抗氧化活性及免疫调节作用提示其在抗菌方面具有有益作用,因此针对白杨素的衍生化有望为发展新型抗菌剂提供思路. 本文利用线虫-病原菌感染模型对白杨素衍生物在天然免疫中的作用进行探讨,以期为了解动物抗感染机制、开发新的抗感染策略提供帮助.
1 实验方法
1.1 线虫株系和菌株
线虫N2 (Bristol)、TJ356 zIs356 [daf-16p::daf-16a/b::GFP+rol-6(su1006)]、CF1553 muIs84 [(pAD76)sod-3p::GFP+rol-6(su1006)]和菌株E.coliOP50、E.faecalis均由兰州大学支德娟老师惠赠,菌株S.aureus、Proteus由五邑大学魏萍老师惠赠,P.aeruginosaPA14 由云南大学邹成钢老师惠赠.
1.2 白杨素衍生物体外抗氧化活性检测
参考文献[13]的方法对 DPPH 自由基清除能力进行检测. 吸取不同浓度的待测样品100 μL 和0.1 mM DPPH 乙醇溶液100 μL 于96 孔板,摇匀,在室温避光放置30 min,在517 nm 处测定吸光值记为At;取不同浓度的待测样品100 μL 和 100 μL H2O,测定其吸光值记为Ar;吸取100 μL DPPH 和100 μL H2O,测定其吸光值记为A0. 按以下公式计算DPPH 自由基清除率:
参考文献[14]的方法对ORAC(总抗氧化能力)进行检测,在96 孔板中加入50 μL 待测样品,加入50 μL 75 mM 的PBS 溶液代替样品作为空白组,以及50 μL VE 溶液代替样品作为标准组,避光加入100 μL FL 工作液,放入酶标仪进行检测(激发波长485 nm,吸收波长535 nm),初始荧光值记为0F;37°C 温育箱振荡3 min 后继续温育10 min,迅速加入50 μL 153 mM 的AAPH 溶液启动反应,放入酶标仪检测孔动力学,检测过程中每隔150 s 测定一次荧光值记为nF,直至荧光值衰减至直线.按以下公式计算标准曲线和总抗氧化能力指数:
VE 的浓度作为X轴,NetAUCVE作为Y轴绘制Trolox 标准曲线,将 NetAUCSample代入Trolox 的线性方程中进行计算,结果以VE 的当量mM 来表示,得到样品的ORAC 值,即为样品的总抗氧化能力指数,记为mM TE/g DW.
1.3 96 孔板法测定白杨素衍生物的抑菌率
参考文献[15]的方法,将复苏的菌株用MHB 培养基培养至F2 代菌,再用MHB 稀释菌液得到与0.5 单位麦氏比浊液OD610 相当的菌液,定义为细菌浓度 1.5 ×108CFU/mL ,再用MHB 将菌液100×稀释得到最终浓度 1.5 ×106CFU/mL 的菌悬液. 然后吸取50 μL 待测样品、50 μL 菌悬液、50 μL MHB 于96 孔板,立刻置于多功能酶标仪上在600nm条件下测定其吸光度,记为OD0,然后将96孔板置于37°C恒温培养箱培养12h后在相同波长条件下测定吸光度,记为OD12.其中以MHB10×稀释的DMSO 代替样品作为阴性对照. 按以下公式计算抑菌率:
1.4 PA14 Killing 实验
PA14 Killing 实验参考文献[16]的方法设计,挑取30 条大小形态相似的不同处理的L4 期成虫至PA14/NGM 板上,然后置于25°C 培养,每天对线虫的存活进行评定,将活着的线虫转移至新的PA14/NGM 板上,直到全部死亡为止,按以下公式对线虫的存活率进行计算:
1.5 线粒体活性氧检测
参考文献[17]的方法设计实验,将不同处理的 L4 期线虫用 M9 缓冲液洗 3 次,再用终浓度1 μM MitoSOXTMRed 于20°C染色20 min,然后用M9 缓冲液洗3 次后加入适量 NaN3麻痹线虫,用琼脂糖进行封片,于BX63 生物荧光显微镜(正置)下选择RFP 模式进行观察,并摄取图片,用ImageJ量化分析.
1.6 荧光定量PCR
按照总RNA 提取试剂盒说明书[18]提取不同处理的线虫RNA,然后配制Master Mix 混合RNA样品在PCR 混合仪上分别进行Step1(42°C ,2 min)和Step2(85°C ,5 s)两步反应,逆转录成稳定的cDNA,然后将Mix 和cDNA 样品溶液混合,置于PCR 扩增仪上,程序如下:
Stage 1:预变性(95°C ,30 s)
Stage 2:PCR 反应40 Cycles(95°C ,3 s;60°C ,30 s)
1.7 突变株线虫转基因荧光可视化及荧光定量分析
本文使用TJ356 和CF1553 两种突变株线虫分别进行DAF-16 核定位实验和SOD-3 蛋白表达分析. 实验步骤如下:将不同处理的L4 期线虫用M9 缓冲液洗3 次后加入适量 NaN3麻痹线虫,用琼脂糖进行封片,于BX63 生物荧光显微镜(正置)下选择GFP 模式进行观察,并摄取图片,用ImageJ量化分析.
2 结果分析
2.1 白杨素衍生物能有效清除自由基
体外测定抗氧化活性实验具有高效、简便、快捷等优点[19],适用于大量化合物的筛选. 对图 1所示的白杨素及13 种白杨素衍生物(a -m)的抗氧化活性进行研究,分别采用DPPH 法和ORAC法进行评价,结果如表1 所示:对照VC 对0.1 mM DPPH 自由基的IC50为12.47±0.78μM ,而白杨素及其衍生物对DPPH 自由基的IC50都比VC 的高(p<0.001),这表明,白杨素衍生物对DPPH 自由基的清除能力相对较弱;其中衍生物h 对0.1 mM DPPH 自由基的IC50值最小,衍生物c 的最大.

图1 白杨素及其衍生物(a-m)结构式

表1 白杨素及其衍生物(a-m)对0.1 mM DPPH 自由基的IC50 值
体外抗氧化活性实验结果如图2 所示:白杨素及其衍生物的总抗氧化能力指数均低于VC(55.47±4.84) mM TE/g DW,p<0.001);50 μM 衍生物h 表现出与白杨素相当的抗氧化能力;衍生物h(4-甲氧基苄溴白杨素醚)的总抗氧化能力指数最高(28.79 ±3.75)mM TE/g DW ),比相同浓度下白杨素的总抗氧化能力指数(23.10 ±6.40) mM TE/g DW)高24.63%;其余12 种白杨素衍生物总抗氧化能力指数均比白杨素低(p<0.001).

图2 白杨素及其衍生物(a-m)的总抗氧化能力指数
2.2 白杨素衍生物 h 能抑制细菌生长,但杀菌能力较弱
研究表明,白杨素具有良好的抑菌活性[8],为了评价白杨素衍生物h 的抑菌活性,分别检测了100、200、400 μM 衍生物h 对金黄色葡萄球菌(Staphylococcus aureus)、粪肠球菌(Enterococcus faecalis)、普通变形杆菌(Proteus)和铜绿假单胞菌PA14(Pseudomonas aeruginosaPA14)的体外抑菌效果,结果如图3 所示(图中所有数据均为(平均值±SEM)):1)图3-a 和3-c 中,400 μM 衍生物h 对金黄色葡萄球菌和普通变形杆菌的抑菌率分别为(76.32 ±11.16)%、(72.95 ±11.56)%,均大于70%,说明高浓度衍生物h 能抑制细菌的生长;而中低浓度衍生物h 对金黄色葡萄球菌和普通变形杆菌均没有明显的抑制作用. 2)图3-b 和3-d 中,不同浓度衍生物h 对粪肠球菌和铜绿假单胞菌PA14 的抑菌率均小于70%,表明衍生物h 不具有抑制粪肠球菌和铜绿假单胞菌PA14 生长的作用.

图3 白杨素衍生物h 对不同细菌的抑制率
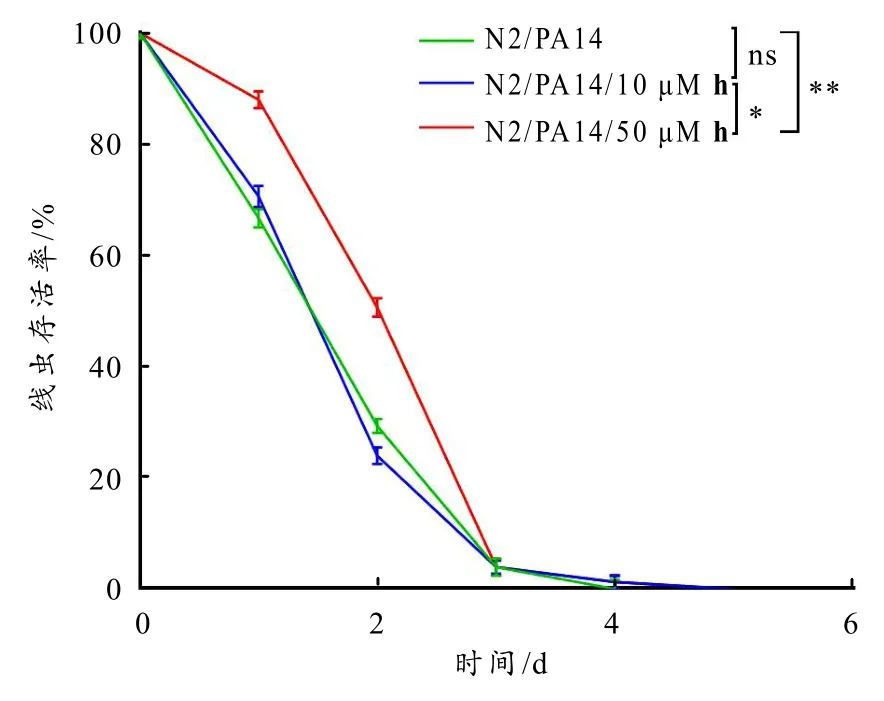
图4 白杨素衍生物h 对PA14 感染后线虫存活率的影响
体外抑菌实验结果表明,衍生物h 对粪肠球菌和铜绿假单胞菌PA14 没有明显的活性抑制,鉴于它的抗氧化活性,我们推测衍生物h 可能通过免疫调节途径来增强宿主的抗感染能力.
2.3 白杨素衍生物h 延长了PA14 感染后线虫的寿命
研究表明,铜绿假单胞菌PA14 可以感染线虫,该病原—宿主模型广泛用于研究宿主天然免疫与细菌感染的关系. PA14 感染后的线虫分别用10 μM 和50 μM 衍生物h 处理或不处理,其存活情况如图 4所示(图中所有数据均表示为(平均值 ±SEM),使用t检验计算p值,*p<0.01,**p<0.001,ns 无显著性差异):在感染PA14 的情况下,未处理组线虫平均生存时间为(2.00 ± 0.14) d ;10 μM 衍生物h 处理组线虫平均生存时间为(2.13 ± 0.17) d ,比未处理组线虫寿命延长了6.50%(无统计学差异,ns);50 μM衍生物h 处理组线虫平均生存时间为(2.44 ± 0.05) d ,比未处理组线虫寿命延长了22.00%(p<0.001). 该结果表明,10 μM 衍生物h 不能显著延长PA14感染线虫的寿命,而50 μM 衍生物h 处理后,显著提高PA14 感染线虫的寿命.
2.4 白杨素衍生物h 部分恢复PA14 感染导致的线粒体活性氧水平的降低
ORAC 实验结果表明白杨素衍生物h具有一定的抗氧化活性,已有研究表明宿主体内的ROS 水平对宿主抵御细菌感染的能力有影响[20]. 本文检测了衍生物h对PA14 感染后线虫体内MitoROS 水平的影响,结果如图5 所示(图中所有数据均为(平均值 ±SEM),使用t检验计算p值,与N2 比较,##p<0.001;与N2/PA14 比较,p<0.01).
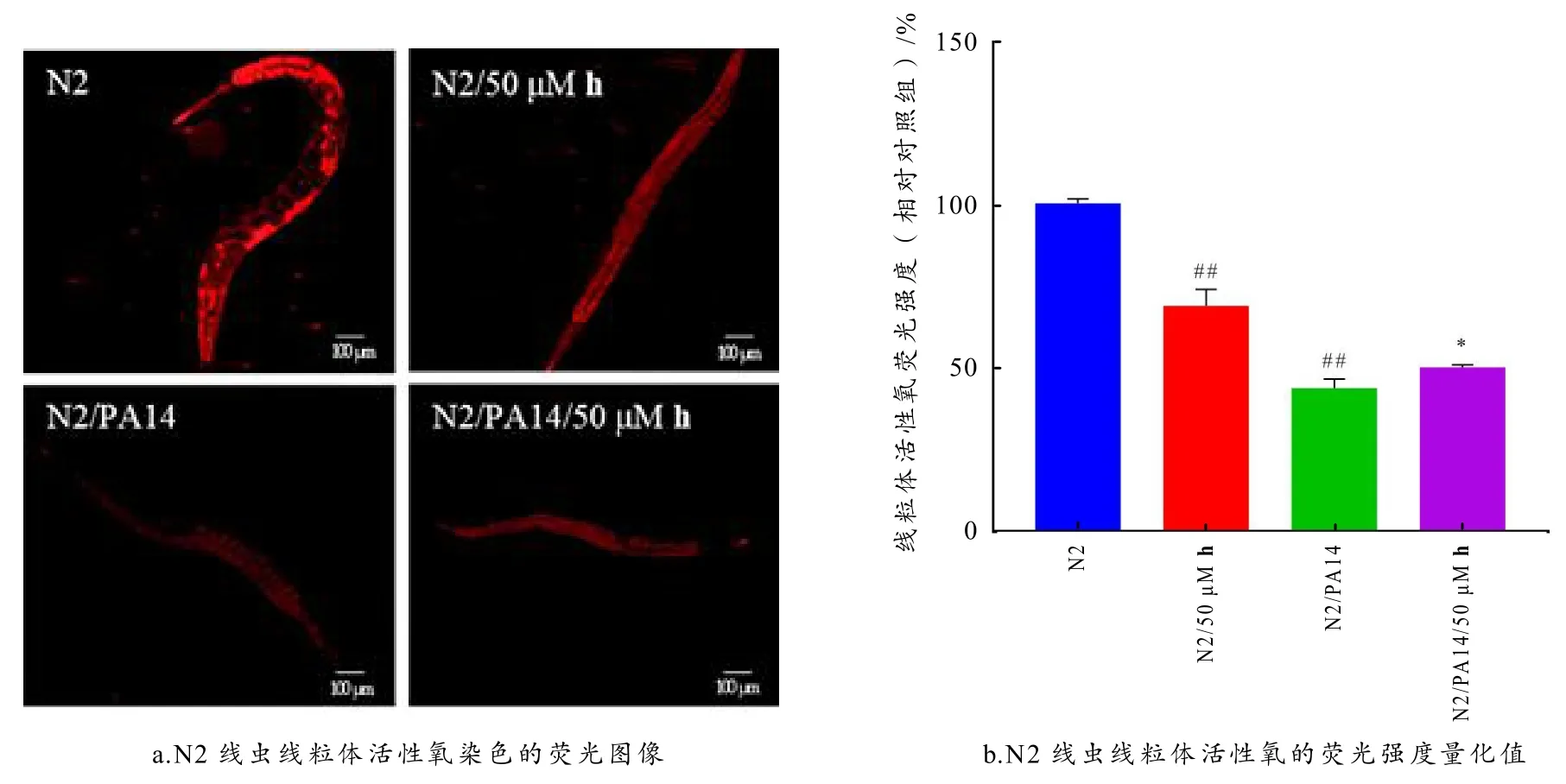
图5 白杨素衍生物h 对PA14 感染后N2 线虫线粒体活性氧水平的影响
如图5-b 所示:与未处理N2 组相比,50 μM 衍生物h处理后线虫ROS 水平显著降低(p<0.001),PA14 感染组的线虫ROS 水平也显著降低(p<0.001),PA14 感染后的线虫经50 μM 衍生物h处理后ROS 水平与PA14 感染组相比显著上调(p<0.001),部分恢复了PA14 感染导致的ROS 降低. 有研究表明,铜绿假单胞菌有规避先天免疫防御的能力,它通过清除中性粒细胞来规避先天免疫反应中ROS 的产生[21]. 在本文中,衍生物h能降低宿主的ROS 水平,这说明衍生物h具有抗氧化活性,与体外的抗氧化实验结果一致;而PA14 感染后线虫的ROS 水平进一步降低,给药处理后ROS 水平显著回升,这可能是衍生物h对宿主的ROS 水平具有调节作用,ROS 作为信号分子诱导了宿主免疫反应,调动下游效应因子增强宿主抵御PA14 感染的能力.
2.5 白杨素衍生物h 上调PA14 感染后线虫的免疫相关基因
DAF-16/FOXO 信号通路参与介导线虫的先天免疫应答,DAF-16 核转位启动免疫相关基因的表达,以防御病原体感染并延长线虫的寿命[22]. 金属硫蛋白同系物mtl- 1和线粒体超氧化物歧化酶基因sod- 3作为DAF-16 的下游基因,参与调节宿主的免疫功能[23].
为了进一步研究白杨素衍生物h在线虫体内发挥抗感染作用的机制,对线虫体内氧化应激相关基因(sod- 1、sod- 2、sod- 3、sod- 4、sod- 5)与抗菌相关基因(mtl- 1)的表达量进行检测,分析衍生物h对线虫体内DAF-16 及其下游靶基因的影响,结果如图6 所示(图中所有数据均为(平均值 ±SEM),使用t检验计算p值,与空白组比较,#p<0.01,##p<0.001;与PA14 组比较,p<0.01,p<0.001).

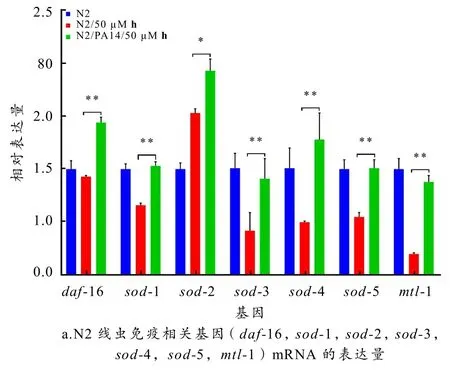
图6 白杨素衍生物h 对PA14 感染后线虫免疫相关基因、DAF-16 核转位和SOD-3 蛋白表达量的影响
如图6-a 所示,与阳性对照N2/PA14 组相比,50 μM 衍生物h处理后线虫的daf- 16(p<0.001)、sod- 1(p<0.001)、sod- 2(p<0.01)、sod-3(p<0.001)、sod- 4(p<0.001)、sod-5(p<0.001)、mtl- 1(p<0.001)的mRNA 水平上调,说明衍生物h 通过上调抗氧化相关基因(sod- 1、sod- 2、sod- 3、sod- 4、sod- 5)和抗菌相关基因(mtl- 1)的表达量发挥抗PA14的作用.
线虫中 DAF-16 在抗应激方面起重要的作用,上游激酶的失活导致DAF-16 从细胞质向细胞核移位[24]. 因此,我们在DAF-16::GFP 标记的突变株线虫TJ356 上分析衍生物h对转录因子 DAF-16 核转位的影响.图6-b 是线虫DAF-16 荧光图像;图6-c 是对不同处理的线虫DAF-16 核转位的量化,对照TJ356 组,DAF-16发生核转运的线虫占比3.85%;PA14感染后发生核转运的线虫占比59.41%,比对照TJ356 组高14.43 倍(p<0.001);经50 µM 衍生物h处理后发生核转运的线虫占比71.43%,发生核转运的线虫比PA14组高20.23%(p<0.01). 说明衍生物h能促进DAF-16 从细胞质到细胞核的易位,在抗感染的过程中通过激活DAF-16 转录因子发挥积极作用.
DAF-16 调节的抗氧化酶SOD-3和CTL-2 通过保护线虫的肠道细胞在感染期间免受ROS 的影响而促进免疫[21],SOD-3 是DAF-16 的下游蛋白之一. 因此,我们使用SOD-3::GFP标记的突变株线虫CF1553 分析衍生物h对 SOD-3 蛋白表达的影响. 图6-d 是不同处理线虫SOD-3 荧光图像,其荧光量化结果如图6-e 所示,PA14 感染激活了线虫体内的天然免疫系统,与对照组 CF1553 相比,PA14/CF1553 组线虫SOD-3 表达量上调2.74%(p<0.01);经50 μM 衍生物h处理后,线虫的天然免疫增强,与PA14/CF1553 组相比,SOD-3 表达量上调1.80%(p<0.01),说明衍生物h可以上调线虫SOD-3 水平.
3 结论
白杨素衍生物h具有抗氧化活性,并且能够延长铜绿假单胞菌 PA14 感染的线虫寿命(p<0.001),说明衍生物h具有抗感染活性. 衍生物h处理显著上调了PA14 感染后宿主ROS 水平和daf-16、sod-1、sod-2、sod-3、sod-4、sod-5、mtl- 1免疫相关基因的mRNA,这表明,ROS 作为信号分子通过促进下游效应因子表达从而激活其他信号通路来增强自身免疫反应,保护宿主抵御PA14 感染. 综上,白杨素衍生物h通过激活转录因子DAF-16 并调控下游免疫相关基因的表达来发挥抗感染作用,机制如图7 所示.
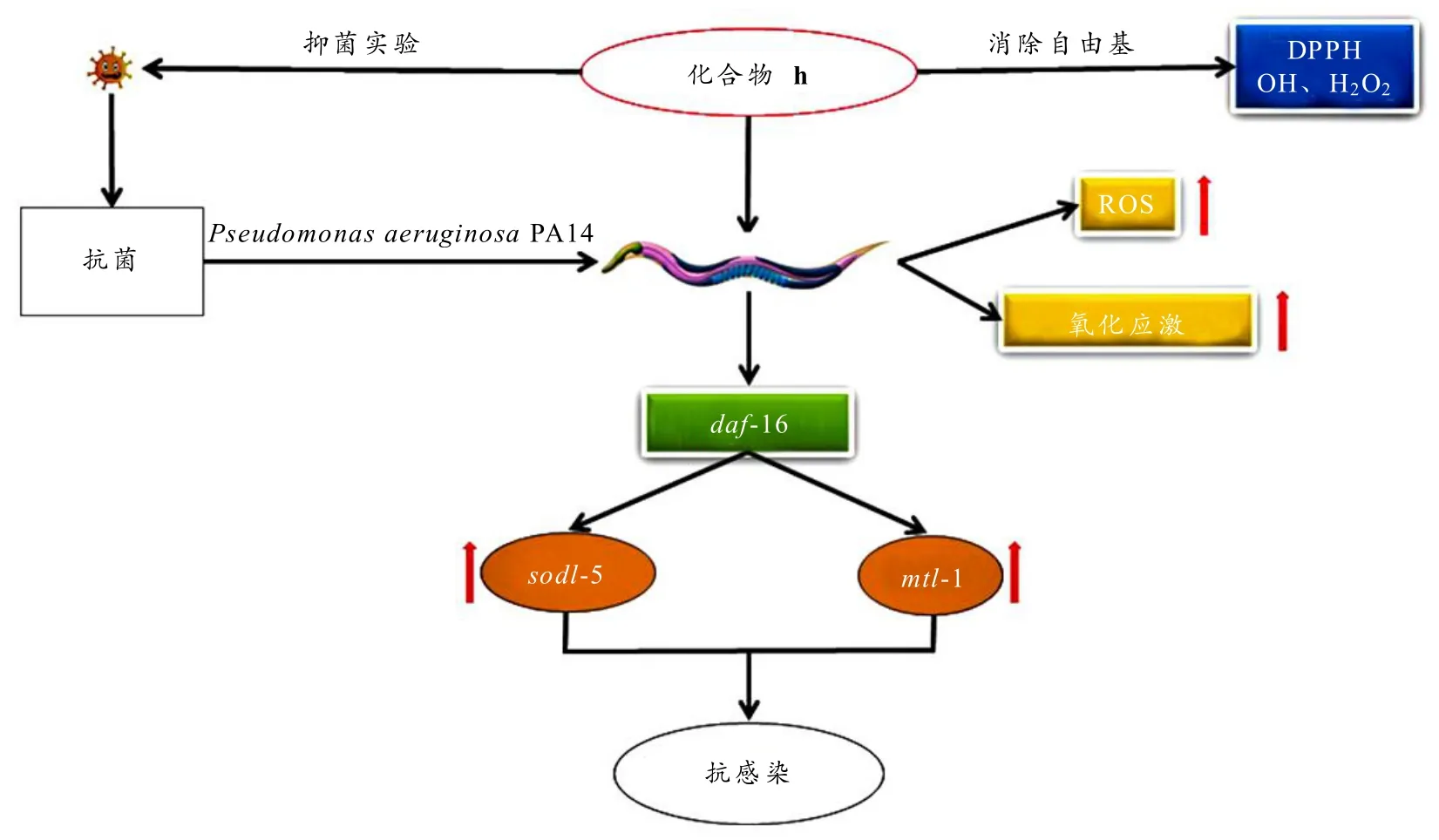
图7 白杨素衍生物h 抗感染作用机制图
