不同模型估计黄河鲤生长性状遗传参数的比较分析
2023-05-18王延晖王冰柯屈长义冯建新常东洲
王延晖,王冰柯,屈长义,冯建新,常东洲,张 芹
( 河南省水产科学研究院,河南 郑州 450044 )
黄河鲤(Cyprinuscarpiohaematopterus),金鳞赤尾、体型梭长、肉质鲜美,是众多养殖的鲤品种中文化底蕴最丰富、养殖历史最悠久的良种,在黄河流域省份具有重要养殖地位。野生黄河鲤因其在生长性能、适应性能方面的优越表现,自20世纪80年代以来,对其人工选择的研究较多,2004年“豫选黄河鲤”通过全国水产原良种审定新品种,此后,针对黄河鲤的遗传改良在国内持续开展[1-3]。以规模化家系为基础的选择育种,是当前黄河鲤遗传改良中主要采用的技术体系,在该体系中,遗传进展主要取决于目标性状的遗传力和选择强度。
遗传力、遗传相关和育种值是3个重要的遗传参数指标。遗传力和遗传相关是预测候选性状的选择响应和育种值的前提,关系到整个育种计划的可行性和良种推广区域的适用性。国内外关于鲤生长性状的遗传参数估计已在多个品种间开展[4-8]。用于评估遗传参数的模型应尽可能准确地反映各种遗传和环境因素对性状的影响,如母本效应和全同胞效应。有学者在草鱼(Ctenopharyngodonidellus)幼鱼[9]、大西洋鲱(Clupeaharengus)[3]、牙鲆(Paralichthysolivaceus)[10]等的相关研究中均发现了母本效应影响的存在,而卢薛等[11-12]则分别证实了全同胞效应对鳜(Sinipercachuatsi)和草鱼生长性状的作用。研究表明,在动物模型中分析不同的效应将影响遗传参数估计的准确性[13-16]。然而,影响黄河鲤及其最近选育群体生长性状遗传参数评估的因素尚不清晰,应对几种可能的模型进行比较,并为提高黄河鲤育种效率提供准确的遗传参数及评估方法。
遗传参数评估依赖于清晰的系谱信息,然而,在大多数鲤的育种实践中,为了便于记录系谱信息,多采用了生长早期家系分开养殖到足以采用物理标记的规格,再进行混合养殖的方式,这导致在后期分析中无法完全剖分环境效应的影响,而且对大量个体进行物理标记成本太高,耗时耗力。基于分子标记的亲子鉴定技术,为控制近交和估计遗传参数提供了有效的技术支持[17],被越来越多地应用到鱼类育种工作中。刘永新等[18]用10个微卫星标记对红鳍东方鲀(Takifugurubripes)混合选育群体进行亲子鉴定,用该系谱估计了150日龄和200日龄体质量和体长的遗传力;Palaiokostas等[19]用SNP标记对镜鲤(C.carpio)混养群体实现了亲子鉴定,并估计了4月龄体质量和体长的遗传力。
笔者以黄河鲤选育群体F3为研究对象,建立全同胞家系,在相同养殖环境中进行混养,以微卫星标记鉴定亲子关系,在分析模型中考虑不同的随机效应,采用3个不同的单性状动物模型估计黄河鲤20月龄生长性状的遗传参数,研究各模型的方差组分间的差异性,筛选出适合黄河鲤20月龄生长性状遗传参数估计的最佳动物模型,同时估计其遗传参数,以期为制定科学高效的黄河鲤育种策略提供基础依据。
1 材料与方法
1.1 试验材料
2015年,自河南省水产科学研究院选育的第2代黄河鲤家系中筛选个体较大、体形良好的个体作为亲本,进行人工催产,雌鱼注射促黄体素释放激素A2100 μg/kg+人绒毛膜促性腺激素 800 IU/kg,雄鱼注射剂量减半。采用半干法授精,建立20个全同胞家系。将每个家系的受精卵分别放入孵化桶中单独培育,经过3周的培育,鱼苗体长达到1.5 cm,不同家系间鱼苗数量差异较大,将全部家系混合后转至3个土池中继续培育。2016年3月,从上述3个池塘中选择5000尾线条流畅、体质量大(前30%)的个体,转移到河南省水产科学研究院试验中心,继续养殖。2017年1月,当混合养殖群体养至20月龄,平均达到上市规格时,进一步筛选700尾线条流畅、体质量大(前30%)的个体留作下一代亲本,从中随机挑选330尾个体进行被动式整合雷达标记(PIT),并测量体长和体质量。同时采集亲本和子代的尾鳍,置于无水乙醇中保存。
1.2 亲子鉴定
使用DNeasy组织试剂盒(北京赛柏生基因技术有限公司)自尾鳍组织中提取基因组DNA。用NanoDrop 2000 UV-Vis分光光度计(Thermo Scientific,美国)检测纯度和浓度,1%琼脂糖凝胶电泳检测完整性后,将DNA样本保存在-70 ℃冰箱中备用。利用亲本DNA自50对微卫星标记[20]中筛选出条带清晰且具多态性的标记,随机选择后代DNA进行荧光引物检测,最终确定8对微卫星标记用于对330个子代和16个亲本的家系鉴定。
用GeneMarker_V2.2分析微卫星基因型,使用Cervus 3.0软件[21]进行亲子鉴定,并计算遗传多样性参数。遗传多样性参数包括等位基因数、观测杂合度、期望杂合度和多态信息含量。
1.3 遗传参数估计
1.3.1 建立混合模型
建立4个单性状动物模型[10,15],分别估计黄河鲤20月龄生长性状的遗传参数:
模型A:yij=μ+Ti+aj+eij
(1)
模型AF:yijk=μ+Ti+aj+fk+eijk
(2)
模型AM:yijk=μ+Ti+aj+ml+eijk
(3)
模型AMF:yijkl=μ+Ti+aj+fk+ml+eijkl
(4)
式中,yij、yijk和yijkl表示第j个个体的生长性状的观测值,μ为总体均值,Ti为池塘的固定效应,aj是第j个个体的生长性状的加性遗传效应,fk为全同胞效应,ml为母本遗传效应,eij、eijk和eijkl为第j个个体的残差效应。
1.3.2 遗传参数评估
通过BLUPF90软件[22],利用平均信息约束极大似然法(AIREML)估计体质量和体长的方差组分。表型方差(σ2P)是模型中所有方差组分的和,以模型AMF为例,表型方差计算公式如下:
(5)

性状遗传力(h2)计算公式如下:
(6)
遗传相关系数(rg)计算公式如下:
(7)
式中,cov(a1,a2)为性状间的遗传协方差,σa1和σa2分别为两性状的加性遗传标准差。
1.3.3 模型比较
用似然比检验(LRT)进行不同模型间的比较,检验统计量(LR)为:
LR=-2(lnL1-lnL2)
(8)

Z值用来检验不同模型估计的遗传力间的差异是否显著,计算公式如下:
(9)
式中,xi和xj分别是不同模型下的遗传力,σi和σj分别是相应的遗传力的标准误。
2 结果与分析
2.1 家系鉴定
用于亲子鉴定的8个位点,其中1个位点(CA2225)因测序质量差最终未被使用。分布在不同连锁群的7个微卫星位点上共检测到64个等位基因,平均等位基因数为9.1个,变化为5~11个。平均观测杂合度为0.795,变化为0.525~0.890。平均期望杂合度为0.801,变化为0.613~0.847。多态性信息含量为0.532~0.828(表1)。7个位点均明显偏离哈迪-温伯格平衡。在95%置信度标准下,模拟鉴定率为100%。各位点的排除概率为19.2%~52.9%,累积排除概率为98.72%。经亲子鉴定,330尾子代中有317尾鉴定到准确的父母本,实际亲子鉴定成功率为96.1%。

表1 7个微卫星标记在黄河鲤中的遗传学统计指标Tab.1 Genetic statistical data of the 7 microsatellite loci in Yellow River common carp C. carpio haematopterus
研究结果显示,317尾子代来自8尾父本、6尾母本,共计15个全同胞家系(图1),第3家系的个体数为111尾,占检测个体的34.91%,有8个家系的个体数超过10尾,剩余家系个体数在1~8尾。各家系、母本和父本对应子代数的卡方检验均存在极显著差异(P<0.01)。

图1 群体的子代分布Fig.1 The offspring distribution of the population
2.2 生长性状描述性统计
获得准确系谱信息个体的20月龄生长性状的描述性统计见表2,体质量变异系数(8.44%)大于体长变异系数(3.09%)。各性状观测值经K-S检验,均符合正态分布(图2a、b)。以箱线图形式给出15个全同胞家系20月龄体质量(图2c)、体长(图2d),2个性状的家系均值变化分别为1051.70~1345.60 g、341~377 mm,体质量和体长在家系间均存在显著差异(P<0.05)。

表2 黄河鲤20月龄生长性状的描述性统计Tab.2 Description statistics of growth traits of 20-month-old Yellow River common carp C. carpio haematopterus

图2 黄河鲤20月龄体长、体质量的分布和家系箱线图Fig.2 Distribute and box plots of body weight, body length in 20-month-old Yellow River common carp C. carpio haematopterus a.体长的频率分布直方图; b.体质量的频率分布直方图; c.体长的家系箱线图; d.体质量的家系箱线图;蓝线表示核密度估计曲线;红线表示正态分布拟合曲线;红线表示正态分布拟合曲线;点为离群值.a.the frequency distribution histogram of body length; b.the frequency distribution histogram of body weight; c.box plots of body length in family; d.box plots of body weight in family; blue line shows kernel density estimation; red line shows normal distribution fitting curve; and spots show outliers.
2.3 生长性状遗传参数估计和模型间的比较
根据标记鉴定结果重构系谱,基于此系谱关系,利用动物模型和平均信息约束极大似然法进行方差组分剖分,对黄河鲤选育群体的体质量和体长的遗传力、遗传和表型相关进行估算(表3)。模型中仅包含加性效应(模型A),估计20月龄体质量和体长的加性方差组分最高,遗传力分别为0.59和0.66。在模型中进一步包含母本效应(模型AM)时,体质量和体长的加性方差组分降低,遗传力分别为0.41和0.16。在模型中考虑全同胞组效应(模型AF)时,加性遗传方差组分为0。将母本效应和全同胞组效应(模型AMF)都纳入模型中,迭代计算不收敛。
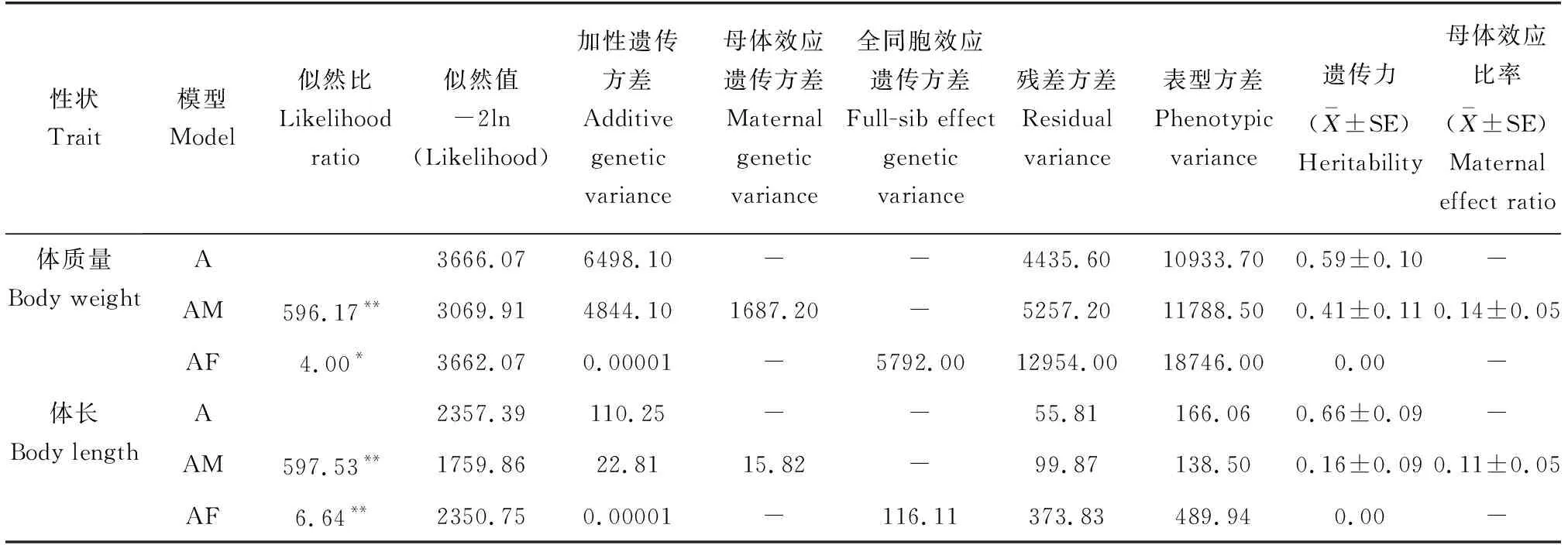
表3 利用动物模型估计的黄河鲤生长性状方差组分Tab.3 Variance components estimation of growth traits in Yellow River common carp C. carpio haematopterus using animal models
不同模型的比较结果见表3。经LRT检验,体质量在模型AM和模型AF与模型A之间的似然比值分别为596.17(P<0.01)和4.00(P<0.05),分别存在极显著差异和显著差异。体长在模型AM和模型AF与模型A之间的似然比值分别为597.53(P<0.01)和6.64(P<0.01),均存在极显著差异。对于体质量和体长,模型AM均为较优模型。
应用两性状动物模型估计性状间的遗传相关系数,体质量和体长的遗传相关系数为0.85,表型相关系数为0.75,表现为高度正相关。综合体质量和体长的遗传变异系数和遗传进展(表4)可知,在一定选择强度下,体质量的选育潜力更大,易获得较大的遗传进展和选择效果。

表4 黄河鲤选育群体生长性状的遗传变异系数与遗传进展Tab.4 Genetic variation coefficients and genetic advances of growth traits in Yellow River common carp C. carpio haematopterus
3 讨 论
3.1 微卫星标记在遗传参数评估中的应用
通过基因分型获得系谱信息进而得以估计遗传力和遗传相关性,被认为是亲子鉴定技术为水产养殖遗传学做出的最重要和富有成果的贡献[17,23]。目前,在水产动物育种中,如草鱼(Ctenopharyngodonidella)[9]、凡纳滨对虾(Litopenaeusvannamei)[24]、红鳍东方鲀(Takifugurubripes)[18]、团头鲂(Megalobramaamblycephala)[25]等多个物种,均有利用微卫星标记辅助家系选育开展遗传参数估计的报道,证实了微卫星标记辅助分子育种的有效性。利用微卫星标记开展遗传参数估计主要有两种策略:一种是利用微卫星对亲本和子代进行基因分型,利用基因型确定父母与子代、子代之间的亲缘关系,即进行亲子鉴定获得系谱,基于该系谱进行遗传参数估计;另一种是直接利用分型结果估计个体间的分子亲缘相关度,进而应用于遗传参数估计。后者通常只对子代进行基因分型。有研究显示,利用分子相关度估计遗传参数更适用于亲缘关系较远的个体,同时标记的数量及遗传多样性也会影响个体间分子相关度估算的准确性。王军[26]利用15个平均等位基因数为39的微卫星位点对中国明对虾(Fenneropenaeuschinensis)67个家系的抗白斑综合征病毒能力进行遗传参数评估,结果显示,利用分子相关度和系谱估计的遗传力之间无显著差异。Lyu等[27]利用20个平均等位基因数为12.4的微卫星位点对大菱鲆79个家系的生长性状进行遗传参数评估,结果显示,利用分子相关度和系谱估计的体质量遗传力分别为0.23和0.33。这进一步证明了不同的标记数目、等位基因数目以及群体亲缘关系不同,分子相关度和系谱对遗传参数评估的影响也不相同。笔者从50对微卫星标记中,筛选出7个均匀分布于黄河鲤7个连锁群上的具有高多态性、高亲本排除概率的标记。对这些标记的遗传学指标进行统计,结果显示,7个微卫星位点的平均等位基因数为9.1个,平均观测杂合度、平均期望杂合度、平均多态信息含量分别为0.795、0.801、0.771,均表明这些标记具有丰富的遗传多样性。但标记数量较少,个体间亲缘关系较近,更适合利用这些标记先开展全/半同胞家系群体的亲缘关系鉴定,再利用此系谱进行混合选育群体体质量和体长的遗传参数估计。本研究结果证明,对于黄河鲤选育群体,采用7个多态性较高的微卫星位点直接对其生长性状进行遗传评估是可行的,并且这些标记可以作为开展类似遗传学研究的备选标记。
3.2 黄河鲤20月龄生长性状遗传参数评估模型
在遗传力估计中,最重要的就是将加性遗传效应从表型方差中准确地剖分出来,而分析模型对于加性遗传效应的剖分具有显著影响,但在实际育种工作中,往往不容易确定影响目标性状的各种遗传因素和环境因素[28],因此,在建立模型时,应尽量考虑可能存在的固定效应和随机效应,通过不同模型比较确定对目标性状具有显著性影响的效应因素。通常认为,母体效应发挥作用的时期多为个体发育早期阶段,而后随着日龄增加母本效应逐渐降低[14,29],如傅建军等[9]基于12对微卫星标记对273尾4月龄草鱼幼鱼的体质量和肌肉成分的遗传参数研究发现,其存在显著的母本效应,在对10月龄和18月龄草鱼的生长性状遗传参数评估中则发现,其没有显著的母本效应和共同环境效应[30]。本研究结果显示,黄河鲤在20月龄的生长性状依然存在显著的母本效应,体质量和体长的母本效应估计值分别为0.14和0.11。分析与以往研究不一致可能与人工繁殖中母本年龄和规格有差异及是否首次产卵等有关。如果在模型中不考虑母本效应,将使遗传力估计值偏高,这与以往研究中,未剖分的母本效应会累加到个体加性遗传效应中的结果[13]一致。
笔者将全同胞家系效应加入分析模型后,体质量和体长的全同胞家系效应值分别为0.30和0.24,但遗传力的估计值均为0。尝试在模型中同时加入母本效应和全同胞家系效应,拟合效果并不理想。虽然Fu等[30]在草鱼相关研究中,对共同环境效应的估计近乎为0,但在本研究中并无证据支持全同胞组效应对生长性状无显著影响,尽管该群体在早期家系单独饲养时间较短。有研究显示,鲤在孵化后的生长早期进行混合饲养可以降低共同环境效应,但是在进入混合饲养阶段前,因家系间成活率差异造成养殖密度不同已经导致生长性状存在明显差异,该经历或已对后期生长差异产生影响[23]。推测本研究中未获得可靠的全同胞家系效应估计值,或与数据结构和子代家系间个体数极不平衡有关,仅有1代系谱,且半同胞家系数量不多,造成全同胞组效应很难被剖分,从而导致遗传力估计值被低估[13,31]。张天时等[15]用4种动物模型对中国明对虾体质量的遗传力进行估计,在不考虑全同胞家系效应时,遗传力为0.76±0.108,但加入全同胞家系效应后,遗传力估计值为0,与本研究结果一致。除了母本效应和全同胞家系效应,显性效应、基因型与环境的互作效应等也应该尽可能被考虑,如有研究显示,鲤8月龄和20月龄体质量主要由显性效应控制[28],星斑川鲽体质量和体长的显性方差分量达到显著水平[32]。
经LRT检验,对于体质量和体长,模型AM均表现出最佳的拟合度,且与其他模型均存在极显著差异,所以对黄河鲤20月龄体质量和体长进行遗传参数估计时,应用包含母本效应的模型AM是比较合理的。总体来说,在遗传方差和育种值估计模型中,不考虑其他非加性遗传效应,会导致加性遗传方差被过高估计,在进行鱼类生长性状遗传评估之前,有必要建立不同的统计遗传模型进行模型选优的过程,会显著提高下一步估计遗传参数和个体育种值的准确度。
3.3 黄河鲤20月龄生长性状的遗传力
黄河鲤在20月龄时基本达到收获规格,因此笔者选择这一时期进行生长性状遗传参数估计,对黄河鲤实际育种工作具有指导意义。国内外对鲤鱼生长性状遗传力评估的研究表明,鲤体质量的遗传力为0~0.71,体长的遗传力为0.01~0.80[33-34],属于中高等水平。本研究获得的体质量和体长的遗传力分别为0~0.59、0~0.66,与以往研究估计范围大体一致[35-37]。遗传力的估计在不同品种或相同品种的不同生长时期、不同的群体遗传背景和结构、不同遗传评估方法以及不同的试验设计中差异很大。Palaiokostas等[19]对黑龙江镜鲤幼鱼(120日龄)体质量的遗传力估计值为0.33;Hu等[38]对松浦镜鲤145日龄、332日龄和392日龄的体质量遗传力估计值分别为0.27、0.28和0.28,属于中等遗传力;镜鲤育成品系17月龄遗传力为0.44[37];Dong等[39]评估了约17月龄的鲤体质量遗传力,为0.17,属于中等遗传力。以上结果进一步表明了生长时期、群体结构和遗传评估方法对遗传力估计有影响。有研究显示,家系混合养殖试验中,尽管有母本效应的存在,但由于家系间没有环境差异,进而使得估计的遗传力往往高于家系单独饲养试验[40]。笔者采用最优模型获得的黄河鲤20月龄体质量和体长的遗传力分别为0.41和0.16,与以往研究相比,黄河鲤20月龄体质量遗传力与镜鲤育成品系17月龄遗传力相当,高于Wang等[41]采用全同胞组内相关法估计的20月龄曲江彩鲤体质量遗传力(0.3)的结果,但体长的遗传力要低于Wang等[41]的结果(0.27),而均低于Wang等[2]对18月龄黄河鲤采用动物模型估计的体质量(0.70)和体长(0.62)的遗传力结果。需要注意的是,上述部分研究在动物模型中只考虑了个体加性遗传效应,无疑提高了遗传力的估计值。
3.4 黄河鲤生长性状的遗传相关
笔者发现,黄河鲤20月龄体质量和体长间存在极显著的高度正相关,与鲤在不同生长时期和其他鱼类中研究结果一致,说明可以通过其中一个性状实现对另一性状的间接选择,综合考虑遗传变异系数和遗传进展,在一定选择强度下,体质量的选育潜力更大,易获得较大的遗传进展和选择效果。目前在黄河鲤的养殖中,20月龄前后是收获期,因此这个时期的体质量和体长是黄河鲤提高生长的主要选择指标。但本研究的局限性也在于只获得了20月龄数据,即收获期的体质量和体长数据,缺少早期生长的表型数据,对于这个种群的早期生长与生长到收获大小之间的相关性尚不清楚。在草鱼生长性状的研究[12]中,10月龄(0.37)和18月龄(0.34)的体质量有较高的遗传相关,认为可以通过评估早期生长率来预测后期的生长潜力,甚至是收获体质量。而另一些研究显示,不同年龄鲤的大小之间的相关性为低到中等[36,38],那么,对幼鱼的预先选择可能是无效的,仅会给体质量带来适度的改善。为了制定合理的育种方案,需要更多的数据来估计不同年龄的遗传力,分析选择阶段和每代的遗传增益。
4 结 论
笔者采用人工授精方式构建黄河鲤全同胞家系,以微卫星分子标记鉴定亲缘关系,通过不同模型的比较分析,确定包含母体效应的模型AM为20月龄黄河鲤生长性状遗传参数估计的最佳动物模型:黄河鲤20月龄体质量和体长受母体遗传效应的影响显著,受全同胞效应的影响难以估计。基于最佳动物模型估计的黄河鲤20月龄体质量和体长的遗传力属于中高水平遗传力,体质量预期遗传进展增加6.72%,以体质量为目标性状可便捷、有效地改良黄河鲤生长性能。
