不同遮荫环境对小粒种咖啡光合作用的影响
2023-04-27萧自位白学慧苏琳琳张枝润马关润
萧自位,白学慧,肖 兵,苏琳琳,张枝润,马关润
(云南省德宏热带农业科学研究所,云南瑞丽 678600)
云南省是我国小粒种咖啡主要种植区,“云南咖啡”品牌在国内外市场逐渐受到欢迎,这对云南的咖啡产业发展具有良好的推动作用[1]。同时,云南发展咖啡产业具有较好的自然优势和经济效益[2]。小粒种咖啡为喜温凉、湿润、荫蔽环境的作物,在无荫蔽栽培条件下开花结果自控性差,这增大了管理咖啡的成本和难度[3-5]。适度荫蔽能增强咖啡开花结果的自控性,且有效地提高咖啡植株钾的含量,减少瘪果干果和枝条回枯现象的发生,使得咖啡高产稳产性增强,品质显著提高,生产成本明显降低[4-7]。
光合作用是植物生长发育的基础,增强作物的叶片光合作用能够有效提高作物的产量和品质。小粒种咖啡是一种喜阴植物,适当地遮荫能够有效提高咖啡树的光合作用[8-11]。虽然国内外许多学者对全光照和遮荫环境下的咖啡树光合作用进行了深入研究,揭示了遮荫不仅有利于提高咖啡树光合作用,而且对提高咖啡产量和品质也有积极作用[8-10]。然而,这些研究并未对遮荫环境条件下的各项环境要素进行量化分析,无法更精准地分析各种遮荫环境下的咖啡光合能力。为了精确地反映不同遮荫环境的差异,本研究引入了陈耀华所提出的遮荫效果概念[12],对两种遮荫环境的相关环境要素定量化,分析比较不同遮荫环境下咖啡树的光合作用能力,探索遮荫环境变化对咖啡树光合作用的影响,以期为生产实践提供科学依据。
1 材料和方法
1.1 研究区域概况
试验地位于云南省德宏热带农业科学研究所的咖啡优良品种种植基地内(24°1′12.28″N,97°51′21.36″E),该区属南亚热带季风气候,干湿两季分明,雨季降雨量约1 390 mm,占全年降雨量的80%以上,年均气温约21℃。
供试小粒种咖啡品种为德热390,属萨奇姆品系,2017 年定植,株行距为1 m×2 m。遮荫树种为龙眼树(Dimocarpus longan),株行距为4 m×12 m,为上世纪80 年代种植。试验地土壤容重约1.23 g/cm3。实验区每年施肥3 次,第一次在雨季来临时洒施复合肥,株施约100 g。第二次在8—9 月份施加尿素,株施约80 g。第三次在雨季结束前施加复合肥,株施约100 g。每年采收结束后对咖啡树进行适当修枝。
1.2 实验材料
在咖啡优良品种种植基地内选取两行具有不同遮荫程度的德热390 进行实验,一行位于两行龙眼树中间(遮荫I),另一行紧邻遮荫树(遮荫II)。每行选取生长健壮,且株型一致的植株5 株进行实验。每棵植株选取中上部同一方向生长良好的一分枝,以一分枝顶端以下的第三对或第四对完全成熟叶片作为测量材料,每株测量一次。
1.3 测定方法
1.3.1 光响应曲线测定
试验选择在植株生长旺盛的9—11 月进行。午后光照较强,遮荫对叶片的光合作用影响较大,因此选择天气晴朗的下午(15:00—17:30)对叶片进行光合测量,利用CIRAS-3(PP-Systems USA)便携式光合作用测定系统对选取的一分枝上的第3 对或第4 对成熟叶片进行光响应曲线测定。
大气相对湿度设置为50%~60%,叶温设置为 27℃,CO2浓度设置为 390 μmol·m-2·s-1,分别设置光合有效辐射(I)梯度设为 0、50、100、150、200、250、300、350、400、500、600、700、800、1 000、1 200 μmol·m-2·s-1,共 15 个梯度。每个光照强度等待120 s 后开始测定,每个梯度连续重复测定3次,每个遮荫环境每天测量1 株,毎株选择1 片叶片进行测量,每个遮荫条件下共测量5 株。
1.3.2 CO2响应曲线测定
CO2响应曲线和光响应曲线使用同一叶片测定,使用接近叶片饱和光强的光合有效辐射(I)为 700 μmol·m-2·s-1,CO2浓度梯度依次为 350、250、150、50、150、250、350、450、550、650、750、850、950、1 050、1 250、1 450、1 650 μmol·m-2·s-1,共17 个梯度。稳定时间和数据记录与测定光响应曲线时相同,每个遮荫环境每天测量1 株,毎株选择1 片叶片进行测量,每个遮荫条件下共测量5 株。
1.3.3 遮荫效果测定
测定完叶片光合作用后,参照陈耀华法进行遮荫效果测定[12],测定时间选择午后(15:30—16:30)天气晴朗的时候进行。在种植基地旁选择一块无遮荫树的裸地作为对照,在离地1.5 m 的高度运用CIRAS-3(PP-Systems USA)便携式光合作用测定系统自带环境光强测定功能测量3 个不同光照条件下的光照强度和温度,每个光照条件重复24 次,同时测树冠的高度。遮光度、降温率、荫质、遮荫效果计算公式如下[12]:
其中,I为全光照下的光强度,I′ 为不同遮荫条件下的光强度,t为全光照下的温度,t′为不同遮荫条件下的温度,S为遮荫面积,荫质单位为遮度,用N表示。
1.4 数据处理与分析
利用叶子飘直角双曲线修正模型能有效拟合小粒种咖啡的光响应曲线[13],本研究采用该模型进行光响应曲线拟合,模型公式如下。
式(3)中:ω是光响应曲线的初始斜率,β和γ为系数,I为光合有效辐射,Rd为暗呼吸速率。
饱和光强Isat为:
最大净光合速率用Pnmax为:
利用 Farquhar-von Caemmerer-BerryFvCB 生物化学光合模型(简称FvCB 模型)对A-Ci 曲线进行拟合,各限制阶段如下。
当Cc>(1+3α)Γ*时,
当Cc≤(1+3α)Γ*时,
其中:Cc表示叶绿体内 CO2分压;α表示光呼吸中未返回的乙醇酸所含C 的比例;Γ*表示叶绿体 CO2补偿点;A表示 CO2净吸收率;Wc表示Rubisco 限制下的羧化速率;Wj表示RuBP 再生限制下的羧化速率;Wp表示TPU 限制下的羧化速率;Rdlight表示光条件下吸速率。
其中:Vcmax为最大羧化速率;J为电子传递速率,在饱和光强下,设J=Jmax;TUP为磷酸丙糖从叶绿体运出的速率;Ci为胞间CO2分压,gm为叶肉 CO2导度;O为 O2分压;Kc为 CO2米氏常数,Ko为 O2米氏常数;Cc-trans为Rubicon 酶限制阶段与RuBP 限制阶段临界点的胞间CO2浓度。
根据 Grassi 和 Magnani(2005)的研究,在饱和光强下, CO2净吸收率(A)受气孔因素(ls)、叶肉导度因素(lmc)和生化因素(lb)限制,3 种限制因素计算公式如下[14]。
其中:gsc为气孔传导能力,可由气孔导度gs来计算(gsc=gs/1.6);gtot表示叶片对CO2的总导度,1/gtot=1/gsc+1/gm;∂A∂Cc是羧化位点叶绿体内 CO2浓度(Cc)为 50~100 μmol·mol-1时 A/Cc 曲线拟合线的 斜 率[15],根 据 Da Matta 的 方 法[11],。gm根据 Harley 的方法计算[16]。
利用叶子飘等的光合计算软件对光响应曲线进行拟合,得出相应的响应参数。A-Ci 曲线测量数据上传到https://www.leafweb.org/ 进行自动分析。根据Gu 提出的方法拟合生理参数和选择最优模型[17]。用 Microsoft Excel 对数据进行整理,采用SPSS 进行数据分析。
2 结果与分析
2.1 不同遮荫条件各环境参数比较
遮荫是影响小粒种咖啡生长的重要因素,两种处理的遮荫效果差异十分明显,遮荫Ⅱ的遮荫效果强于遮荫Ⅰ(表1),且遮荫Ⅱ遮光率和降温率也较高。
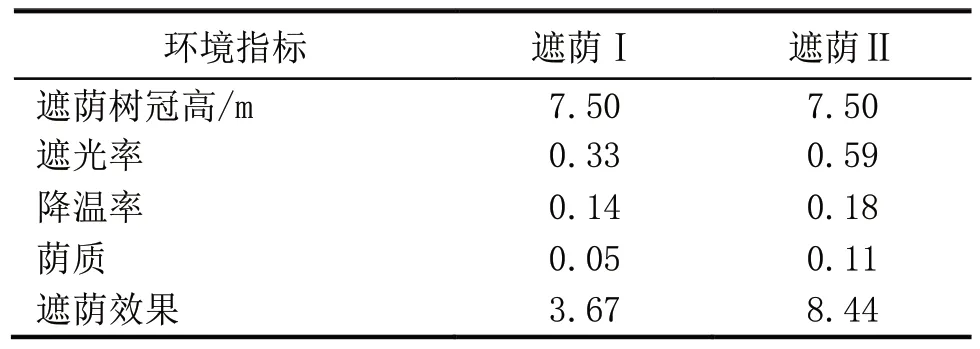
表1 两种遮荫环境的特征
2.2 不同遮荫环境下光合作用参数比较
两种遮荫环境下,小粒种咖啡光合作用参数如表2 所示,除ω和gm两个光合参数差异较小外,其它光合参数的差异都较为明显,但进行统计学比较时,二者均无显著差异(P>0.05)。两种遮荫环境小粒种咖啡光响应曲线的初始斜率(ω)相同,表明两种生长环境对咖啡叶片的初始量子效率影响较小。此外,遮荫效果较强时,最大净光合速率(Pnmax)、光补偿点(Ic)和暗呼吸速率(Rd)也较高,但光饱和点较低。
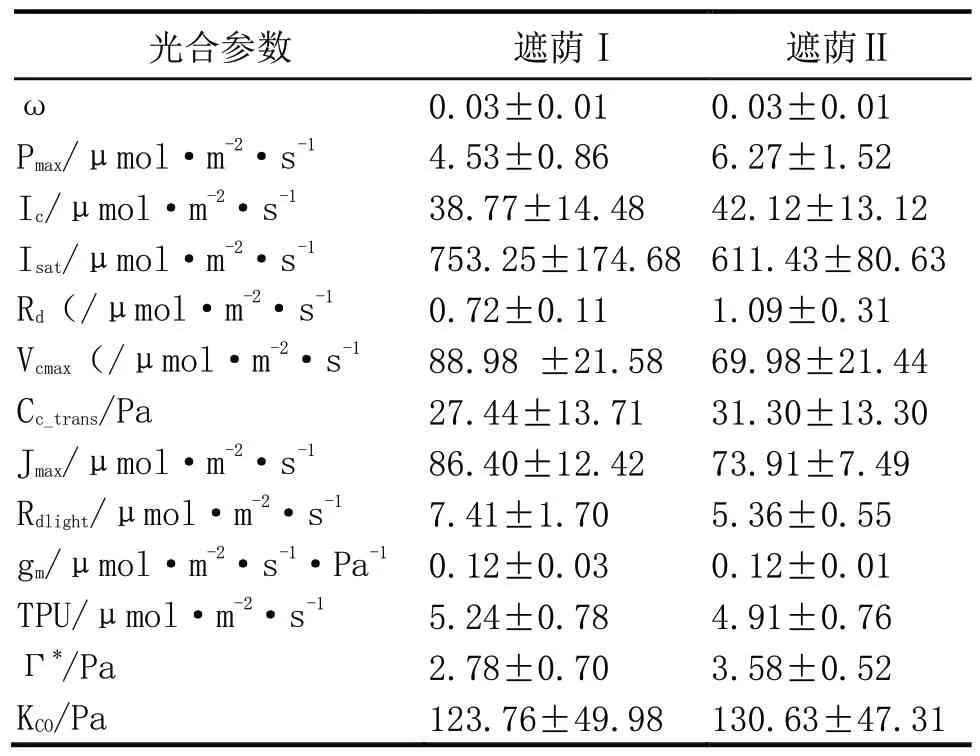
表2 两种遮荫环境下小粒种咖啡光合作用参数比较
运用FvCB 模型对两种环境下的咖啡叶片光合碳反应过程进行拟合,结果表明(表2),在遮荫效果相对低时,叶片的最大羧化速率(Vcmax)、最大电子传递速率(Jmax)、磷酸丙糖利用速率(TPU)和明呼吸速率(Rdlight)都相对较高,而叶绿体CO2光合补偿点(Γ*),Rubicon 酶限制阶段与 RuBP 限制阶段转换的胞间CO2浓度(Cc_trans)和KCO相对较低。
利用FvCB 模型拟和参数,对两种遮荫条件下小粒种咖啡叶片CO2响应曲线作图,结果如图1和图2 所示。遮荫效果较低时(遮荫Ⅰ),叶片的碳反应过程的各个阶段并不处于理想状态。随着胞间CO2浓度的增加,叶片中如果存在TPU 限制,CO2净吸收速率(A)先受核酮糖-1, 5-双磷酸羧化酶/加氧酶(Rubisco)的限制,然后受 TPU 限制。如果叶片中不存在TPU 限制,CO2净吸收速率(A)先受Rubisco 的限制,然后受核酮糖-1, 5-双磷酸(RuBP)再生限制。遮荫效果相对较高时(遮荫Ⅱ),随着胞间CO2浓度的增加, CO2净吸收速率(A)先受Rubisco 的限制,然后受RuBP 再生限制,如果叶绿体中CO2浓度继续增加,CO2净吸收速率(A)则很快受TPU 限制。
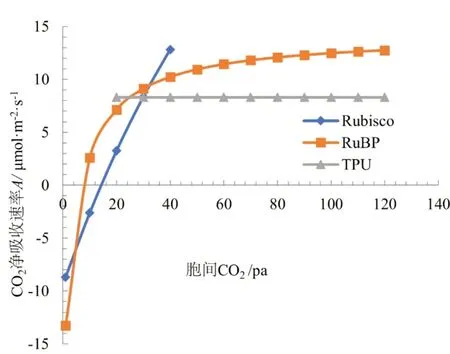
图1 遮荫I条件下小粒种咖啡叶片CO2响应曲线

图2 遮荫Ⅱ条件下小粒种咖啡叶片CO2响应曲线
2.3 光合限制因子定量分析
对不同遮荫条件下咖啡叶片光合限制因子进行定量分析(图3),结果表明,饱和光强下,两种遮荫条件下CO2净吸收速率(A)主要受气孔因素限制(ls),其次是叶肉导度因素(lmc),受生化因素(lb)的限制作用较小。虽然遮荫Ⅰ条件下气孔因素限制(ls)作用大于遮荫Ⅱ,但二者差异并不显著(P>0.05)。同样,两种不同遮荫条件的叶肉导度限制(lmc),生化限制(lb)差异也不显著(P>0.05)。
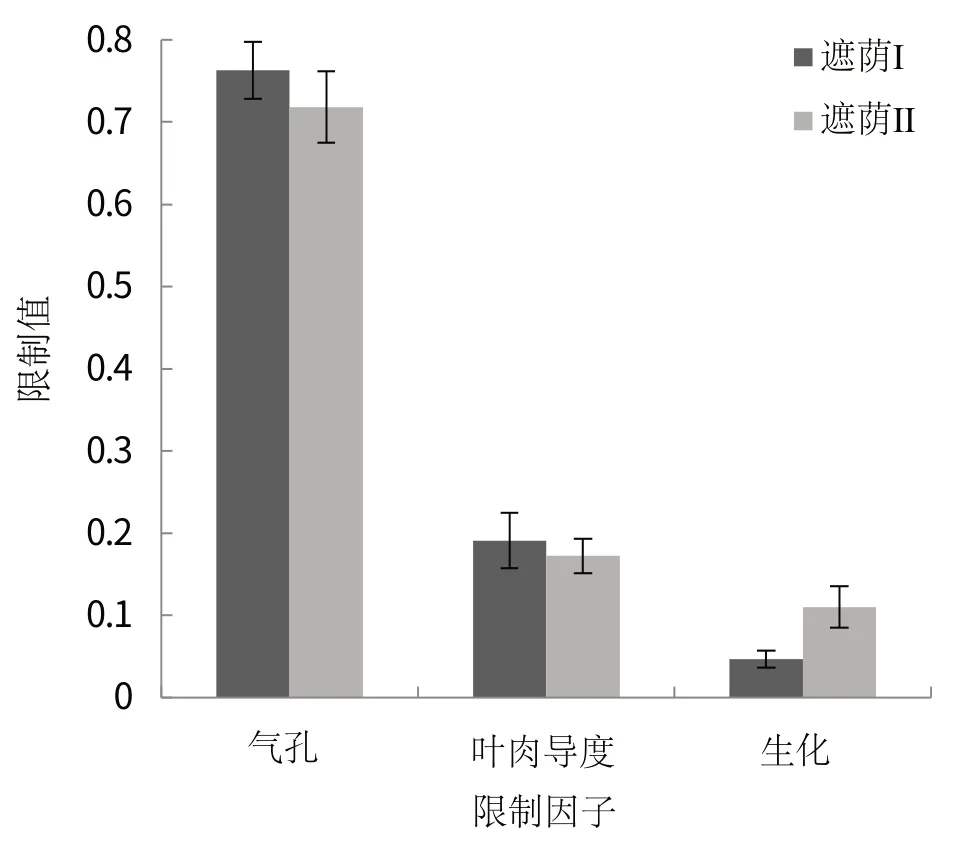
图3 不同遮荫条件下咖啡叶片光合限制因子定量分析
3 讨论与结论
陈耀华首次提出了关于荫质、遮荫效果等概念,运用定量计算公式量化遮荫树下相关的环境因子[12]。本文利用该方法量化了两种环境的遮荫效果,有效区分了咖啡树生长的两种不同环境。虽然两地不同遮荫环境下咖啡树光合作用参数无显著差异,但遮荫效果较强时,咖啡树光合作用能力也较高。该结果与Neto 等的研究并不一致,Neto 的研究发现紧邻遮荫树下咖啡树的光合能力明显低于两行遮荫树间的,但二者间无显著差异[18]。研究表明,咖啡树向阳面叶片能够承受较高光合有效辐射强度(如 2 000 μmol·m-2·s-1),但该条件下叶片的光合能力却很低[19],这可能因为咖啡树叶片受到了过强的太阳光辐射,叶片的光合能力受到抑制。同时,咖啡树叶片的饱和光强相对较低(300~700 μmol·m-2·s-1),在光线较强的条件下极易出现光抑制现象[18,20]。更高的遮荫效果能够有效降低太阳光辐射,在有遮荫树的环境下,咖啡叶片所接收的光合有效辐射强度降低,但叶片对光合有效辐射的利用率升高,使得遮荫条件下的咖啡树具有更高的光合能力[21]。
本研究中两种生长环境下咖啡叶片的初始量子效率相等,表明在光照条件很弱时,叶片对光合有效辐射的利用率相同。光呼吸速率与光合电子传递系统密切相关[22],并且随光照强度增加而增加[23]。在遮荫效果低的环境中,随着光照强度的增加,咖啡树的叶片会接收到更多的光合有效辐射,这可能是遮荫效果低的条件下,光呼吸速率(Rdlight)较高的原因。另外,在强光条件下,光呼吸速率的增加可能对叶片光合系统起到一定的保护作用[24-25]。因此,遮荫效果低的条件下具有较高的光呼吸速率(Rdlight)可能是叶片的一种自我保护作用,但这种保护机制或会降低叶片的净光合速率。已有研究表明,光照强度增加会提高咖啡叶片的光饱和点[10],这有效解释了遮荫效果低的咖啡树叶片光饱和点较高的结果。综上,适当提高遮荫效果可能有利于咖啡树的光合作用。
在生长季的强光条件下,咖啡树叶片CO2净吸收速率(A)主要受气孔因素限制(ls),这与已有的研究结果相一致[8-9,11]。因此,在生长季通过各种管理措施促使叶片气孔开放,降低气孔因素对咖啡叶片CO2净吸收速率(A)的限制,可能会起到有效增强光合速率的作用,进而增加有机物质积累,从而达到提高咖啡产量和品质的目的。
