基于碳氮稳定同位素的太湖鱼类营养生态位研究
2023-04-29刘素群申明华刘学勤
刘素群 申明华 刘学勤
摘要:为探究太湖鱼类的营养生态位特征,2019-2020年开展了鱼类碳氮稳定同位素调查,运用贝叶斯稳定同位素椭圆模型解析了鱼类的营养生态位大小及生态位重叠,运用双基线法分析了鱼类的营养级。结果显示,鱼类δ13C值为-27.67‰~-17.92‰,δ15N值为6.02‰~20.31‰。营养生态位大小(SEAc值)为0.14‰2~20.43‰2,其中黄颡鱼(Pelteobagrus fulvidraco)的生态位最大,光泽黄颡鱼(Pelteobagrus nitidus)的生态位最小。刀鲚(Coilia nasus)、陈氏短吻银鱼(Salangichthys tangkahkeii)和鲢(Hypophthalmichthys molitrix)的SEAc值均较小,表明其可利用的食物源和生境范围较窄,鳙(Aristichthys nobilis)的SEAc值较大,表明其可利用的食物源和生境范围较广。鱼类的营养生态位重叠度为0~70.6%。大银鱼(Protosalanx hyalocranius)与翘嘴鲌(Culter alburnus)的生态位重叠度最大(70.6%),其次为中国花鲈(Lateolabrax maculatus)与翘嘴鲌(39.9%),其余物种间的生态位重叠度为0~38.9%。刀鲚与其他肉食性鱼类的生态位重叠度较小,表明二者间的食物资源竞争较弱。陈氏短吻银鱼与鲢、鳙在生态位上存在明显的分离,这与陈氏短吻银鱼专食浮游动物有关。鱼类营养级为0.73~3.62,鮠(Leiocassis sp.)的营养级最高,泥鳅(Misgurnus anguillicaudatus)的营养级最低。研究表明,鱼类营养生态位大小与其摄食的食物种类以及对不同生境的利用程度有关,大部分鱼类之间的生态位重叠度较低,说明其在资源利用上存在较大分离。研究结果可为太湖鱼类食物网分析及渔业资源管理提供基础数据。
关键词:稳定同位素;营养生态位;生态位重叠;营养级;太湖鱼类
中图分类号:Q148 文献标志码:A 文章编号:1674-3075(2023)05-0076-08
生态位是生态学研究的重要内容(朱春全,1997)。目前,有关生态位的定义被普遍接受的是Hutchinson(1957)提出的超体积生态位(hypervolume niche)概念框架,其将生态位定义为“所有的能够允许个体或物种无限期地存在的变量集合”。营养生态位指个体或物种对食物资源能够实际和潜在占据、利用或适应的部分,营养生态位不仅体现了物种自身的营养需求,同时反映了物种在生态系统中所处的营养层次(地位)和功能(康乐和陈永林,1994)。营养生态位研究对于理解物种的食物资源利用、种间相互作用以及食物网结构与功能(如物质循环与能量流动)均有重要作用(Begon et al,2006);同时,生态位研究也可为生态系统管理提供理论支撑,如应用于外来种控制、渔业资源管理、生态修复等(Taylor et al,2018;Vasquez et al,2021;Wainright et al,2021)。
营养生态位包括生态位大小(宽度)、生态位重叠和营养级,其定量研究主要基于食性分析。在鱼类生态学中,胃肠内含物分析及稳定同位素示踪是两种主要的食性分析方法(Nithirojpakdee et al,2014)。胃肠内含物分析可直接观察鱼类摄取的食物种类,但存在一定的偶然性,不能反映鱼类的长期食性。稳定同位素分析可反映鱼类长期的摄食情况以及对不同生境的利用,但对食物种类的确定较为粗略(王姝然,2019)。在生态位研究中,基于碳、氮等稳定同位素的分析方法已较为成熟,在指标的量化上也较为有效和准确(Rahman et al,2021;Wang et al,2021)。
太湖是长江中下游典型的浅水湖泊,也是我国重要的渔业生产基地。近年来,受江湖阻隔、富营养化和过度捕捞等影响,太湖渔业资源不断衰退,鱼类组成也发生了较大变化(张彤晴等,2016)。这些环境干扰必然会对其营养生态位产生影响。目前,针对太湖鱼类食物网结构及Ecopath模型有较多研究(徐德琳,2013;李云凯等,2014;Li et al,2018)。然而,现有研究多关注鱼类的营养级特征,对生态位大小及生态位重叠等缺乏分析;同时,大部分研究采用的是2010年以前的采集数据。近10年来,太湖水环境及鱼类群落均发生了较大变化(张翔等,2021),因此,有必要开展太湖鱼类营养生态位研究,以了解鱼类对不同生境的利用情况。鉴于太湖于2020年10月开始实行“十年退捕”,整个太湖水域全面退出生产性捕捞,本研究于2019-2020年对太湖鱼类的碳氮稳定同位素进行调查,分析鱼类的营养生态位特征,以期为太湖鱼类食物网研究、渔业资源管理及“十年退捕”后的生态修复效果评估提供基础资料及科学依据。
1 材料与方法
1.1 研究区域概况
太湖(30°55'40"~31°32'58" N,119°52'32"~120°36'10" E)地处长江三角洲南缘,横跨苏浙两省,北临无锡,南濒湖州,西依宜兴,东近苏州、吴江,湖泊面积约2 338 km2,湖岸线全长约393 km,平均水深1.9 m(秦伯强和罗潋葱,2004)。
1.2 样品采集与处理
分别于2019年10-11月和2020年9-11月开展鱼类稳定同位素样品采集与处理(王姝然,2019;徐军等,2020)。鱼类样品主要在苏州市相城区的望渔村、吴中区的渔港村以及2个渔村之间的沿岸渔码头采集;其中,渔港村是太湖最大的渔港。渔民使用的渔具主要有渔簖、刺网、地笼。收集不同种类的渔获物,鱼类经分类鉴定、个体体长和体重测量后,取背部肌肉用于稳定同位素分析。个体较小的鱼类取3~5尾肌肉混合后作为1个样品,个体较大的鱼类每尾取1个样品。在渔获物调查中,同时收集了螺类和蚌类等底栖动物,作为营养级计算中的基线生物。收集的螺类主要为环棱螺(Bellamya sp.),蚌类主要为河蚬(Corbicula fluminea)。环陵螺取腹足肌肉,河蚬取斧足肌肉。所有样品冷冻保存,带回实验室在60℃下烘干至恒重。烘干的样品经碾磨后放入干燥器中保存,用于稳定同位素分析。鱼类样品共采集132份,螺类和蚌类样品共21份。
肌肉样品经过均质碾磨,用4 mm×6 mm或5 mm×9 mm的锡杯包裹,送入Delta V Advantage同位素质谱仪(Thermo Fisher Scientific,Germany)进行碳氮稳定同位素测定,参考标准分别是美国拟箭石(PDB)和空气中的氮气(N2),标准品分别是USGS40和USGS41。稳定同位素样品的测定在中国科学院水生生物研究所分析测试中心进行。测定结果以δX形式表示(Peterson & Fry,1987),计算公式如下:
δX=(Rs/Rst-1)×1000‰ ①
式中:X代表13C或15N,Rs和Rst分别为测定样品和参考标准样品的同位素比值(13C/12C或15N/14N)。样品分析误差在±0.3‰以内。
1.3 数据分析
对于生态位大小的量化,Layman等(2007)提出了基于碳氮稳定同位素双坐标图的生态位总面积(TA,‰2)指标,计算δ13C~δ15N双坐标图中样点围成的凸包面积。该指标反映了物种或群落占据营养生态位空间的总量,但受样本量的影响较大。为克服样本量的影响,Jackson等(2011)随后提出了标准差椭圆面积(SEAc,‰2)指标,该指标计算δ13C~δ15N双坐标图中样点的标准差椭圆面积,包含40%置信区间的数据,反映了物种或群落占据多维空间的核心区域大小。由于采集的样本量较小(3~12个),本研究同时采用TA和SEAc定量分析鱼类生态位大小,TA仅用于群落生态位量化,SEAc同时用于群落和物种生态位计算。运用贝叶斯稳定同位素椭圆模型(R软件SIBER包)计算了物种及群落的生态位大小,因SEAc计算要求样本数量不低于3个(Jackson et al,2011;R Core Team,2021),故本研究仅对满足计算要求的19种鱼类进行了分析。营养生态位重叠度计算公式如下:
D = S0 / S ②
式中:D为物种间的营养生态位重叠度,S0为物种1标准差椭圆和物种2标准差椭圆的重叠区域面积,S为物种1和物种2中较大的标准差椭圆面积(Jackson et al,2011)。
营养级计算采用Post(2002)提出的双基线法。本研究中采用环棱螺和河蚬分别作为碎屑食物链和浮游食物链基础计算鱼类的营养级(Trophic position,TP),计算公式如下:
TP = λ+{δ15Nc-[δ15Nb1×α+δ15Nb2×(1-α)]}/F ③
式中:λ为基线生物的营养级(λ=2,本研究中基线生物为初级消费者);δ15Nc为鱼类的δ15N值;F为δ15N的营养富集度,取值为3.4‰(Post,2002);α为鱼类能量来自于浮游食物链的比例,计算公式如下:
α = (δ13Cc-δ13Cb2)/(δ13Cb1-δ13Cb2) ④
式中:δ13Cc为鱼类的δ13C值;δ13Cb1和δ15Nb1分别为河蚬的δ13C和δ15N值;δ13Cb2和δ15Nb2分别为环棱螺的δ13C和δ15N值。
数据分析在R 4.0.4和Excel 2019中完成。
2 结果
2.1 鱼类碳氮稳定同位素分布
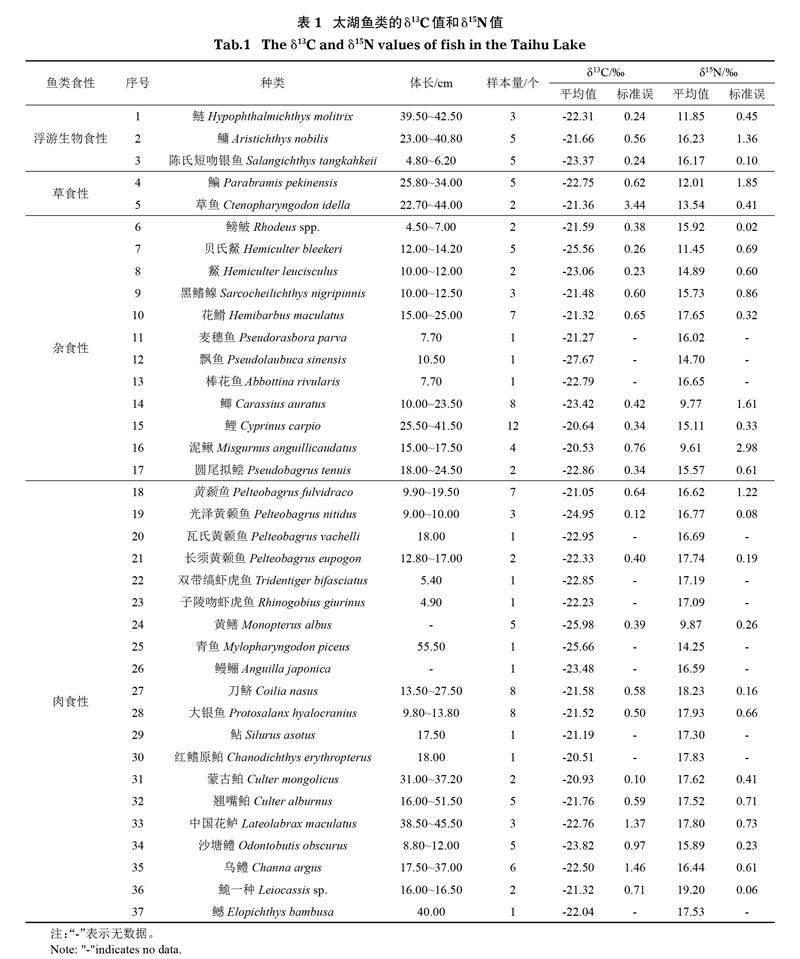
本次调查共采集鱼类37种(表1)。鱼类δ13C值为-27.67‰~-17.92‰,δ15N值为6.02‰~20.31‰(表1、图1)。δ13C均值最大的为红鳍原鲌(Chanodichthys erythropterus)(-20.51‰),最小的为飘鱼(Pseudolaubuca sinensis)(-27.67‰)。δ15N均值最大的为鮠一种(Leiocassis sp.)(19.20‰),最小的为泥鳅(Misgurnus anguillicaudatus)(9.61‰)。不同摄食类群之间δ13C值差异不明显(P=0.92),δ15N值差异显著(P<0.001)。大部分鱼类样点聚集,小部分鱼类飘鱼、黄鳝(Monopterus albus)、泥鳅、鲫(Carassius auratus)等样点离散(图1)。
2.2 鱼类营养生态位
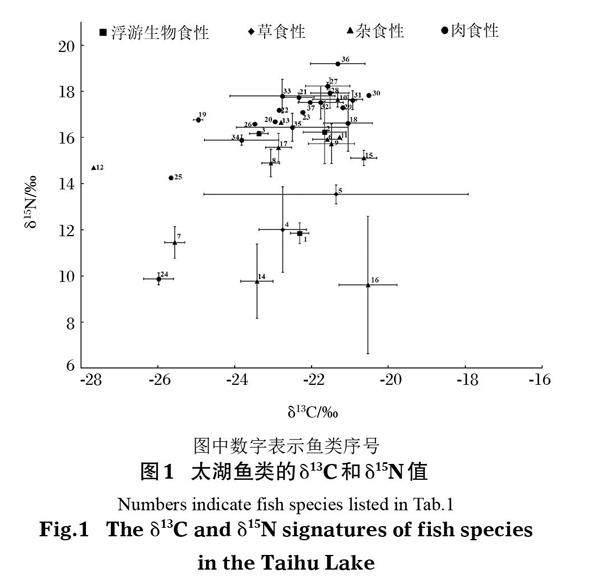
鱼类群落生态位大小的TA值为48.96‰2,SEAc值为12.43‰2。位于凸包顶点的鱼类有飘鱼、鲫、泥鳅、光泽黄颡鱼(Pelteobagrus nitidus)、黄鳝、红鳍原鲌和鮠一种(图2)。
各种类生态位大小的SEAc值在0.14‰2~20.43‰2(图3)。黄颡鱼(Pelteobagrus fulvidraco)SEAc值(20.43‰2)最大,其次依次是鳊(Parabramis pekinensis)(19.78‰2)、鲫(16.54‰2)、鳙(Aristichthys nobilis)(15.68‰2)、中国花鲈(Lateolabrax maculatus)(15.52‰2)、泥鳅(13.5‰2)等,光泽黄颡鱼SEAc值(0.14‰2)最小。
浮游生物食性、杂食性鱼类营养生态位重叠度均较小(<9.3%),生态位存在明显分离(图4)。在肉食性鱼类中,大银鱼(Protosalanx hyalocranius)与翘嘴鲌(Culter alburnus)的生态位重叠度最大(70.6%);黄鳝与其他肉食性鱼类的生态位完全分离,刀鲚(Coilia nasus)与沙塘鳢(Odontobutis obscurus)的生态位分离,光泽黄颡鱼仅与中国花鲈生态位有少量重叠(0.9%);其余肉食性鱼类间均存在一定程度的生态位重叠(0.4%~39.9%)。
太湖鱼类平均营养级范围为0.73~3.62(图5)。营养级最高的鱼类为鮠一种(3.62),最低的为泥鳅(0.73)。营养级大于3的鱼类有16种,营养级在2~3的有14种,小于2的有7种。泥鳅、黄颡鱼、鲫、鳊、鳙的营养级变化幅度较大。
3 讨论
3.1 太湖鱼类碳氮稳定同位素的变化特征
本研究对2019-2020年太湖鱼类的碳氮稳定同位素特征进行了分析,与以往研究相比(Mao et al,2012;Mao et al,2014;温周瑞等,2016),本研究涵盖了更多的鱼类物种。总体上看,太湖大部分鱼类的δ13C值分布范围较小(-24‰~-20‰),不同摄食类群间的差异不大,表明这些鱼类的能量来源较为接近,同时与部分鱼类生态位较大有关。鱼类δ15N值显示了较大的差异,这与不同种类的食性不同有关,也反映了鱼类在营养级上的差异(图5)。与以往研究数据相比,发现近20年部分鱼类如鲢(Hypophthalmichthys molitrix)、鳙、鲤(Cyprinus carpio)、鲫、刀鲚、红鳍原鲌、翘嘴鲌等的δ13C值有逐渐增大的趋势,δ15N值则逐渐下降。δ13C值指示了湖泊有机质来源情况,通常湖滨带及陆源有机质δ13C值较低,内源有机质δ13C值较高(吴汉等,2020)。鱼类δ13C值的增加表明湖滨带及陆源有机质对这些鱼类的能量贡献降低。鱼类δ15N值的降低主要与太湖水体总氮浓度降低有关(谷孝鸿等,2019)。
3.2 生态位大小与食物源及生境利用的关系
在稳定同位素生态学中,δ13C值可用于指示消费者的能量来源,δ15N值可用于指示营养位置,两者联合分析可较为准确地量化物种或群落的营养生态位空间(Peterson & Fry,1987;Jackson et al,2011)。本研究采用的标准差椭圆面积指标(SEAc)不仅反映了物种的食谱宽度,同时也反映了物种对不同生境的利用程度。例如,本研究发现黄颡鱼的SEAc值最大,表明其营养生态位空间较大,利用的食物源和生境范围较广。光泽黄颡鱼的SEAc值最小,表明其利用的生态位空间较小,利用的食物源和生境范围较窄。黄颡鱼、光泽黄颡鱼的生态位大小差异较大,主要与两者食性差异较大有关。研究表明,光泽黄颡鱼主要以水生昆虫幼虫和甲壳类为食(重量比为90.6%)(袁刚等,2011)。黄颡鱼个体较大,其食物颗粒也较大,主要以水生昆虫、虾类及软体动物为食(邹社校,1999;梁正其等,2019)。一般认为,鳊为草食性鱼类,其生态位空间应较小。本研究中鳊的SEAc值也较大,这主要与其食物源的稳定同位素差异较大有关。研究表明,太湖贡湖湾水生植物的δ13C值范围较大,为-29.1‰~-15.6‰,其中沉水植物δ13C值明显偏大,浮叶植物、漂浮植物和挺水植物则偏小(温周瑞等,2016),表明鳊在湖滨带和敞水区间存在跨生境利用。刀鲚、银鱼(icefish)、鲢、鳙是太湖重要的经济鱼类;本研究中,刀鲚和鲢的SEAc值均较小,表明其利用的食物源和生境范围较小,陈氏短吻银鱼(Salangichthys tangkahkeii)的SEAc值较小与其专食浮游动物有关,鳙的SEAc值较大,表明其利用的食物源和生境范围较广。总之,太湖鱼类营养生态位大小与摄食的食物源种类及生境均有关系。
3.3 生态位重叠与食物资源竞争的关系
本研究发现,同一摄食类群中既有营养生态位空间较大的物种,也存在生态位较小的物种,表明鱼类对同类型食物资源的利用方式上也存在分离,这与生态位重叠的分析结果一致(图4)。太湖大部分鱼类间的生态位重叠度较小,在生态位上均表现出明显的分离,这可能是太湖鱼类共存的一个重要原因。生态位重叠度在肉食性鱼类中较高,表明其食物资源竞争可能更激烈。在肉食性鱼类中,大银鱼与翘嘴鲌在营养生态位上的重叠度最大(70.6%),大银鱼主要摄食鱼苗而翘嘴鲌主要摄食幼鱼及成鱼(朱成德,1985;刘恩生,2008),两者在对资源利用的时间尺度上存在分离。因此,尽管两者在营养生态位上有较高重叠,但对食物资源的竞争可能并不强烈。翘嘴鲌与乌鳢(Channa argus)在营养生态位上的重叠度也较大(38.9%),表明两者存在一定程度的食物竞争,但两者栖息的水层有差异,翘嘴鲌多活动于中上层而乌鳢主要在水体底层觅食,两者在对资源利用的空间尺度上存在分离(倪勇和朱成德,2005)。刀鲚与其他肉食性鱼类的营养生态位重叠度较小,表明食物资源竞争较弱,这可能是刀鲚在太湖能长期维持高产的原因之一。鲢、鳙和陈氏短吻银鱼都是太湖重要的经济鱼类,这3种鱼类均摄食浮游生物,陈氏短吻银鱼与鲢、鳙在生态位上存在明显的分离,这与陈氏短吻银鱼专食浮游动物有关。研究表明,陈氏短吻银鱼属于视觉性摄食鱼类,摄食具有明显的选择性,主动捕食大型浮游动物,在大型浮游动物满足不了其需求时会转向摄食小型浮游动物(刘正文和朱松泉,1994)。鲢、鳙同为滤食性鱼类,但对饵料生物的选择能力较弱(谢平,2003)。有研究表明,在严重的富营养化情况下,太湖鲢、鳙摄食微囊藻的比例高达90%(刘恩生等,2007);同时,本研究发现以传统的摄食类群分类,无法体现物种在生态位空间上的差异。因此,在食物网研究中,基于物种的精细化研究是十分必要的。
3.4 太湖鱼类营养级及其与食物网的关系
营养级是生态位分析的一个重要内容,反映了物种在食物网中的地位和功能(王少鹏,2020)。本研究中,肉食性鱼类营养级最高,杂食性和草食性鱼类营养级较低,与以往研究结果基本一致(Mao et al,2012;李云凯和贡艺,2014;Li et al,2018)。鱼类中营养级最高的是鮠一种,其次是刀鲚、中国花鲈、长须黄颡鱼(Pelteobagrus eupogon)、大银鱼等,表明这些鱼类处于太湖鱼类食物网的顶级。泥鳅和黄鳝的营养级均较低,可能与其摄食较高比例的有机碎屑和底栖藻类有关(王姝然,2019)。本研究还发现泥鳅、鲫、鳊、鳙、黄颡鱼的营养级在个体间有较大差异,这主要与这些鱼类的营养生态位空间较大有关(图3),由此可见营养级变化受食物组成变化的影响较大。本研究中,太湖鱼类营养级最高为3.62,低于以往的研究(4.20)(Mao et al,2012)。营养级降低一定程度上反映了顶级捕食者(食鱼性鱼类)食物质量的降低,太湖鱼类面临小型化及渔业资源降低等问题(陈卫东等,2017),优质食物资源的缺乏可能导致部分食鱼性鱼类转食更多低营养级饵料生物;同时,营养级的降低可能导致太湖食物网出现扁平化趋势,即低营养级物种主导食物网结构与能量流动。
综上所述,分析“十年退捕”实施前太湖鱼类的营养生态位特征对太湖渔业资源的管理有一定借鉴意义。一方面,营养生态位分析中,位于凸包顶点的鱼类对于维系整个食物网稳定性具有十分重要的作用(Layman et al,2007),这些鱼类包括鮠一种、红鳍原鲌、黄鳝、泥鳅、光泽黄颡鱼、飘鱼和鲫,应在渔业资源管理中重点关注;另一方面,营养生态位空间分离是不同鱼类共存的一个重要前提,修复太湖生境多样性可为鱼类跨生境摄食提供条件,降低种间食物竞争。
志谢:感谢徐强强、梁磊、李颖、王锐在样品采集及王姝然、王康在样品处理和数据分析方面提供帮助。
参考文献
陈卫东,生楠,朱法明,2017. 太湖渔业资源现状及产业发展对策[J]. 安徽农业科学, 45(7):226-228.
谷孝鸿,曾庆飞,毛志刚,等,2019. 太湖2007-2016十年水环境演变及“以渔改水”策略探讨[J]. 湖泊科学, 31(2):305-318.
康乐,陈永林,1994. 草原蝗虫营养生态位的研究[J]. 昆虫学报, 37(2):178-189.
李云凯,贡艺,2014. 基于碳、氮稳定同位素技术的东太湖水生食物网结构[J]. 生态学杂志, 33(6):1534-1538.
李云凯,刘恩生,王辉,等,2014. 基于Ecopath 模型的太湖生态系统结构与功能分析[J]. 应用生态学报, 25(7):2033-2040.
梁正其,杨桂琴,杨秋菊,2019. 锦江河干流黄颡鱼的年龄、生长和资源利用研究[J]. 水产科学, 38(2):213-219.
刘恩生,鲍传和,曹萍,等,2007. 太湖鲢、鳙的食物组成及渔获量变化原因分析[J]. 水利渔业, 27(4):72-74.
刘恩生,2008. 太湖主要鱼类的食物组成[J]. 水产学报, 32(3):395-401.
刘正文,朱松泉,1994. 滇池产太湖新银鱼食性与摄食行为的初步研究[J]. 动物学报, 40(3):253-261.
倪勇,朱成德,2005. 太湖鱼类志[M]. 上海:上海科学技术出版社.
秦伯强,罗潋葱,2004. 太湖生态环境演化及其原因分析[J]. 第四纪研究, 24(5):561-568.
王少鹏,2020. 食物网结构与功能: 理论进展与展望[J]. 生物多样性, 28(11):1391-1404.
王姝然,2019. 富营养化对全湖食物网能流的影响-滇池和洱海的比较研究[D]. 武汉:中国科学院水生生物研究所.
温周瑞,熊鹰,徐军,等,2016. 太湖贡湖湾食物网特征研究[J]. 水生生物学报, 40(1):131-138.
吴汉,常凤琴,张虎才,等,2020. 异龙湖近百年来湖泊沉积物有机碳、氮稳定同位素变化及其环境指示意义[J]. 生态学杂志, 39(8):2478-2487.
谢平,2003. 鲢、鳙与藻类水华控制[M]. 北京:科学出版社.
徐德琳,2013. 人为干扰下的太湖食物网时间演变与区域分异[D]. 南京:南京大学.
徐军,王玉玉,王康,等,2020. 水域生态学中生物稳定同位素样品采集、处理与保存[J]. 水生生物学报, 44(5):989-997.
袁刚,茹辉军,刘学勤,2011. 洞庭湖光泽黄颡鱼食性研究[J]. 水生生物学报, 35(2):270-275.
张彤晴,唐晟凯,李大命,等,2016. 太湖鲢鳙放流增殖效果评价和容量研究[J]. 江苏农业科学, 44(9):243-247.
张翔,沈伟,周国栋,2021. 2018-2020年太湖鱼类群落结构及其环境因子典范对应分析[J]. 生态与农村环境学报, 37(5):674-680.
朱成德,1985. 太湖大银鱼生长与食性的初步研究[J]. 水产学报, 9(3):275-287.
朱春全,1997. 生态位态势理论与扩充假说[J]. 生态学报, 17(3):324-332.
邹社校,1999. 洪湖黄颡鱼的生长、食性和渔业地位[J]. 湖北农学院学报, 19(3):240-249.
Begon M, Townsend C R, Harper J L, 2006. Ecology: from individuals to ecosystems[M].4th edition. Oxford: Blackwell Publishing.
Hutchinson G E, 1957. Concluding remarks: Cold spring harbor symposium[J]. Quantitative Biology, 22:415-427.
Jackson A L, Inger R, Parnell A C, et al, 2011. Comparing isotopic niche widths among and within communities: SIBER-Stable Isotope Bayesian Ellipses in R[J]. Journal of Animal Ecology, 80(3):595-602.
Layman C A, Arrington D A, Montana C G, et al, 2007. Can stable isotope ratios provide for community-wide measures of trophic structure?[J]. Ecology, 88(1):42-48.
Li Y K, Zhang Y Y, Xu J, et al, 2018. Spatial variations in food web structures with alternative stable states: evidence from stable isotope analysis in a large eutrophic lake[J]. Journal of Oceanology and Limnology, 36(2):385-394.
Mao Z G, Gu X H, Zeng Q F, et al, 2012. Food web structure of a shallow eutrophic lake (Lake Taihu, China) assessed by stable isotope analysis[J]. Hydrobiologia, 683(1):173-183.
Mao Z G, Gu X H, Zeng Q F, et al, 2014. Seasonal and spatial variations of the food web structure in a shallow eutrophic lake assessed by stable isotope analysis[J]. Fisheries Science, 80(5):1045-1056.
Nithirojpakdee P, Beamish F W H, Boonphakdee T, 2014. Diet diversity among five co-existing fish species in a tropical river: Integration of dietary and stable isotope data[J]. Limnology, 15(1):99-107.
Peterson B J, Fry B, 1987. Stable isotopes in ecosystem studies[J]. Annual Review of Ecology and Systematics, 18:293-320.
Post D M, 2002. Using stable isotopes to estimate trophic position: models, methods and assumptions[J]. Ecology, 83(3):703-718.
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria[R/OL]. https://www.R-project.org/.
Rahman A U, Jones H P, Barber N A, et al, 2021. Disturbance-induced trophic niche shifts in ground beetles (coleoptera: carabidae) in restored grasslands[J]. Environmental Entomology, 50(5):1075-1087.
Taylor M D, Lowry M B, Becker A, 2018. Evaluating potential competitive interactions following stocking through trophic niche breadth[J]. Marine and Freshwater Research, 69(10):1614-1625.
Vasquez B R, Whitinger J A, Sitar S P, et al, 2021. Diet and trophic ecology of introduced salmonines at two south shore ports of Lake Superior, 2019[J]. Journal of Great Lakes Research, 47(4):1117-1125.
Wainright C A, Muhlfeld C C, Elser J J, et al, 2021. Species invasion progressively disrupts the trophic structure of native food webs[J]. Proceedings of the National Academy of Sciences of the United States of America, 118(45):e2102179118.
Wang S, Luo B K, Qin Y J, et al, 2021. Fish isotopic niches associated with environmental indicators and human disturbance along a disturbed large subtropical river in China[J]. Science of The Total Environment, 750:141667.
(责任编辑 万月华 熊美华)
Trophic Niche Analysis of Fish in Taihu Lake Using Stable Isotopes
of Carbon and Nitrogen
LIU Su‐qun1,2, SHEN Ming‐hua1,2, LIU Xue‐qin1
(1. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, P.R. China;
2. University of Chinese Academy of Sciences, Beijing 100049, P.R. China)
Abstract:In this study, we analyzed trophic niches across the fish community in the Taihu Lake using stable isotope signatures of carbon (δ13C) and nitrogen (δ15N) in fish species collected from the Taihu Lake in October-November of 2019 and September-November of 2020. Trophic niche size and niche overlap of different fish species were analyzed using SIBER (Stable Isotope Bayesian Ellipses in R) and the trophic position of fish was analyzed using double stable isotopic baselines. A total of 37 fish species were collected during the surveys, and the δ13C values ranged from -27.67‰ to -17.92‰, and δ15N values ranged from 6.02‰ to 20.31‰. Fish trophic niche sizes, quantified as the corrected standard ellipse area (SEAc), ranged from 0.14‰2 to 20.43‰2, with the largest niche size (20.43‰2) found in Pelteobagrus fulvidraco and the smallest niche size (0.14‰2) in Pelteobagrus nitidus. The SEAc of Coilia nasus, Salangichthys tangkahkeii and Hypophthalmichthys molitrix were small, indicating a narrow range of food sources and habitats, while the result of Aristichthys nobilis was the opposite. Niche overlaps among fish species varied from 0 to 70.6%. The largest overlap (70.6%) was between Protosalanx hyalocranius and Culter alburnus, followed by Lateolabrax maculatus and C. alburnus (39.9%), with niche overlaps among other fish species varying from 0 to 38.9%. Niche overlaps between C. nasus and other carnivorous fishes were low, indicating that competition for food resources was low. The trophic niche of S. tangkahkeii was clearly separated from H. molitrix and A. nobilis because S. tangkahkeii feeds exclusively on zooplankton. The trophic position of the fish community in the Taihu Lake ranged from 0.73 to 3.62, with Leiocassis sp. occupying the highest trophic level and Misgurnus anguillicaudatus the lowest. This study shows that the trophic niche size of different fish species is related to the type of food consumed and the habitat occupied. The niche overlaps among fish species were generally low in the Taihu Lake, indicating large separations in resource utilization among species. The study provides basic data for food web research on fish, fishery resource management and evaluation of ecological restoration in the Taihu Lake.
Key words:stable isotope; trophic niche; niche overlap; trophic position; fish species in Taihu Lake
收稿日期:2022-01-24 修回日期:2023-03-19
基金项目:国家重点研发计划项目(2018YFC0407200)。
作者简介:刘素群,1997年生,女,硕士研究生,研究方向为湖泊生态学。E-mail: liusuqun19@mails.ucas.ac.cn
通信作者:刘学勤,1979年生,男,副研究员,主要从事河流―泛滥平原生态学及水文生态学研究。E-mail: xqliu@ihb.ac.cn
