陆地棉BGLU基因家族成员的全基因组鉴定与表达分析
2023-05-17张曼王志城刘正文王国宁王省芬张艳
张曼, 王志城, 刘正文, 王国宁, 王省芬, 张艳
(河北农业大学农学院,华北作物改良与调控国家重点实验室,河北省作物种质资源重点实验室,河北 保定 071000)
棉花是重要的经济作物,对中国经济发展有着重要的作用。继二倍体雷蒙德氏棉(Gossypium raimondii)[1]和亚洲棉(Gossypium arboreum)[2]测序完成,生产上应用最广泛的陆地棉(Gossypium hirsutum)和海岛棉(Gossypium barabdense)基因组序列也已绘制完成[3-4],标志着棉花研究进入了后基因组时代,为开展基因家族的全基因组鉴定与生物信息学分析提供了基础信息。
β-葡萄糖苷酶(β-glucosidase)EC3.2.1.21,又称β-D-葡萄糖糖苷水解酶,属于纤维素酶类,是纤维素分解酶系中的重要组成成分。β-葡萄糖苷酶能够从含糖化合物中催化水解末端的非还原性β-D-葡萄糖苷键,同时释放β-D-葡萄糖和相应的配基[5]。在高等植物中,β-葡萄糖苷酶主要包括糖基水解酶家族1 (glycoside hydrolase family 1,GH1)[6],其广泛参与防御反应[7]、细胞壁木质化[8]、植物激素信号激活[9-10]和次生代谢[11]等重要生理过程。通过对β-葡萄糖苷酶家族的系统发育进行分析,发现拟南芥AtBGLU45、AtBGLU46和AtBGLU47与松柏(Pinus contorta conifin)β-葡萄糖苷酶基因聚为一类,它们可能通过水解单酚苷参与木质化过程[12]。抑制水稻Os3BGLU6的表达导致植株矮化、叶片中ABA 含量降低、干旱敏感性增加、光合速率降低、细胞间CO2浓度升高,影响植株的耐旱性[13]。比较分析5 种蔷薇科植物的β-葡萄糖苷酶家族全基因组发现,梨(Pyrus bretschneideri)果实中BGLU基因与木质化和石细胞发育有关[14]。水稻(Orazy sativa)中的OsGH3基因可以调控生长激素、茉莉酸和水杨酸信号通路之间的串联,从而影响水稻对生物和非生物胁迫的耐受性[15]。β-葡萄糖苷酶家族除了参与植物的细胞壁代谢及激素的激活外,还在植物逆境胁迫的防御反应中起重要作用。
生物信息学在全基因组分析中发挥着越来越重要的作用,棉花的多个重要基因家族已经通过全基因组分析取得了重要成果[16-17]。目前,从基因组水平解析棉花β-葡萄糖苷酶基因(BGLU)家族的相关研究还未见报道。本研究从全基因组水平鉴定了陆地棉中的BGLU基因家族成员,对各成员的基因结构、染色体定位、系统进化关系、启动子调控元件等进行了系统分析,并结合黄萎病菌处理后的抗病‘农大601'的转录组数据对参与黄萎病抗性反应的GhBGLU基因进行了筛选和相关qPCR 验证,为后续深入研究BGLU基因在陆地棉中的功能奠定基础。
1 材料与方法
1.1 GhBGLU基因家族的鉴定
根据GhBGLU基因序列对最新版本的高质量陆地棉基因组(https://cottonfgd.org/)进行BLAST搜索,确定候选基因,进一步利用GhBGLU家族特有保守结构域(PF00232)通过Pfam 数据库[18](http://pfam.xfam.org/)和NCBI CD-Search 数据库(https://www.ncbi.nlm.nih.gov/cdd/)进行验证,剔除冗余序列后,最终得到陆地棉BGLU家族成员的序列。
1.2 GhBGLU家族多重序列比对及进化树构建
通过Clustal W 程序对陆地棉BGLU 氨基酸序列进行多序列比对[19],利用MEGA 7.0 软件的邻接法(Neighbor-Joining, NJ)构建系统进化树,Bootstrap重复测试(replications)设置为1 000。
1.3 GhBGLU家族基因生物信息学分析
利用在线软件MEME (http://meme.sdsc.edu/meme/)分析保守结构域,结合GhBGLU家族基因外显子-内含子结构,利用TBtools 进行可视化。根据基因组结构注释文件的基因信息可知各BGLU家族成员在染色体上的位置,利用MapChart 软件进行基因染色体定位。使用MCScanX 软件对陆地棉GhBGLU家族成员的共线性进行分析[20],最后使用Circos 软件对结果进行可视化展示[21]。利用TBtools 软件对共线性基因进行Ka/Ks计算,并评估其进化选择方向[22]。从陆地棉基因组数据库中选取BGLU家族中起始密码子ATG 上游2 000 bp 的序列,利用PlantCARE 数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析其可能存在的顺式作用元件,筛选后利用TBtools软件作图。
1.4 GhBGLU 基因家族在黄萎病胁迫下的表达量热图绘制
利用本实验室大丽轮枝菌胁迫的棉花转录组数据库[23]进行BGLU家族基因表达模式分析,使用TBtools 软件绘制BGLU基因家族各成员表达量热图。
1.5 GhBGLU基因的qRT-PCR
以本实验室保存的陆地棉抗病品种‘农大601'(ND601)为材料,在黄萎病菌胁迫6、12、24、36、48 h后取根部样品,提取RNA并进行Real-time PCR。以UBQ作为内参,采用 2-△△CT法[24]计算基因的相对表达量,每个基因的表达反应有3个生物重复和3个技术重复。本文所用引物详见表1。

表1 本研究中所用qRT-PCR引物序列Table 1 Primer sequences for qRT-PCR in this study
2 结果与分析
2.1 GhBGLU基因家族鉴定与理化性质分析
在陆地棉‘TM-1'基因组中共鉴定到53 个编码GhBGLU基因的家族成员,根据它们在染色体上的位置分别命名为GhBGLU01~GhBGLU53(表2)。GhBGLU基因编码的氨基酸范围在121~546 aa 之间,蛋白质分子量分布在13.9~66.3 kD 之间;等电点变化范围则从4.694~9.728。利用不同的软件对GhBGLU基因家族成员进行亚细胞定位预测,分析结果显示,GhBGLU家族不同成员定位于细胞质膜、液泡、胞质、胞外和溶酶体中。信号肽预测发现,53 个基因家族成员在N 末端均存在信号肽,且有16 个成员含有跨膜域,推测它们可能为分泌蛋白。
2.2 GhBGLU家族基因的系统发育分析
基于陆地棉和拟南芥蛋白序列构建系统发育树,GhBGLU基因被分为6 个亚家族(A~F)(图1),每个亚族均同时包含GhBGLU和AtBGLU基因,表明这些成员功能保守。AT1G52400、AT2G44490、AT5G57220、AT1G51490、AT3G09260等基因参与了拟南芥对病菌及虫害的防御反应[25]。另外,拟南芥AT2G44450可催化黄酮类物质的合成反应[26];AT3G09260、AT3G15950参与了茉莉酸调控通路[27];AT5G52310、AT5G60360通过调节脱落酸水解活性在拟南芥渗透胁迫响应中起重要作用[28]。根据同源进化关系,推测棉花中与拟南芥同源的这些基因可能参与了棉花对生物、非生物的胁迫响应。
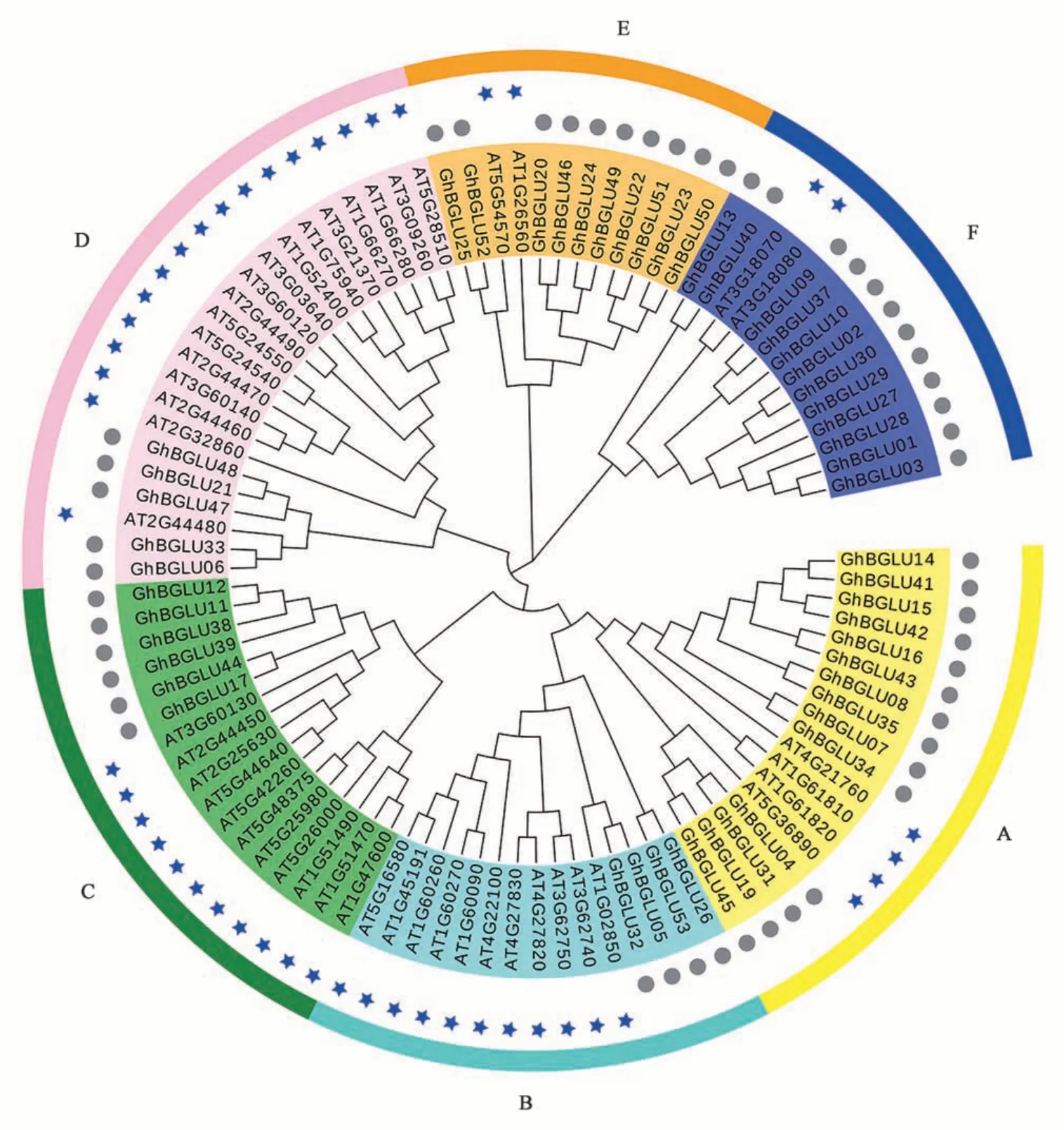
图1 陆地棉和拟南芥BGLU基因家族进化分析Fig. 1 Phylogenetic analysis of BGLU proteins in G. hirsutum and Arabidopsis
2.3 GhBGLU 家族基因的染色体定位、基因结构和保守域分析
根据基因位置信息,发现陆地棉中53 个GhBGLU基因不均匀地分布在除A 亚组第3、4 号染色体和D亚组第2、4号染色之外的22条染色体上,且主要分布在染色体两端(图2)。此外,GhBGLU在A、D亚基因组间无明显偏好性。
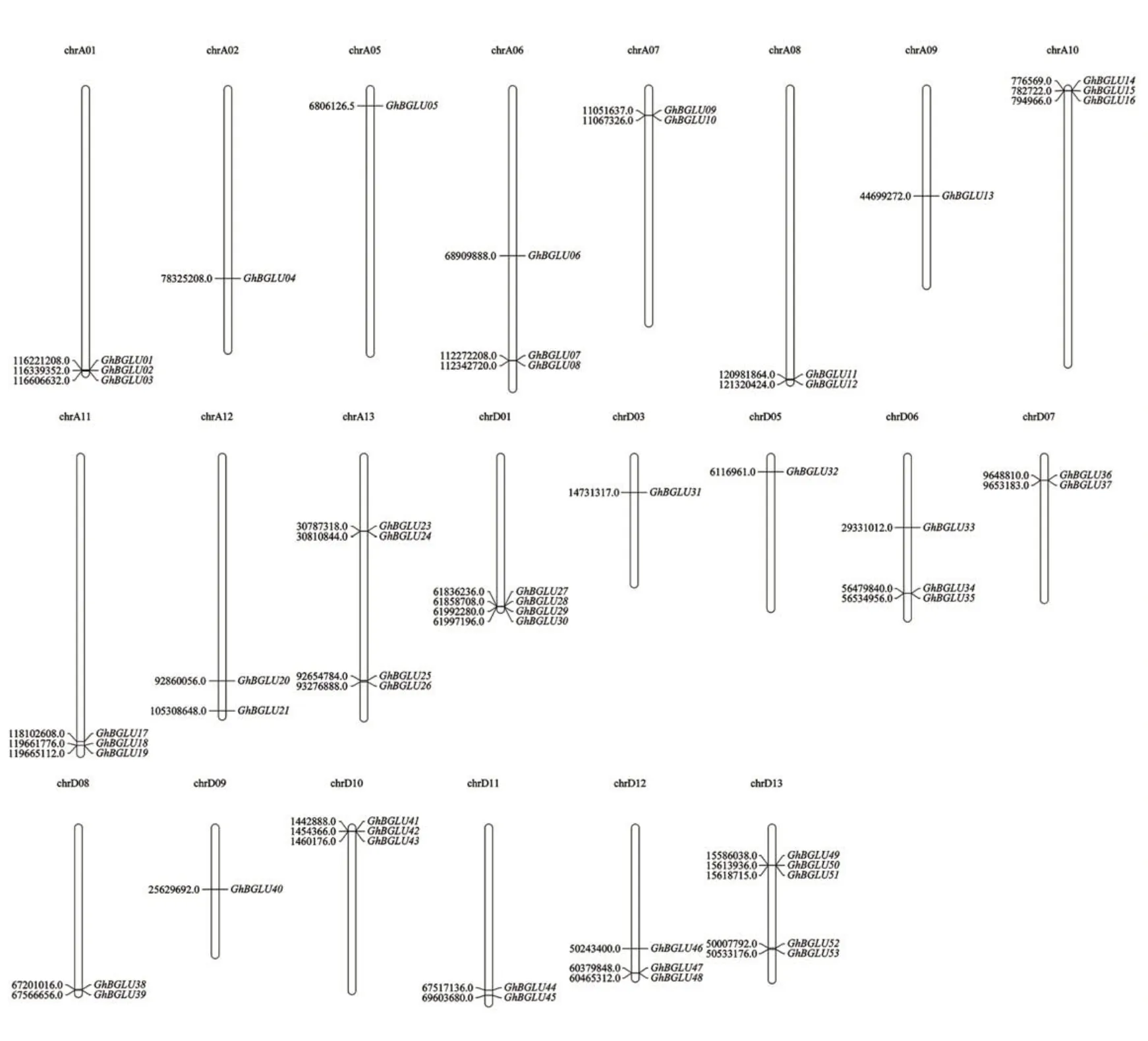
图2 陆地棉BGLU基因家族成员的染色体分布Fig. 2 Chromosomal location of the GhBGLU genes in G. hirsutum
陆地棉GhBGLU基因编码的蛋白含有8 种motif,分别命名为motif 1~motif 8(图3)。这些motif 在GhBGLU家族大部分成员中均有出现,为该家族的保守基序,推测其在该家族中行使基本功能。GhBGLU家族外显子个数变异较大,GhBGLU17、GhBGLU38、GhBGLU44仅含有1 个外显子,而GhBGLU4基因含有的外显子高达14 个。大豆[29]和水稻[30]中外显子数目基本在11~13 左右,个别基因出现了8~9个。

图3 陆地棉GhBGLU基因家族保守基序Fig. 3 Conserved motifs of GhBGLU gene family in G. hirsutum
2.4 GhBGLU 家族基因共线性及顺式作用元件分析
为研究GhBGLU家族基因进化的关系,使用MCScanX 软件进行共线性分析,共发现21对共线性基因对。A、D 亚组中均发现2 对片段复制基因,A 亚组和D 亚组之间有17 对同源基因呈绝对共线性(图4)。通过计算上述基因对的非同义突变率与同义突变率的比值(Ka/Ks)发现,同源基因对的Ka/Ks均小于1,表明这些基因在进化过程中受到了纯化选择。

图4 陆地棉GhBGLU共线性分析Fig.4 Collinearity analysis of GhBGLU in G. hirsutum
通过对GhBGLU基因上游2 000 bp 序列的启动子区顺式作用元件进行分析发现,存在与激素信号相关的脱落酸(abscisic acid, ABA)响应元件、赤霉素(gibberellic acid, GA)响应元件、auxin响应元件、水杨酸(salicyclic acid, SA)应答元件、茉莉酸甲酶(methyl jasmonate, MeJA)应答元件以及与环境胁迫相关的低温应答元件、干旱应答元件和防御胁迫应答元件(图5),其中,MeJA 反应元件最丰富(121 个)。上述应答元件的富集表明,棉花BGLU基因可能参与了植物对各种病原菌及环境胁迫的响应。

图5 陆地棉GhBGLU启动子顺式作用元件分析Fig.5 Analysis of cis-acting elements of GhBGLU promoter in G. hirsutum
2.5 黄萎病菌胁迫下GhBGLU基因表达分析
利用黄萎病菌胁迫下抗病陆地棉农大601 的转录组数据库[23],依据GhBGLU基因家族成员接菌后不同时间点的RPKM 值构建基因表达热图,共筛选出33个GhBGLU基因在大丽轮枝菌胁迫下差异表达(图6),表现为2 种模式:一种是基因在病菌胁迫处理后表达量出现上升趋势,包括GhBGLU5、GhBGLU34等,表明这些基因在棉花抗黄萎病反应中起正调控作用;另一种是在黄萎病菌处理后基因表达量呈现出下降趋势,包括GhBGLU22、GhBGLU52等,表明这些基因在棉花抗黄萎病反应中起负调控作用。
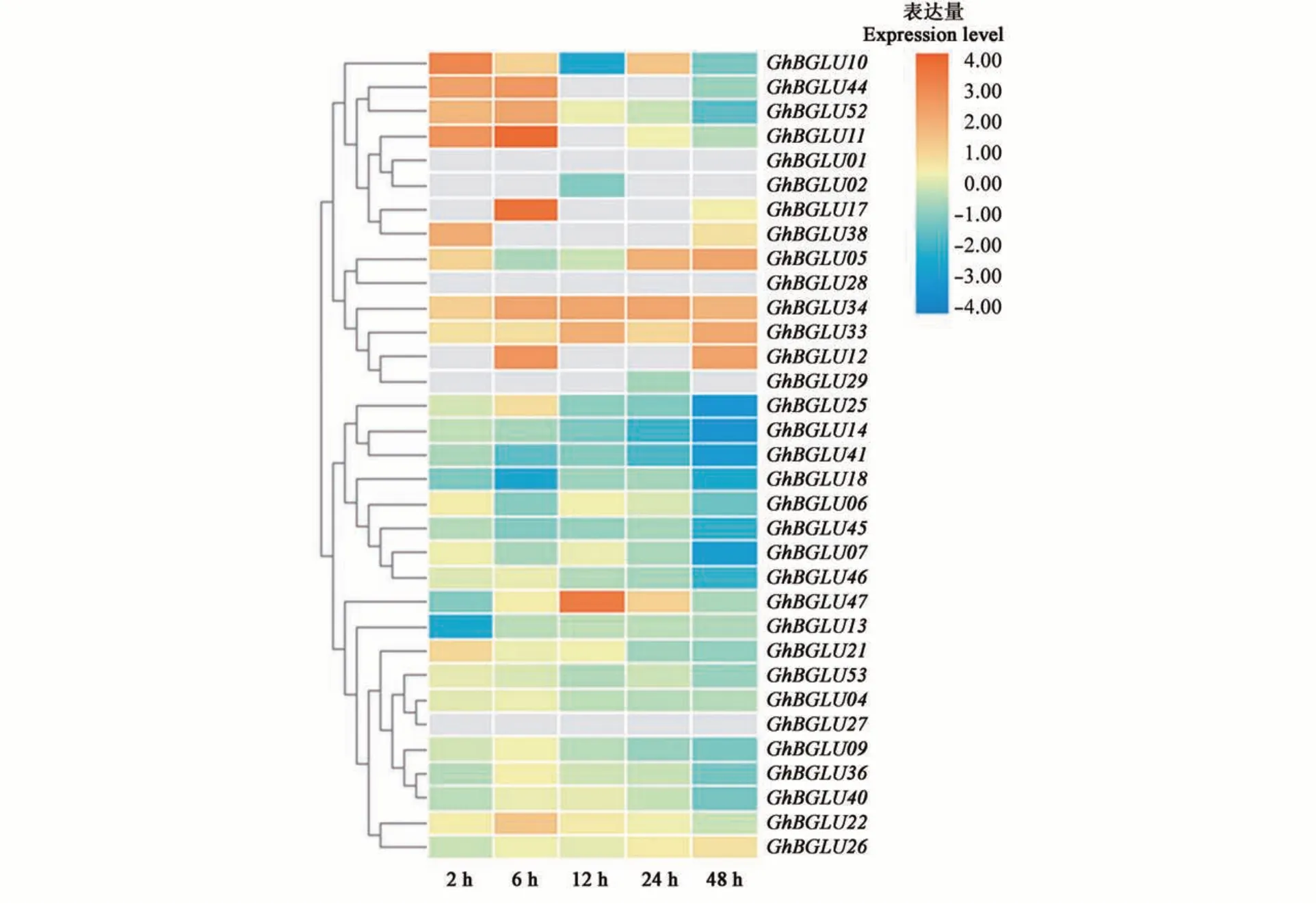
图6 陆地棉大丽轮枝菌胁迫下GhBGLU基因表达分析Fig. 6 GhBGLU genes expression under Verticillium dahliae stress
2.6 不同抗性棉花品种中GhBGLU基因的表达分析
为明确上述响应大丽轮枝菌胁迫的GhBGLU基因是否参与棉花抗病反应,从表达差异明显的基因中挑选了9 个成员,进一步在不同抗感棉花品种中检测这9 个基因的表达水平(图7)。荧光定 量PCR 结 果 显 示,GhBGLU5、GhBGLU14、GhBGLU18、GhBGLU26、GhBGLU34、GhBGLU33在受到黄萎病菌胁迫后,表达量显著提高,且在抗病品种‘ND601'中的表达量显著高于感病品种‘CCRI8',推测上述基因正向调控棉花黄萎病的抗性。GhBGLU22、GhBGLU40、GhBGLU52表达量在‘ND601'中显著下调,但在‘CCRI8'中的表达没有明显规律,需要进一步研究其参与黄萎病菌抗性的作用方式。此外,在接菌后12 h 的抗病品种‘澳SiV2'和感病品种‘湘棉18'的转录组数据中也发现,上述9个基因中的7个基因在‘澳SiV2'的转录水平高于‘湘棉18'中,与荧光定量结果较吻合。结合抗感品荧光定量和转录组测序结果,推测GhBGLU5、GhBGLU14、GhBGLU18、GhBGLU26、GhBGLU34在抗黄萎病反应中起正调控作用。
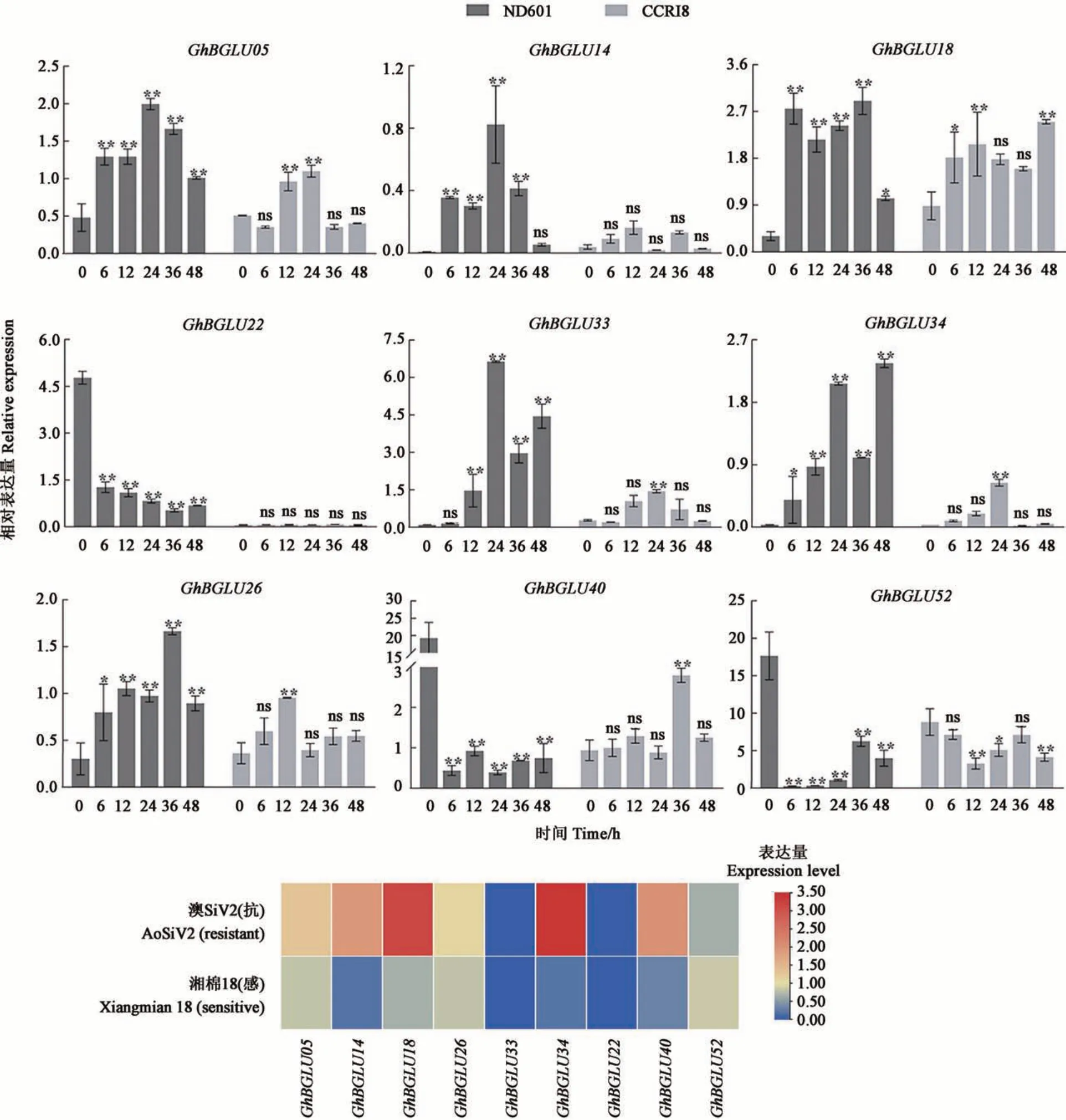
图7 黄萎病菌胁迫下9个GhBGLU基因的表达分析Fig. 7 Analysis of the expression of 9 GhBGLU genes under the stress of V. dahliae
3 讨论
β-葡萄糖糖苷酶参与植物体内的各类生理过程,主要在植物次生代谢及逆境胁迫中发挥重要作用[5]。本研究利用生物信息学手段从陆地棉中共鉴定出53 个BGLU家族基因,它们不均匀地分布在22 条染色体上,且主要分布在染色体两端,包含8 个典型保守基序。大多数BGLU家族基因都是成对出现的,同源性高,推测这些基因在功能上有一定的相似性。通过系统发育分析,53 个BGLU家族基因被分为6 个亚组,且每个亚组均有拟南芥出现,表明棉花BGLU基因与拟南芥BGLU基因具有同源性,但是拟南芥成员分布不均,导致棉花在某些蛋白序列表现出进化过程中的差异,可能和物种的自然选择有关系。共线性分析发现了21 对基因对,同源基因对的Ka/Ks均小于1,表明这些基因在进化过程中受到了纯化选择。亚细胞定位结果显示这些成员均定位在胞外,N 端均存在信号肽,表明该家族基因功能保守。
通过对几种已测序的模式植物进行研究,如拟南芥[31]、苜蓿[32]、玉米[33]、水稻[34],发现BGLU基因均响应生物和非生物胁迫,与激活植物激素和防御化合物相关。植物BGLU基因家族对病菌胁迫有着不同程度的响应,其应答机制也不尽相同,拟南芥BGLU基因家族与抗病相关的通路均有关联,如木质素[35]、水杨酸[36]、黄酮醇[37]、乙烯[38]。AtBGLU6(AT1g60270)过表达可以调控异黄酮的积累,提高植物对病菌胁迫防御。OsbspP(BGLU_RS28885)的缺失减弱了水稻稻瘟病对寄主的侵染力。研究表明,GhBGLU与β-1,3-葡聚糖酶同属于GH 超家族,定位于液泡和胞外的β-1,3-葡聚糖酶可以通过降解真菌细胞壁相关成分对病原菌进行防御[39-40],亚细胞定位预测将GhBGLU家族中的多个成员定位在液泡或胞外,推测其可能与β-1,3-葡聚糖酶发挥类似的功能。
本研究分析了陆地棉BGLU基因家族对黄萎病胁迫的响应,发现大多数GhBGLU基因受黄萎病菌胁迫后显著表达,表明棉花可通过调控部分BGLU基因来提高植物对病原菌抗性。对GhBGLU基因家族进行顺式作用元件分析,发现多个与抗病相关的应答元件,如多种激素应答元件、防御相关应答元件和非生物胁迫应答元件等。植物激素水杨酸、茉莉酸被认为是植物抗病防卫反应的内源及中间信号分子,而MeJA 作为数目最多的应答元件可以直接对病原菌起到抑制作用,并作为抗病防卫反应的内源及中间信号分子发挥作用[41]。因此,推测GhBGLU基因启动子序列中植物激素响应元件可能调控棉花抗病过程。根据qRT-PCR 结果,棉花中有6 个GhBGLU基因受黄萎病菌诱导后表达量显著上调,3 个GhBGLU基因受黄萎病菌诱导后表达量显著下调,这些BGLU基因可能作为介导棉花对黄萎病抗性的调控因子,在响应黄萎病胁迫中具有重要作用,具体的抗病机制还需要进一步研究。
