基于红外相机技术的野生勺鸡行为谱及PAE编码系统
2023-05-16周一凡赵铁建冯小梅孙国明王玉申沈素侠卢宪旺关洪武张卫兵赵大鹏
周一凡,赵铁建,冯小梅,孙国明,王玉申,沈素侠,卢宪旺,关洪武,张卫兵,赵大鹏
(1.天津师范大学生命科学学院,天津,300387;2.天津八仙山国家级自然保护区管理服务中心,天津,301909;3.天津盘山风景名胜区管理局,天津,301900)
行为多样性作为生物多样性的有机组成成分,是物种对环境适应能力的体现[1],因此开展行为多样性研究对于物种的科学保护具有积极意义[2]。栖息地资源特点[3−4]、季节[5]、性别[6]、年龄[7−8]和人为干扰[9−10]等均会对物种行为表达产生一定影响,如灰鹤(Grus grus)在食物资源较难获取时会增加取食时间[11];朱鹮(Nipponia nippon)在冬季的休息和理羽行为高于夏秋季[12];黑颈鹤(Grus nigricollis)在产卵前期雌性个体较雄性个体表现出更多的取食行为[13];成年白鹤(G.leucogeranus)用于警戒的时间显著高于幼体,而取食行为的时间分配则相反[14];人为干扰会使野生小天鹅(Cygnus columbianus)减少能量摄入,并增大警戒行为的能耗[15]。物种行为学研究应充分考虑生活环境,而以“姿势-动作-环境”(posture-act-environment,PAE)编码系统对物种行为展开研究有助于全面了解物种行为的功能、目的及行为之间的关系[16]。完整行为谱的建立对物种行为学方面的定量研究具有重要意义[17],能为物种更深层面的行为研究提供坚实基础。
勺鸡(Pucrasia macrolopha)为国家二级重点保护野生动物,在IUCN物种红色名录中评估等级为无危(LC)[18],野生种群主要分布于中国、印度、巴基斯坦和阿富汗等多个国家,国内主要分布于天津、西藏、陕西和辽宁等地[19]。勺鸡属典型的森林鸟类,偷猎、栖息地丧失和生境破碎化等生存威胁[20−21]使其种群数量不断下降。目前关于勺鸡行为分类方面的专业研究仅局限于笼养个体[22−23],而物种在笼养条件下难以充分体现自然行为特征[24−25]。野生勺鸡性情机警,常隐于密林,传统的样线法等人工调查可能对野生种群行为产生影响[26],而无损伤、低干扰的红外相机技术对于开展野生环境下物种行为学的研究具有独特的优势[27]。本研究首次应用红外相机技术建立野生勺鸡行为谱和PAE 编码系统,为相关物种的行为生态学研究和综合保护管理提供科学参考依据。
1 研究区概况
研究区位于燕山山脉、天津市蓟州区的天津八仙山国家级自然保护区(40°10′47″—40°13′56″ N,117°31′55″—117°33′53″ E)(以下简称为八仙山)和天津盘山风景名胜区(40°4′45″—40°6′25″ N,117°14′40″—117°17′20″ E)(以下简称为盘山),监测起止时间分别为2017 年6 月—2018 年10 月和2019 年7 月—2021 年4 月。两地的总面积分别为10.49、110.90 km2,植被类型丰富,包括针叶林、阔叶林和 针阔混交林等,属于暖温带季风型气候,四季分 明,夏季高温多雨,冬季寒冷干燥。八仙山年平均 降水量为968.5 mm,主峰聚仙峰海拔1 046.8 m;盘山年平均降水量为800 mm,主峰挂月峰海拔864.4 m[28−29]。根据研究区域气候特点,将3—5月划为春季,6—8 月为夏季,9—11 月为秋季,12—翌年2月为冬季[30]。
2 研究方法
2.1 相机布设
基于地理信息系统将八仙山和盘山分别划分为500 m×500 m 和1 000 m×1 000 m 的网格,在每个网格内选择具有勺鸡活动痕迹(兽道、水源地等)的位置布设红外相机,红外相机主要固定在距地面约0.5 m 处的树干或岩石等固定物上,基础设置为2或3 次连拍、拍摄间隔1 s 和24 h 工作,拍摄模式为照片或照片加视频。红外相机最初布设时,分别记录每台放置的日期、地点、编号、经纬度、海拔和生境等信息。两地红外相机的布设数量分别为30、40 台。监测期间,每3~4 个月更换1 次相机储存卡和电池。
2.2 数据整理与统计分析
从收集到的红外影像数据中,分别对勺鸡的年龄、性别及个体行为进行分类整理。将勺鸡的年龄性别组划分为成年雄性、成年雌性及幼体,其中成年雄性的判定依据为头部金属暗绿色,具黑色长冠羽,颈部具明显白斑,白斑后及背上部黄棕色,喉黑色,上体羽黑白相间呈披针形,下体羽从黑喉至腹深栗色;成年雌性的判定依据为体型大小和成年雄性接近,头部皮黄色,具短冠羽,颈部、喉部均具白斑,眼周具棕白色眉纹,黑点紧密分布在眉纹之上,上体羽黄褐色并杂有黑色斑,下体羽从白喉至腹部多为淡黄栗色;幼体的判定依据为体型小,身披绒毛或与成年雌性羽毛颜色相近,难以区分雌雄[31]。
行为辨识方面,参考笼养勺鸡和褐马鸡(Crosso⁃ptilon mantchuricum)的相关行为分类体系和行为特征[22−23,32]来定义野生勺鸡的行为、姿势及动作。对拍摄到的所有野生勺鸡的照片和视频数据进行行为辨识,对姿势、动作和环境分别进行P码、A码和E码的编写,并依据行为产生的环境,以及对应产生的姿势和动作建立PAE 编码系统,依据行为功能将野生勺鸡的行为划分为停歇、警戒、整理、取食、运动、家庭和社会7类。
活动特征方面,为降低物种与照片数目的自相关性,将30 min 内拍摄到的第1 张照片作为独立探测首张,若30 min内拍摄到多只野生勺鸡,则取其中数目最多的照片进行分析[33]。使用Pi=Ti/Bi×100%计算勺鸡在不同时间区段内的行为频次比例,其中i代表月份、季节,Ti表示该时间段内物种的行为频次,Bi表示该时间段物种行为的总次数。在数据分析方面,对不同月份、不同季节对比分析时,首先使用Kolmogorov-SmirnovZ-test 检验数据的正态性,若不符合正态分布,则使用Mann-WhitneyU检验或卡方检验,若符合正态分布,则使用独立样本的t检验[34]。
行为多样性方面,参考阿拉伯狒狒(Papio hama⁃dryas)[35]、中华鬣羚(Capricornis sumatraensis)[36]等物种行为多样性的研究方法,分别计算样本的绝对行为多样性指数H(absolute diversity index)、最大行为多样性指数Hmax(maximum diversity index)、相对多样性指数r(relative diversity index)和校正多样性指数r-variable(regulate diversity index),各计算公式如下:
式中:fi是野生勺鸡第i种行为的发生频次。
式中:N是野生勺鸡行为谱中的行为类别数。
式中:n是野生勺鸡特定行为类别的所有行为元素数。
在数据分析方面,考虑到具体数据量及各行为多样性指数的特性,仅关注H、r两项行为多样性指数的季节差异与性别组差异,其中在季节性差异方面,应用Wilcoxon 检验仅关注整体数据上H、r两项行为多样性指数两两季节间的差异,应用Friedman检验H、r两项行为多样性指数全年季节间的差异;而在性别组差异方面,应用Mann-WhitneyU检验仅关注H、r两项行为多样性指数之间的性别差异。应用SPSS 26.0软件完成相关数据分析,显著性差异水平定义为p≤0.05。
3 结果
3.1 姿势编码
共监测到勺鸡有效照片数据556张(八仙山,430张;盘山,126 张),有效视频58 段(八仙山,51 段;盘山,7 段)包含有效独立探测首张113 张(八仙山,77张;盘山,36 张)。共辨识并记录到“站”“立”“蹲”“卧”“走”“跑”“飞”和“跳”8种姿势,前4种是静止姿势,后4种为运动姿势(表1)。

表1 勺鸡的姿势编码及定义Tab.1 The code and description for each posture in Pucrasia macrolopha
3.2 动作编码
根据照片和视频数据中野生勺鸡发生动作的部位不同,将动作划分为头颈部、躯干及尾部、喙部、翅部和腿部5 个部分,共辨识记录到野生勺鸡31种动作,并对其编码(表2)。

表2 勺鸡的动作编码Tab.2 Action codes of Pucrasia macrolopha
3.3 环境编码
依据野生勺鸡行为发生的非生物环境和生物环境、年龄以及集群模式等,将环境划分为灌木、石头、水源、落叶阔叶林、针阔混交林、雌性与幼体、成体群和单一个体8种类型,并对其编码(表3)。

表3 勺鸡的环境编码Tab.3 Environment codes of Pucrasia macrolopha
3.4 勺鸡PAE编码系统
共监测到野生勺鸡21种行为,分别在停歇、警戒、整理、取食、运动、家庭和社会7 大类型之下,综合野生勺鸡行为发生的姿势编码、动作编码和环境编码,再加上季节PAE编码,并与已有研究中的笼养和野生勺鸡种群进行对比,建立勺鸡PAE 编码系统(表4)。停歇行为,在环境中维持同一姿势,个体呈放松状态;警戒行为,在环境中持续徘徊,对周围环境保持警惕;整理行为,维持自身整洁或排出代谢废物;取食行为,从自然环境中获取食物资源;运动行为,通过调整姿势改变自身位置;家庭行为,雌雄集群,雌幼集群下产生的行为;社会行为,不同个体间的相互交流。
3.5 活动特征
基于整体数据,野生勺鸡各行为类型的频次在月份间存在显著差异(χ2=26.762,dƒ=11,p=0.005);频次比例位居前3 位的行为类型为运动、警戒和取食,每种行为的行为频次比例均不存在季节间显著差异(表5,图1)。
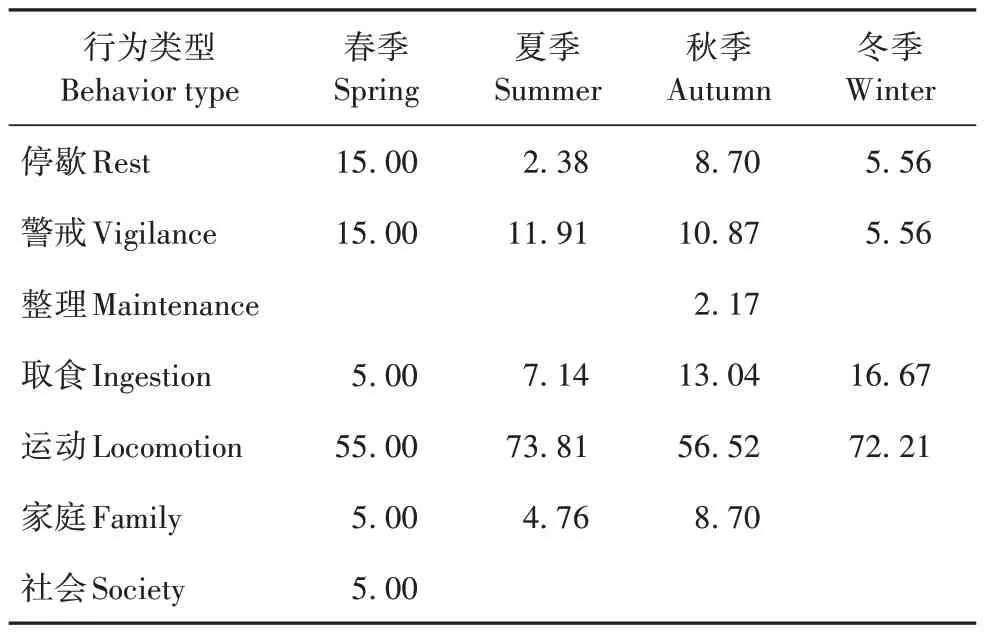
表5 不同季节勺鸡各类行为的频次比例Tab.5 Frequency percentages of each behavior at different seasons in Pucrasia macrolopha %

图1 不同月份勺鸡各类行为的频次比例Fig.1 Frequency percentages of each behavior at different months in Pu⁃crasia macrolopha
3.6 行为多样性
共辨识出野生勺鸡21种行为元素(成年雄性18种,成年雌性12种,幼体4种),Hmax为4.39。基于整体数据发现:(1)成年雄性和成年雌性的Hvariable、H、r和r-variable 均高于幼体(表6);(2)成年雄性的Hvariable高于成年雌性,而成年雌性的H、r和r-variable均高于成年雄性(表6);(3)H、r季节间不存在显著差异(H:χ2=3.000,df=3,p=0.392;r:χ2=4.071,df=3,p=0.254)。

表6 不同年龄性别组勺鸡的行为多样性Tab.6 Behavioral diversity indexes at different age and sex groups of Pucrasia macrolopha
在性别组比较方面,发现:(1)成年雄性和成年雌性秋季的H和r均高于夏季;(2)成年雄性春季和秋季的H和r均高于成年雌性;(3)行为多样性指数H和r均不存在显著性别组差异(H:Z=−0.857,p=0.391;r:Z=−0.938,p=0.348)(图2)。
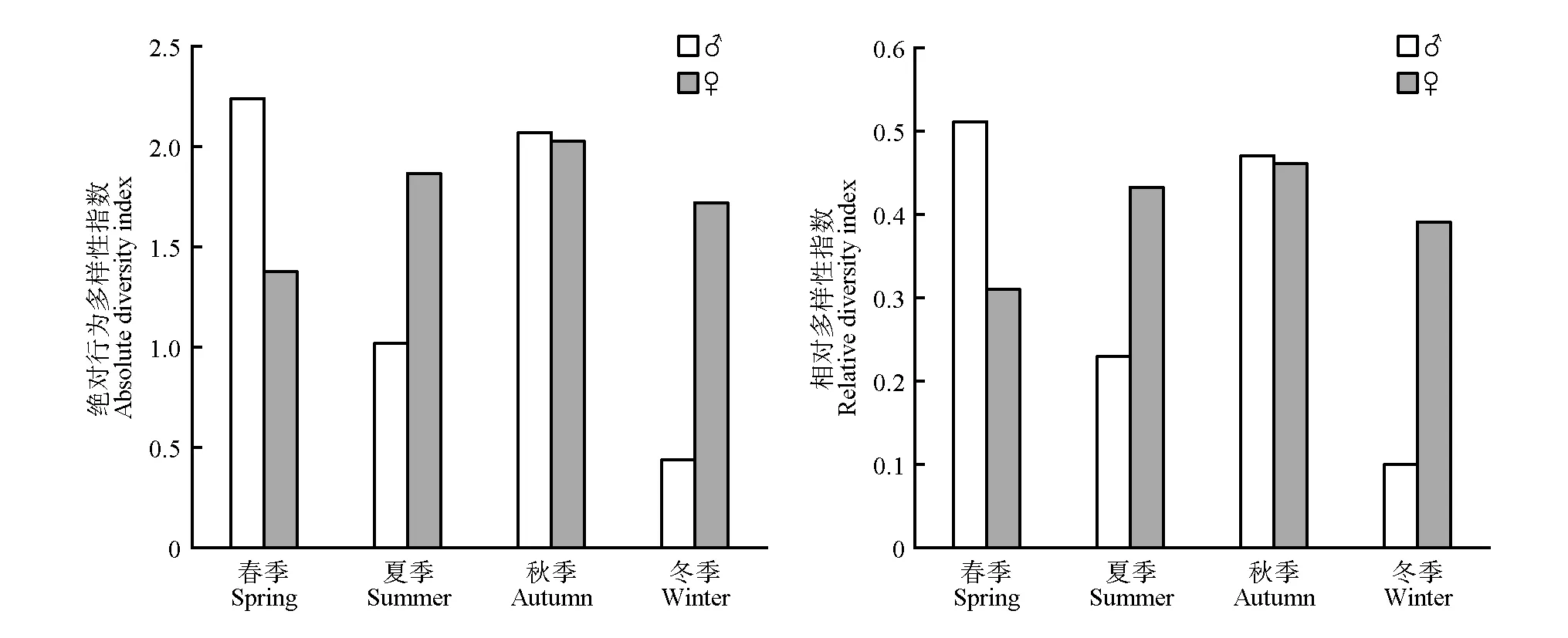
图2 各季节成年勺鸡的H和rFig.2 H and r of Pucrasia macrolopha at different seasons
4 讨论
物种的行为受环境因素和自身因素等多方面影响[45−51]。笼养物种由于活动范围有限,没有天敌,食物来源稳定,易受到人为干扰等方面的影响,与野生种群在行为上存在一定差异[52]。与笼养种群相比,野生种群常在取食和警戒行为方面分配更多的时间,如蓝冠噪鹛(Garrulax courtoisi)[53]、丹顶鹤(Grus japonensis)[54]。本研究通过红外相机辨识并统计出野生勺鸡停歇、警戒、整理、取食、运动、家庭和社会 7类行为,8种姿势,31种动作和21种行为,构建PAE编码系统并建立行为谱。与笼养勺鸡行为研究[22−23]相比,本研究对行为的划分更为细致,两者在停歇、取食及警戒行为方面具有一定差别。停歇行为方面,本研究缺少野生勺鸡高处停歇的行为,该行为的缺失可能与红外相机的拍摄范围相关,红外相机拍摄范围的局限性使其不能捕获到树上栖息的勺鸡[43];取食行为方面,鸟类会根据食物的可获得性调整自身的取食行为[55],与笼养勺鸡相比,野生勺鸡取食生境更为复杂,食物可获得性相对较低,食物种类也更加丰富,因此野生勺鸡取食行为更为多样;警戒行为方面,笼养勺鸡研究中并未提到警戒行为,可能是笼养勺鸡长期暴露于人为干扰环境中对外界干扰形成习惯[56],对环境的适应性增强,从而花费较少的时间用于警戒。
本研究基于建立的行为谱对野生勺鸡不同年龄性别组的行为多样性进行分析,显示野生勺鸡幼体Hvariable、H、r和r-variable 均为最低,一方面,物种行为会随着个体发育才能完整表达[7],因此野生勺鸡幼体的行为种类相对较少;另一方面,野生勺鸡幼体出现的时间区段较短,收集到的数据量较少。整体而言,勺鸡的H和r不存在季节性差异。冬季雌性的H和r均高于雄性,雄性勺鸡体型较大,颜色较雌性更为艳丽,冬季植被的缺乏减少了勺鸡的隐蔽环境,雄性勺鸡可能会降低活动强度以减小被天敌捕食的风险,从而使行为多样性指数相应降低。
本研究结果表明,野生勺鸡的警戒行为在春季处于较高水平,这可能与勺鸡在春季进入繁殖期[20]有关。本研究监测到雄性勺鸡在繁殖前期存在争夺配偶的现象,双雄模式下的野生勺鸡会采取驱逐的方式守护领地,使雄性之间警戒增加。已有研究表明在雌性孵卵期时,雄性会相应提高警戒频次为雌性取食、运动等行为争取时间,为雌性的生存、卵的孵化等提供进一步保障[57−58]。本研究发现,随着6月勺鸡幼体的出现,勺鸡警戒行为随之增加并达到最大值,由于幼体取食经验不足,且被捕食的风险较大,携带幼体的雌性警戒行为会处于较高水平,以提高幼体的取食时间。此外6月勺鸡的运动行为和取食行为频次比例也相应增大。一方面勺鸡幼体需要摄入充足食物以满足自身生长发育的要求;另一方面,勺鸡幼体觅食经验不足,一定程度上使群体觅食效率降低,这都需要勺鸡增加觅食时间[59]。6月中下旬后逐渐出现单独活动的幼体,7月时勺鸡群体的警戒行为和取食行为的频次比例随之下降。
雉类会根据食物条件及个体能量需求,采取最优的取食策略[60−61]。野生勺鸡冬季的运动行为与取食行为频次比例均有增加,且取食行为频次比例达到最大值。冬季植被稀疏,天然隐蔽环境减少,食物资源相对匮乏,野生勺鸡可通过增加觅食时间的方式以弥补食物资源的不足[62],这与绿尾虹雉(Lophophorus lhuysii)[63]、白冠长尾雉(Syrmaticus reevesii)[64]和高原山鹑(Perdix hodgsoniae)[65]等野生雉科(Phasianidae)鸟类的研究结果一致。基于全年数据,野生勺鸡取食行为与警戒行为频次比列呈反比,说明野生勺鸡在取食上分配的时间越长,用于警戒的时间就越短,这与鸟类常见的行为分配方式一致,如白鹇(Lophura nycthemera)[33,59]、绿头鸭(Anas platyrhyn⁃chos)[66]。
虽然本研究优先选择勺鸡活动的生境布设红外相机,但仍然没有监测到理羽、饮水、沙浴和繁殖 4 类行为,在今后的研究中,或可通过样线法调查野生勺鸡这4 类活动发生的生境偏好,在相应区域(水源处、阳坡和营巢地等)增加红外相机的数量和更多的布设方式,扩大研究监测面积,延长拍摄时间,对野生勺鸡的行为进行更全面地监测,进而补充和完善野生勺鸡行为谱和行为谱编码系统。
