高分辨全极化昆虫雷达极化校准与昆虫体轴方向估计
2023-05-05李沐阳李卫东李云龙钱李昌王江涛北京理工大学信息与电子学院雷达技术研究所北京100081卫星导航电子信息技术教育部重点实验室北京理工大学北京100081北京理工大学前沿技术研究院济南250300中国人民解放军31511部队北京100037
李沐阳 胡 程* 王 锐 李卫东 姜 琦 李云龙 钱李昌 王江涛(北京理工大学信息与电子学院雷达技术研究所 北京 100081)(卫星导航电子信息技术教育部重点实验室(北京理工大学) 北京 100081)(北京理工大学前沿技术研究院 济南 250300)(中国人民解放军31511部队 北京 100037)
1 引言
为了觅食和繁衍,全球数以百亿计的害虫会随着季节的变化以一定规律朝不同的方向迁飞[1],这会对全球粮食、果木等农作物生产安全造成极大的威胁。作为迁飞虫害监测的重要传感器,昆虫雷达不仅可以实时对空中目标进行检测,还能给出昆虫的体轴方向、振翅频率等行为参数和体长、体重等体型参数[2–5]。其中,昆虫的体轴方向、体长和体重等参数的计算,均依赖于目标的极化信息。
传统昆虫雷达多采用“极化旋转”的方式获取目标极化信息[6]。测量过程中,雷达波束垂直对天,通过周期性机械旋转天线极化方向获取波束中的目标在不同角度的雷达散射截面积(Radar Cross Section,RCS),也就是“极化方向图”[7]。但是,由于测量体制的限制,传统昆虫雷达的距离分辨率和极化信息获取效率较低,这会导致密集场景下相距较近的目标难以被区分且高速飞行目标的“极化方向图”质量较低,所以传统昆虫雷达的参数反演精度通常较低。
新型的昆虫雷达采用调频步进频波形提高雷达距离分辨率,采用瞬时全极化测量技术提高雷达获取目标极化信息的效率[8],可以极大提高雷达在参数反演方面的性能。高分辨全极化雷达两个极化正交的通道同时发射波形互相正交的调频步进频信号,并同时接收目标回波,最终得到高分辨距离像以及目标的散射矩阵[9]。与昆虫体轴方向、体长以及体重等参数相关的极化信息都包含在散射矩阵中。
但是,由于全极化系统通道间的不一致和交叉串扰等非理想因素的存在[10],全极化雷达直接测量得到的目标散射矩阵存在误差,需要通过极化校准对测量误差进行补偿,从而得到精确的目标散射矩阵。因此,对于全极化雷达,其散射矩阵测量精度和基于散射矩阵的参数反演结果正确性取决于极化校准的精度。
针对合成孔径雷达[11,12](Synthetic Aperture Radar,SAR)和气象雷达[13]等典型的全极化雷达,已有一系列通过测量已知散射矩阵定标体进行极化校准的方法[10,14–20]。但是,对于采用调频步进频体制的高分辨全极化雷达,为了确保高分辨距离像的质量,需要在成像之前对4个极化通道的步进频信号分别进行补偿[21],这会给极化测量模型引入新的通道间幅相不一致误差分量。此外,相比于SAR和气象雷达等典型的全极化雷达,昆虫雷达测量距离较近。在极化校准的过程中,地面定标体回波的信杂比通常较低,而空中定标体姿态通常难以精确控制。为此,本文结合高分辨波形特点对全极化雷达极化测量模型进行了优化,提出了一种以金属球和不需要精确控制姿态的金属丝为定标体的高分辨全极化雷达外场校准的方法。该方法以雷达极化旋转的方式遍历测量各个角度的金属丝,通过金属球初步标定结果确定金属丝角度,然后利用特定角度金属丝测量结果进一步估计系统误差,最终以这种误差逐步估计的方式得到并补偿系统通道间幅相不一致,为基于极化信息的昆虫参数估计提供基础。
在基于散射矩阵的昆虫参数估计方面,目前利用昆虫雷达回波求解昆虫体轴方向时,多依赖于“极化方向图最大值方向对应昆虫体轴方向”[4]的经验假设,缺少模型支撑,且不适用于频率较高的情况。为此,本文基于昆虫生物几何模型的对称性,结合微波暗室测量到的昆虫目标散射矩阵特征,对昆虫散射矩阵重新建模,并给出了适用范围更广的昆虫体轴方向解析计算方法。在此基础上,通过解析推导和仿真分析了交叉串扰对体轴方向测量的影响机制。最后利用高分辨多频全极化雷达进行了极化校准实验,通过对昆虫个体和夜间自然飞行昆虫群体的体轴方向估计结果的分析,验证了极化校准和体轴方向估计方法在多个频段(X,Ku,Ka)的有效性。
2 高分辨雷达极化校准方法
极化校准通过测量散射矩阵已知的定标体求解系统误差,然后利用求解出的系统误差对其他被测目标进行补偿,从而获取目标准确的散射矩阵。而系统极化误差模型会很大程度上影响定标体的选择和系统误差计算方法。因此,在介绍具体校准方法之前,首先对高分辨全极化系统测量模型建模,并针对高隔离度场景对模型进行改进。
2.1 测量误差模型
如图1所示,全极化雷达的接收机、发射机和天线都有两路,分别是H通道和V通道。理想情况下H和V通道的传输特性相同,且H和V通道互相没有影响。但是,在接收机、发射机和天线中,H和V通道通常都难以保持一致。并且,天线的H和V通道之间还存在着泄露。经典的窄带单天线全极化系统测量模型解析形式可以表示为[14]

图1 全极化雷达测量模型Fig.1 Fully polarimetric entomological radar measurement model
其中,r表示目标距离,k0=2π/λ表示波数。Mxy(x,y=h,v)表示目标散射矩阵元素的测量值,Sxy(x,y=h,v)表示目标散射矩阵元素的真实值。Rh,Rv,Th,Tv分别表示射频链路中H接收通道、V接收通道、H发射通道和V发射通道导致的幅相不一致。S13和S24表示天线H通道和V通道导致的不一致;C1和C2表示天线H通道和V通道间的交叉串扰,C1和C2的幅度远小于1。对于一个新的系统Rh,Rv,Th,Tv,S13,S24,C1和C2都是未知的,这些参数可以统称为系统极化误差。
对于采用调频步进频的高分辨全极化系统,在式(1)中所展示的极化误差之外,还存在着由于4个极化通道(HH,HV,VH和VV)距离像成像效果不同引入的误差。由于每个通道的传输特性都是非理想的,每个极化通道的回波都需要补偿脉冲间幅相误差、脉冲内幅相误差以及脉冲间时延才能保证成像效果[21]。为测定雷达发射步进频信号的脉冲间幅相误差、脉冲内幅相误差以及脉冲间时延,通常需要对额外的标准目标进行测量。但是,通常同一个目标难以满足4个通道的测量需求,因此每个极化通道测量脉冲间幅相误差、脉冲内幅相误差时所使用的标准目标也会有差异,测量场景也会有所不同,这些会导致最终4个极化通道的距离像成像效果有所不同。这相当于为每个极化通道重新引入了一个乘性系数,且通常不同通道的系数不同。因此,针对调频步进频体制的高分辨全极化雷达系统,其极化测量模型可以建模为为了降低极化校准的复杂程度,针对隔离度较高的系统,可以在保证参数精度的情况下忽略C1和C2(后文给出了具体分析)。忽略交叉串扰之后,新的系统模型可以表示为
定义:
则式(4)可以重新表示为
与传统窄带测量模型相比,式(6)所表示的系统测量模型具有更复杂的通道间幅相不一致,传统单目标极化校准算法[14]不再适用。此外,因为模型简化过程中忽略了系统的交叉串扰,所以该模型与高隔离度系统更加相符。
昆虫体轴方向与散射矩阵强度无关,只需要4个通道的相对大小关系即可求解体轴方向。所以,对于式(6)所表示的系统模型,只需要估计出和,然后对测量结果进行补偿,确保各个极化通道之间具有准确的相对关系,完成极化校准。校准后的散射矩阵可以用来计算昆虫体轴方向。其中,g1可以利用金属球进行标定,其标定过程与本文主线无关,因此后文不再赘述。
2.2 系统极化误差校准
根据系统测量模型选取能够求解系统误差且便于测量的定标体是极化校准的关键。通过对式(6)的分析可知,若想获取准确的各个极化通道之间的相对关系,只需要估计出和。一种直观的解决方法是,测量一个散射矩阵已知的目标,记为
其中,x和y不同时为h。
对于昆虫雷达,金属球各向同性,从各个角度测量到的散射矩阵均是相同且已知的,是一种合适的可在空中测量的定标体。但是其交叉通道元素理论值为0,实际回波较弱,难以被用于估计交叉通道的幅相不一致系数和。金属丝的散射矩阵会随视角变化,存在某些角度金属丝交叉通道和主通道回波都较强,可以用于对通道间幅相不一致进行标定,但其角度通常难以准确控制。本文联合金属球和金属丝作为定标体,首先利用金属球进行初步标定,在此基础上定位金属丝角度,最后利用特定且已知角度的金属丝回波对雷达进行极化校准。
2.2.1 金属丝角度定位
金属丝的散射矩阵是随横滚角变换的。当金属丝与垂直于入射波的平面平行时,其散射矩阵可以表示为
其中,θ为金属丝在垂直于入射波平面上的横滚角,是相对于H极化的角度,Aline为常数,与金属丝的结构和材料相关。(·)T表示矩阵的转置。Rθ为旋转矩阵,其具体形式为
理想情况下,δ极小(后文可以看出,δ约为–30 dB),可将其忽略,则Sline(θ)可表示为
此时,金属丝4个极化通道的幅度随横滚角变化规律如图2所示。
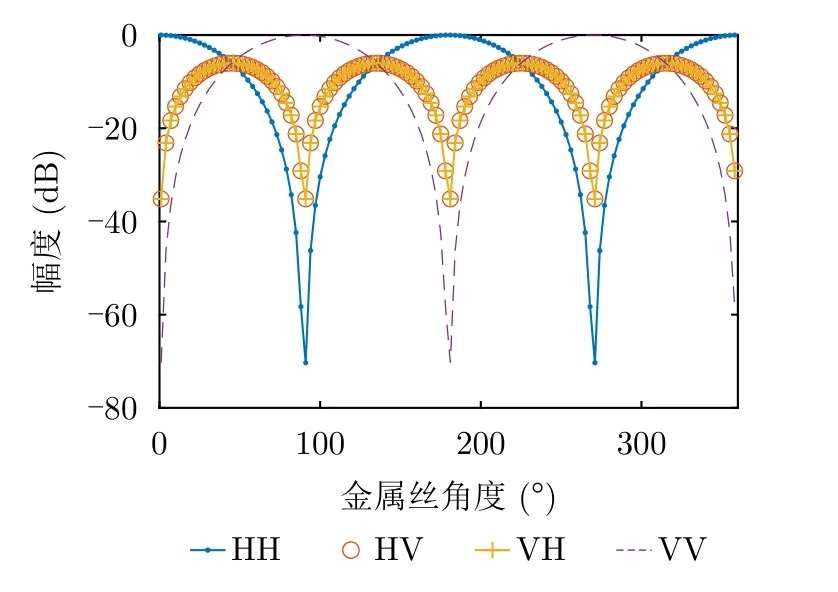
图2 不同角度时金属丝散射矩阵4个通道的幅度Fig.2 Amplitude of the scattering matrix elements at different angles
金属丝在空中的横滚角通常难以准确固定,所以在利用金属丝回波标定系统误差之前,首先需要确定金属丝角度。通过式(11)和图2可以看出,在角度θc=π/4+πn/2(n∈Z)处,金属丝HH和VV通道的回波幅度相同,因此可通过定位HH和VV通道幅值相同的角度确定金属丝角度。
其中,σball为金属球的RCS。根据式(7)和式(12)可知,雷达测量到的金属球散射矩阵为
金属丝散射矩阵测量结果可表示为
2.2.2 系统极化误差估计和补偿
实际操作过程中,可以控制雷达垂直对天观测,然后利用两架无人机通过细线拉扯将金属丝携带至波束中心且近似与H极化方向平行的位置,并在方位角旋转的过程中持续采集不同角度金属丝的散射矩阵。
假设最初金属丝与雷达H极化方向近似平行,雷达方位角逐渐增加,当的HH和VV通道回波第1次相同时,金属丝相对于雷达H极化方向为–45°,金属丝真实散射矩阵为
则测量到的散射矩阵为
3 基于生物几何模型的体轴方向估计方法
昆虫体轴方向估计是极化信息在昆虫雷达上的一种重要应用。昆虫体轴方向表示昆虫的主动飞行方向,结合风向和风速等信息,可以对昆虫的飞行轨迹和飞行规律进行分析,这对迁飞虫害爆发的预警以及昆虫飞行特性的探索至关重要。在完成极化校准的基础上,可利用准确的目标散射矩阵求解昆虫轴向。但传统体轴方向估计方法多依赖于“极化方向图最大值方向对应昆虫体轴方向”的经验假设[4],该假设缺少物理意义上的解释且误差随着频率升高逐渐增加。本文通过对暗室昆虫数据的分析,利用对称模型对昆虫散射矩阵进行建模,并给出了在较高频段同样适用的昆虫体轴方向估计方法。
3.1 昆虫散射矩阵模型
从生物外形几何结构出发,当从昆虫腹部视角进行观测时,大多数昆虫关于其体轴对称[22]。目标的散射矩阵也能反映目标的对称程度,通常可以用θsym衡量目标对称程度[23]:(A,B)表示向量A和B的内积,∥A∥表示矢量A的二范数。θsym越小表明目标对称程度越高。研究表明暗室中测量到的绝大部分昆虫目标θsym都小于π/50[24],这说明大部分昆虫对称的程度都很高。
可以证明,双边对称目标对称轴平行于雷达H或V极化方向时,散射矩阵交叉通道为0[22]。定义昆虫体轴平行于H极化时散射矩阵为
如图3,当昆虫体轴与H极化方向夹角为θ时,昆虫散射矩阵可以重新表示为

图3 昆虫体轴与极化方向示意图Fig.3 Schematic diagram of orientation and polarization direction
当昆虫体轴方向平行于V极化方向时,θ为0 .5π,此时昆虫散射矩阵可表示为
可以看出,相较于昆虫平行于体轴时的散射矩阵,垂直于体轴时的昆虫散射矩阵交叉通道仍为0,但HH与VV通道元素位置互换。
可以看出依据昆虫几何模型给出的散射矩阵所包含的未知参数只有s1,s2和θ。其中θ与昆虫体轴方向相关,s1和s2与昆虫体型相关。该模型舍弃了目标散射矩阵中较小的非对称分量,将昆虫体轴求解转化为求解对称目标对称轴,物理意义更加清晰。
3.2 模型驱动下的体轴方向计算方法
通过式(25)和式(30)可以看出,当昆虫平行或垂直于极化方向时,散射矩阵交叉通道为0。因此,可以通过最小化的交叉通道,求解昆虫体轴方向与雷达极化方向的夹角:
3.3 方法对比
传统昆虫体轴估计方法认为“极化方向图最大值”方向为昆虫体轴方向[4]。对于采用单一线极化天线的雷达,极化方向图指的是昆虫RCS随体轴与极化方向夹角变化的曲线。全极化雷达可以通过散射矩阵计算目标极化方向图,进而找到RCS最大的方向,并确定昆虫体轴方向。
为了对比验证新体轴估计方法的有效性,基于暗室测量到的昆虫散射矩阵数据集[25]进行了仿真。仿真过程中,每只昆虫进行500次体轴方向估计,进行轴向计算之前昆虫的体轴方向随机设置,并通过式(26)获取昆虫体轴在不同方向时的散射矩阵,信噪比恒定为20 dB,最后通过式(44)计算昆虫体轴方向。昆虫的体轴方向估计误差定义为
其中,orimeasured为计算得到的昆虫体轴方向,orireal为昆虫真实轴向。仿真过程中分别通过“极化方向图最大值方向”和几何对称模型两种方法估计昆虫体轴方向,并与已知真实方向对比得到估计误差。
图4给出估计得到的所有昆虫体轴方向测量的平均误差以及误差较大(>10°)的昆虫数目,其中误差较大的昆虫被认为是超出对应方法适用范围。可以看出,平均体轴方向误差与大误差昆虫数目相关性较高,这说明大误差昆虫的数目很大程度上决定了平均误差。通过两种方法在不同频率时的昆虫体轴方向平均测量误差可以看出,“极化方向图最大值方向”方法的误差在所有频点都大于基于几何对称模型的方法。这是因为对于样本中的大型昆虫,前者往往失效,并造成最终估计误差较大。并且,随着频率升高,传统方法的误差和大误差昆虫数目会逐渐增大,这说明在高频段会有更多的昆虫不再满足“极化方向图最大值方向对应体轴方向”这一假设。而基于几何对称模型的方法适合更多数量的昆虫,且估计误差在各个频段都相对较小,平均误差都小于10°。
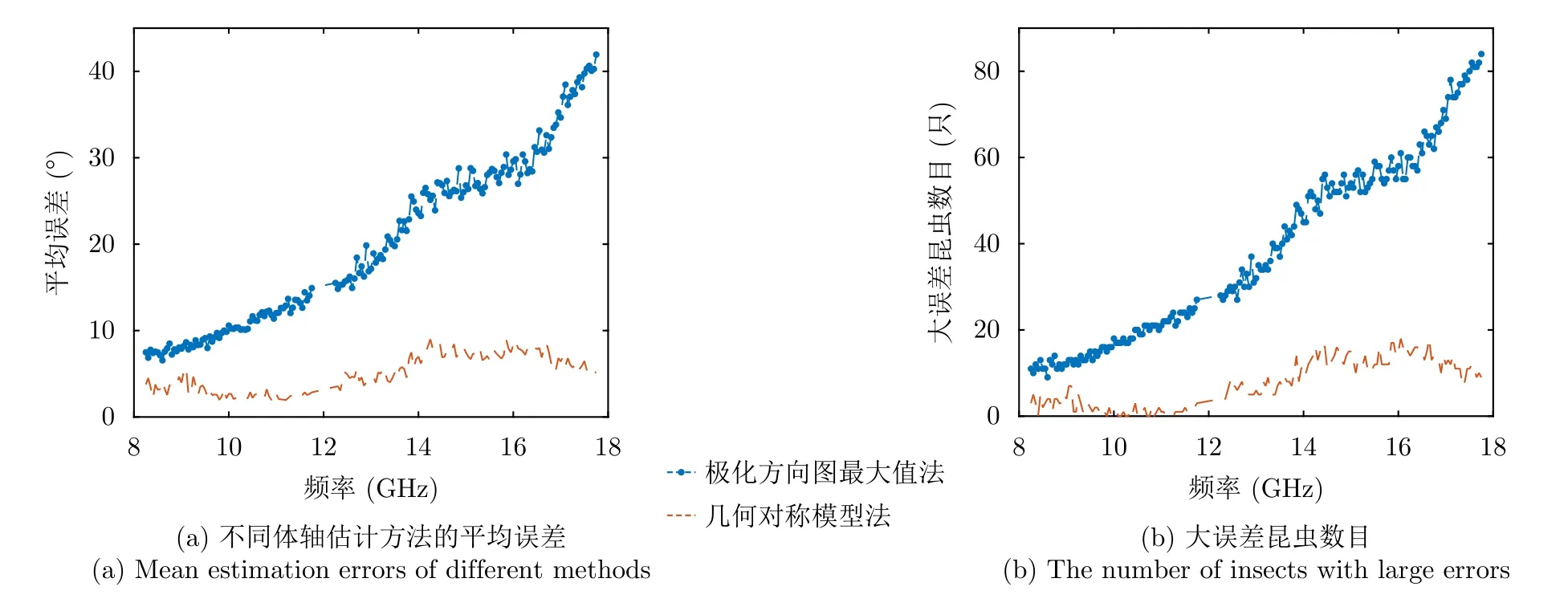
图4 不同频率时的昆虫朝向误差统计结果Fig.4 Statistical results of insect orientation errors at different frequencies
4 误差分析和方法验证
第2节和第3节分别介绍了一种高分辨全极化雷达极化校准方法和基于生物几何对称模型的昆虫体轴方向估计方法。但为了方便极化校准的实现,在系统建模的时候忽略了交叉串扰的影响。为此,在用外场实测数据评估两种方法性能之前,首先理论分析了忽略交叉串扰对体轴方向估计的影响。
4.1 交叉串扰对体轴方向测量的影响
对于昆虫雷达所采用的双极化抛物面天线,天线在不同空域处的极化特性不同。在主瓣中时,天线交叉串扰的方向图通常在0°时趋于−∞,并随着角度增加而强度增加[26]。因此,当目标在波束中心时,C1和C2趋于0,甚至可以忽略,此时系统模型等同于式(4)和式(6)。同时,由于极化校准过程中,通常会控制定标体位于波束中心。因此,测量定标体时不会受到交叉串扰的影响,利用定标体估计出的和是准确的。
随着目标逐渐偏离波束中心,C1和C2的强度逐渐增加,此时用式(3)描述系统则更加合理,这里重写式(3):
可以看出,利用本文的方法进行极化校准之后,只有C1和C2会影响散射矩阵。
将式(47)代入式(36),可以得到包含交叉串扰的体轴方向测量结果为
在雷达单站且目标无源的情况下,目标通常满足互易性,目标散射矩阵交叉通道相等,也就是Shv=Svh。则式(48)可以表示为
on表示高阶误差项,化简后可以表示为
取实部并解模糊之后为
从式(52)可以看出,在交叉串扰的影响下,测量得到的体轴方向分为4部分,分别是目标真实体轴方向、固定角度误差、被忽略的高阶项以及随昆虫自身角度变化的误差。
可以看出,交叉串扰从两个方面影响体轴方向估计。当C1和C2不同时,测出来的体轴方向会存在一个固定的角度误差,这个角度的大小为
另一方面,C1和C2会共同带来一个随着昆虫体轴方向余弦函数变化的误差:
该误差峰值大小同时与交叉串扰以及昆虫自身散射特性相关。通过式(54)可以看出,该误差在体轴方向为45°或–45°时较小,在体轴方向为0°或90°较大。把固定角度误差和变化的误差结合起来,最终测量出的体轴方向误差为θerr0+θerr1。
大量仿真表明式(52)可以准确表示昆虫体轴方向测量误差,本文以一只棉铃虫X波段暗室测量为例,通过两组仿真说明式(52)的准确性。第1组仿真中C1=C2=0.055ejπ/8,第2 组实验中C1=0.055ejπ/8=2C2。仿真过程中,首先通过式(26)将目标旋转至不同的角度,然后一方面通过仿真计算交叉串扰带来的体轴方向估计误差,另一方面计算通过1阶泰勒展开和2阶泰勒展开并忽略高阶项后的体轴方向估计误差。仿真结果如图5。
可以看出,通过泰勒展开计算得到的昆虫体轴方向误差与仿真结果近似,差异相对较小,并且1阶展开结果与2阶展开结果类似。此外,对比两次仿真可以看出,当C1=C2时,体轴方向误差在0°附近波动;当C1≠C2时,体轴方向误差则在一个近似余弦周期的波动外,还有存在着一个固定偏差。这些都与式(52)所展示出的结果一致。此外,由于体轴方向误差随着体轴方向的变化而不同,最终统计结果会偏向0.5real(C2−C1)+π/4或0.5real(C2−C1)−π/4。图6给出了基于暗室测量到的X波段183只昆虫散射矩阵数据集[3]进行的仿真,仿真方式也与图5对应的仿真类似,首先通过式(26)将目标旋转至不同的角度,然后通过计算特定交叉串扰下的体轴方向估计结果并进行统计,其中第1组仿真中C1=C2=0.055ejπ/8,第2组实验中C1=0.055ejπ/8=2C2。可以看出,在引入交叉串扰的影响之后,体轴方向统计结果朝45°附近的方向集中。

图5 交叉串扰对昆虫体轴方向估计的影响Fig.5 Influence of cross-talk on the estimates of insect orientation

图6 交叉串扰对虫群体轴方向分布的影响Fig.6 Influence of cross-talk on the orientation distribution of insect swarms
表1给出了多组不同交叉串扰下的虫群的体轴方向误差。其中第1组C1和C2相同,且幅度近似为25 dB;第2组C1和C2相位相同,幅度不同;第3组C1和C2幅度相同,相位不同。可以看出,当C1和C2相同时,体轴估计误差均值近似为0°。当C1和C2幅度或相位有差异时,体轴估计误差均值不再为0,这与前文的分析一致。在仿真场景中,体轴估计误差都不超过5°。

表1 不同交叉串扰时昆虫体轴方向估计误差Tab.1 Orientation estimation error under different cross-talk
4.2 个体和群体昆虫观测
4.2.1 实验场景
为了验证极化校准方法和体轴方向估计方法的有效性,我们利用一部多频全极化雷达进行了极化校准实验并对单只昆虫进行了吊飞测试。该雷达共5个频段,实验过程中我们利用了X1,X2,Ku1,Ku2和Ka频段的极化信息以及Ka波段的测角信息,具体参数见表2。

表2 多频全极化雷达参数Tab.2 The parameters of multi-frequency fully polarimetric radar
金属丝数据采集场景如图7。在无人机飞行之前利用少量胶水将金属丝和细线平行固定。无人机飞行过程中,两无人机停留在相同高度,并拉扯细线至绷紧。实验在微风条件下进行,目标和无人机位置相对比较平稳。实际数据采集过程中,首先控制雷达俯仰角为90°(垂直对天),方位角为0°,此时H极化与东西方向平行。利用两架无人机通过120 m长细线将金属丝带至雷达正上方约480 m处,金属丝近似与雷达H极化方向平行。然后雷达在方位角旋转的过程中,持续获取金属丝的散射矩阵。金属球的测量场景与金属丝相同。共进行了3组昆虫测量实验,测试过程中昆虫高度分别为480 m,440 m和400 m,其他场景与金属丝相同,此外在昆虫测量过程中,飞机携带实时动态测量(Real Time Kinematic,RTK)设备精确记录飞机位置信息。

图7 定标体和昆虫测量场景Fig.7 Calibrators and insects measurement scene
为确保无人机和细线回波不会干扰到金属丝回波,我们还额外进行了一组不携带金属丝或昆虫且其他实验场景完全相同的实验。图8给出了X1频段细线与雷达H极化方向近似平行时雷达在两种场景下获取到的HH通道距离像。可以看出,当不存在金属丝时,距离像上没有明显目标,这说明无人机和细线的回波被噪声完全淹没,因此不会影响后续获取到的金属丝回波。对于其他频段,由于频率更高,天线波束更窄,无人机和细线的回波对距离像的影响更小,因此同样被噪底淹没。
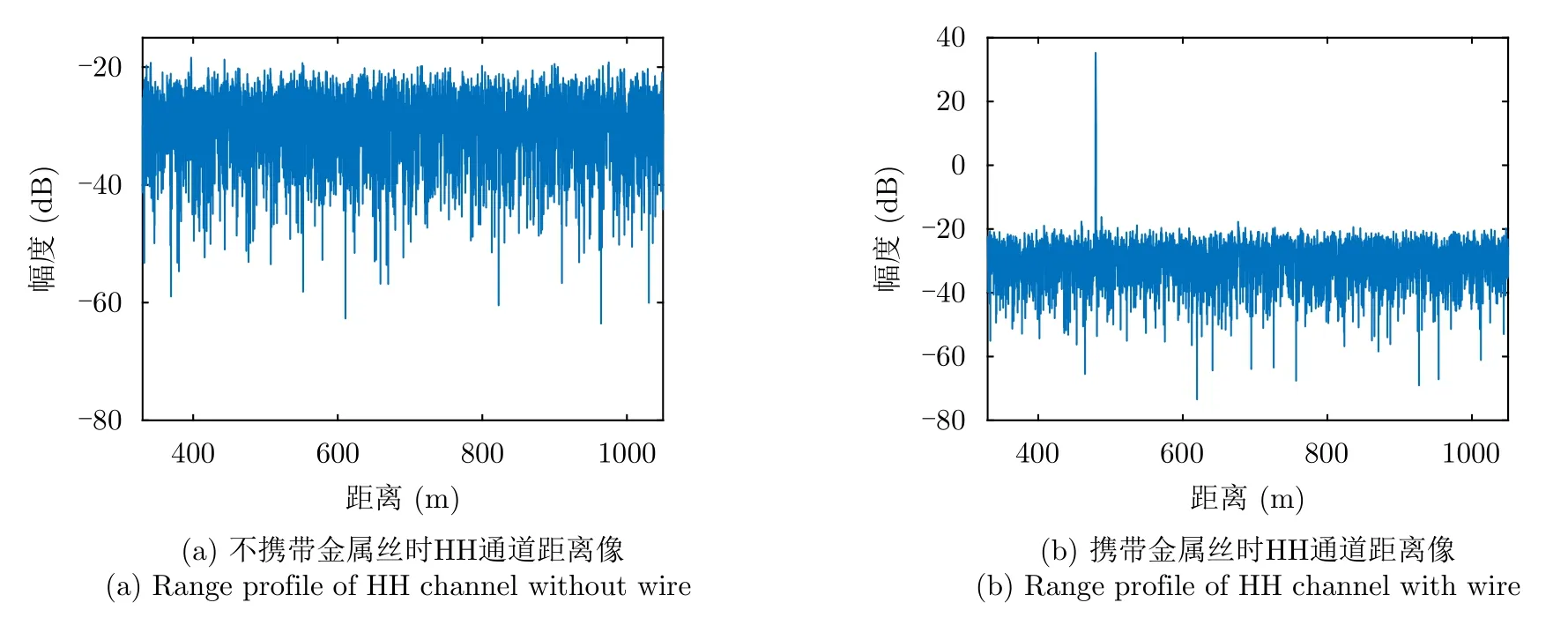
图8 不同实验场景下HH通道距离像Fig.8 HH channel range profile under different experimental scenarios
4.2.2 极化校准结果
实验过程中使用的金属丝直径为1 mm,长度为200 mm;金属球直径为60 mm。以X1频段为例,采集到的金属丝4个通道的回波幅度如图9(a)所示。

图9 金属丝散射矩阵4个通道幅度测量结果Fig.9 The measured amplitude of wire scattering matrix elements
经过金属球标定,在方位角为42.8°时HH和VV通道幅度相同,根据2.2节的分析,此时金属丝相对于H极化方向为–45°。可以提取此时的金属丝散射矩阵作为–45°金属丝测量结果,用于校准其他数据。
为了验证校准效果,我们对金属球重新进行了测量,并利用金属丝数据对该组金属球散射矩阵进行校准。理论上金属球散射矩阵为对角矩阵,HH和VV通道相同。定义校准前后的金属球散射矩阵分别为Mball和
4.2.3 体轴方向估计结果
利用金属丝校准昆虫数据,并通过式(44)计算得到其中1组昆虫体轴方向随雷达方位角变化的曲线如图10(a),这里的体轴方向指的是昆虫真实体轴方向与雷达H极化方向的夹角。
可以看出,估计出的昆虫体轴方向随方位角线性变化。由于在测量过程中两架无人机的位置信息被准确记录,可假设两架无人机与昆虫在同一条直线上,从而计算出昆虫的真实体轴方向。根据昆虫真实体轴方向可以计算得到在不同方位角时昆虫相对于H极化的角度,进而可以与体轴方向测量结果对比计算出体轴方向估计误差,如图10(b)。

图10 昆虫方向测量结果Fig.10 Insect orientation measurement result
3组昆虫测量实验中各个频段的体轴方向估计误差见表3。可以看出,3组实验结果中的角度估计误差均值都很小,接近于0,角度估计误差最大值都小于3°。

表3 不同频段昆虫体轴方向测量误差Tab.3 Measurement error of insect orientation in different frequencies
夜间观测到的虫群体轴方向分布也从侧面反映了极化校准和体轴方向估计结果的正确性。图11给出了校准前后整个夜间(19:00—次日7:00)观测到的昆虫体轴方向的分布。可以看出,在校准之前各个频段统计出的昆虫体轴方向分布完全不一致,且没有规律。校准之后,各个频段统计出的体轴方向均集中在0~20°区间中。但是,由于不同频段的威力不同,每个频段检测到的昆虫数目不是统一的,因此不同频段的体轴方向分布略有差异。
此外,还统计了利用Ka频段测角功能计算出的昆虫轨迹方向。夜间观测过程中,昆虫雷达通常波束垂直对天,当目标穿过波束时,可以利用测角结果判断昆虫的轨迹方向。通常,在无风或风速较小的情况下,昆虫体轴方向会与运动轨迹方向保持一致。对比图11和图12可以看出,统计出的昆虫体轴方向与运动轨迹基本一致,均集中于0~20°范围。这从侧面说明了昆虫体轴方向计算结果的有效性。

图11 夜间观测到的虫群体轴方向分布Fig.11 Swarm orientation distribution observed at night

图12 昆虫轨迹方向分布Fig.12 The distribution of insect trajectory direction
4.3 性能分析
通过对比表1和表3可以看出,表3所统计的实验误差要整体小于表1所统计的仿真误差。实验测量结果的体轴角度估计误差均值明显小于第2组和第3组仿真结果,实验测量到的体轴角度估计误差最大值要明显小于仿真结果。这主要是因为在仿真中,系统交叉串扰被设置为0.055(近似25 dB),而实际系统在3 dB波束中的交叉串扰要优于25 dB。结合对式(46)和式(47)的分析可知,在工程应用中,交叉串扰是主要影响体轴方向测量精度的因素。对于高隔离度的系统,系统通道间交叉串扰更小,极化校准后散射矩阵受到系统交叉串扰的影响较小,轴向测量误差也更小。对于低隔离度系统,则需进一步进行误差评估。
极化校准后的轴向测量误差可以表示为
其中,θerr0与交叉串扰C1和C2的差异相关,θerr1同时受到Aerr和目标所在角度θ的影响。假设被测目标体轴方向均匀分布,θerr1是随目标体轴方向周期性变化的分量,从而不会影响体轴估计误差均值。此时,体轴估计误差均值只与θerr0相关。对于最优的情况,C1和C2相同,此时体轴方向估计误差均值极小,这一结论与表1第1组仿真结果相符。对于最恶劣的情况,C1和C2幅度相同但相位相反,此时目标体轴方向估计误差均值与系统交叉串扰的关系如图13。实际情况中体轴方向误差均值介于两种极端情况之间。

图13 最差情况下θerr0与系统隔离度的关系Fig.13 The relationship between θerr0 and system isolation in the worst case
Aerr取决于交叉串扰和目标散射特性,可通过式(54)进行计算。对于某一确定目标,若θerr0>0,当cos 2θ=1时,目标体轴方向测量误差绝对值最大,为θerr0+Aerr;若θerr0<0,当cos 2θ=−1时,体轴方向测量误差绝对值最大,为−θerr0+Aerr。但由于Aerr的情况比较复杂,通常可以通过仿真或实验对体轴方向测量误差最大值进行评估。对于本文的仿真场景,如表1所示,体轴方向测量误差小于5°;在实验中,如表3所示,5个频段的体轴方向测量误差均小于3°。
5 结语
本文首先对高分辨全极化雷达的极化测量模型进行建模,然后针对新的测量模型设计了一种基于松姿态约束下双定标体(金属球和金属丝)联合的高分辨全极化雷达极化校准方法。在极化校准的基础上,提出了一种基于生物几何对称模型且适用范围更广的昆虫体轴方向估计方法,并分析了交叉串扰对体轴方向估计的影响机制。为了验证这两种方法的性能,利用一部多频高分辨全极化雷达进行了极化校准和昆虫个体的轴向测量,实验结果表明新的极化校准方法在多个频段(X,Ku,Ka)均可以很好地补偿系统极化误差,且补偿之后昆虫体轴方向测量误差不超过3°;此外,校准之后,夜间观测到的昆虫体轴方向与昆虫飞行轨迹契合度很高。
随着全极化/压缩极化测量和校准技术的成熟,极化信息逐渐被不同类型的雷达应用于目标检测、杂波抑制以及参数反演等任务。同时,极化信息与多频、多视、空间等其他域的信息的联合处理正逐渐成为研究热点。后续将针对全极化信息与多频段信息的深度融合进行研究,以提高包括昆虫体轴方向在内的不同类型参数的反演精度。
