树番茄幼苗叶片性状和生理参数对遮阴的响应及评价
2023-04-18李世民董琼金友帆李树萍李猛刘廷彪赵兴杰陈静叶平吕梦
李世民 董琼 金友帆 李树萍 李猛 刘廷彪 赵兴杰 陈静 叶平 吕梦


摘要:为探究树番茄幼苗在不同遮阴环境下叶片性状和生理响应机制,明确其幼苗光适应性及其生长的适宜光照环境,以树番茄幼苗为试验材料,研究正常光照(CK)、轻度遮阴(SL)、中度遮阴(SM)和重度遮阴(SH)4个处理对树番茄幼苗叶片性状、光合色素、非结构性碳水化合物(non-structural carbohydrates,NSC)及丙二醛(malonaldehyde,MDA)、脯氨酸(proline,Pro)、抗氧化酶等参数的影响,并通过主成分分析筛选树番茄耐阴指标,采用隶属函数对耐阴能力进行综合评价。结果表明,不同遮阴条件下树番茄幼苗各指标间均存在显著差异(P<0.05)。随着遮阴程度的加重,树番茄幼苗的叶长、叶面积及叶绿素a和类胡萝卜素含量等指标逐渐增大,叶绿素a/b及可溶性糖和淀粉含量、NSC均逐渐降低,而MDA、Pro、可溶性蛋白含量及抗氧化酶等指标呈先降低再升高的趋势,且在SM处理下最低。主成分分析表明,MDA、NSC、叶绿素总量、叶绿素a/b、叶鲜重、比叶面积、游离脯氨酸和SOD共8个指标可作为树番茄幼苗耐阴能力的鉴定指标。不同处理的综合评价排序依次为SM>SH>SL>CK。由此表明,树番茄幼苗期具有阴生特性,自然光照对树番茄幼苗存在强光胁迫;重度遮阴环境下存在明显的弱光胁迫。弱光胁迫下,树番茄幼苗主要通过增加单叶面积、叶绿素和类胡萝卜素含量,减小叶绿素a/b值,吸收更多的蓝紫光,以此来捕获更多的光能以提高光能利用率。此外,通过增加可溶性蛋白和Pro含量、提高抗氧化酶活性来缓解强光或弱光所产生的MDA毒害作用。综上,热区树番茄栽培、林下推广种植以及园林应用中可选择适当的半阴环境,遮阴强度50%左右较为适宜,为树番茄在热区栽培及林下推广种植提供了理论依据。
关键词:树番茄;叶片性状;生理参数;遮阴doi:10.13304/j.nykjdb.2021.0508
中图分类号:S641.9 文献标志码:A 文章编号:10080864(2023)01007211
光是植物进行光合作用必不可少的条件,是植物生长的能量来源,对植物的生长发育、生理生化反应、形态结构建成等方面有重要影响[1]。不同植物对光的需求不同,当植物长期处于过强或过弱的光环境下,可能遭受强光或弱光胁迫,其生长发育会受到抑制。强光环境下,植物的光合器官易受到光氧化损伤,使光合作用受到抑制;弱光环境下,植物净光合速率下降,合成的碳水化合物减少,植物体内的碳平衡被打破,从而造成植物碳饥饿[23]。当光照条件发生变化时,植物通过调整生长策略来适应环境变化。研究表明,随着光照强度的减弱,植物可能会通过增大单叶面积和比叶面积、减小比叶质量等来适应光环境的变化[4];提高可溶性蛋白、游离脯氨酸等渗透调节物质含量及提高过氧化物酶、过氧化氢酶等抗氧化酶活性来保护细胞膜不受损害[5];同时调节光合作用以提高光能利用率[6]。因此,研究叶片性状及生理参数的变化对深入了解植物对不同光照环境的适应性具有重要意义。
树番茄(Cyphomandra betacea Sendt.)属于茄科、树番茄属小乔木,有时为灌木,原产于南美洲,世界热带和亚热带地区均有引种,我国云南和西藏南部有栽培[7]。树番茄果实含有丰富的蛋白质、氨基酸、维生素C和钾、钙等矿物质元素及铁、硒等多种微量元素[8]。树番茄所含的花青素、酚类物质等生物活性成分已被证明具有抗氧化[9]、抗炎和抗癌作用[1011],对肝脏、扁桃体、喉咙痛及预防呼吸系统疾病和贫血有一定疗效[12]。此外,树番茄还可用于观花赏果,在庭园和街道种植能起到美化和绿化作用;也是农庄的优良树种,具有极高的经济价值[13]。目前,国内外学者对树番茄的研究多集中于种子育苗[14]、幼苗耐镉性[15]以及果实成分分析[1617]等方面。然而,关于不同遮阴环境下树番茄幼苗叶片性狀和生理生化响应特征的研究却鲜有报道。为此,本研究以树番茄幼苗为研究对象,设置不同遮阴环境,研究树番茄幼苗叶片性状及生理生化指标在不同处理下的变化,探寻其光适应性特征及适宜的光照条件,以期为树番茄幼苗在热区栽培、林下推广种植及园林应用中提供基础支撑和理论参考。
1 材料与方法
1.1 试验地概况
试验地位于云南省昆明市西南林业大学树木园(25°03′N,102°45′E),地处云贵高原中部。昆明属山原地貌,海拔为1 954 m,年平均气温16.5 ℃,年均降雨量700~1 100 mm,且降雨集中在6—9月,无霜期278 d,属于北亚热带低纬度半湿润高原山地季风性气候。土壤为偏酸性红壤,pH 5.45。
1.2 试验材料
于2020年7月中旬从云南省德宏州芒市树番茄分布区采集果实,带回实验室后剥出种子进行净种,风干后贮存于4 ℃;8月初进行穴盘育苗;10月初选取200株健康、无病害的幼苗移植于棕色育苗盆中。育苗盆规格:16.5 cm(直径)×14.5 cm(高),每盆1株。基质按腐殖质土和红壤体积比2:1充分混合消毒。
遮阴网:市场上常售的2针、3针和6针黑色遮阴网,具体遮光率由广州速为手持亮度检测仪SW-6023实测,遮阴程度分别为25.0%、47.2%、70.0%。
1.3 试验设计
试验于2021年3月初在西南林业大学后山树木园进行,采用单因素顶棚式设计,将幼苗随机分成4组,分别进行无遮荫(CK:自然光照,无遮阴网)、轻度遮荫(SL:75.0%自然光照,2针遮阴网)、中度遮荫(SM:52.8%自然光照,3针遮阴网)及重度遮荫(SH:30%自然光照,6针遮阴网)处理,每个处理10株幼苗,随机排列,3次重复,共120株幼苗。幼苗株高(28.12±2.17) cm,地径(7.13±1.42) mm,株行距均为40 cm。试验期间进行统一水肥管理,每周随机调换盆的位置,以保证各处理内的光照条件基本一致。于6月初测定树番茄幼苗的叶片性状和生理生化指标。
1.4 测定项目及方法
1.4.1 叶片性状 于2021年6月初,每个处理随机选取4株苗木,每株苗木上选择正常、无病害的功能叶片用于测定叶片参数。采用手持叶面积仪准确扫描和测量叶长(leaf length,LL)、叶宽(leafwide,LW)和单叶面积(single leaf area,LA);用万分之一天平称量扫描叶片的鲜重(leaf freshweight,LFW);然后将叶片放入烘箱,105 ℃杀青30 min,最后80 ℃烘干至恒重,称量干重(leaf dryweight,LDW)。根据以下公式计算叶片含水率(leaf moisture content,LMC)、叶形指数(leaf shapeindex,LSI)、比叶面积(specific leaf area,SLA)和比叶质量(leaf mass per unit area,LMA)。
叶片含水率=(鲜重-干重)/鲜重(1)
叶形指数=叶长/叶宽(2)
比叶面积(cm2·g-1)=叶片面积/叶片干重(3)
比叶质量(g·cm-2)=叶片干重/叶片面积(4)
1.4.2 生理生化指标 于2021年6月初,每个处理随机选取4株苗木,每株苗木选择正常、无病害的功能叶片,用液氮冷冻后置于-80 ℃冰箱中保存,用于生理指标测定。采用紫外分光光度计法测定叶绿素a(chlorophyll,Chl a)、叶绿素b(chlorophyll,Chl b)、叶绿素a+b(chlorophyll,Chla+b)和类胡萝卜素(carotenoid,Car)含量,并计算叶绿素a/b(Chl a/b)和類胡萝卜素/叶绿素(Car/Chl);采用硫代巴比妥酸法测定丙二醛(malondialdehyde,MDA)含量;采用磺基水杨酸法测定游离脯氨酸(proline,Pro)含量;采用滴定法测定过氧化氢酶(catalase,CAT)活性;采用愈创木酚法测定过氧化物酶(peroxisome,POD)活性;采用氮蓝四唑法测定超氧化物歧化酶(superoxidedismutase,SOD)活性;采用苯酚比色法测定可溶性糖(soluble sugar,SS)和淀粉(starch)含量;采用G-250 比色法测定可溶性蛋白(soluble protein,SP)含量;采用酶解法测定非结构性碳水化合物(non-structural carbohydrates,NSC)含量[18]。各生理指标均设置3次重复。
1.5 数据处理
采用Excel 2016 进行数据的整理和分析,Oringin 8.0绘图;利用 SPSS 26.0进行单因素方差分析和LSD 多重比较。并进一步进行主成分分析,采用模糊数学的隶属函数法[19]计算不同遮阴处理下的隶属函数值U(Xi ),综合评价树番茄幼苗的光适应性。隶属函数计算公式如下。
U(Xi)=(Xi-Xmin)/(Xmax-Xmin) (5)
式中,Xi为第i 个指标测定值;Xmax和Xmin分别为该指标的最大值和最小值。反隶属函数用1-U(Xi)计算。
2 结果与分析
2.1 遮阴对树番茄幼苗叶片性状的影响
如表1所示,不同遮阴处理下树番茄叶片的叶长、叶宽、单叶面积、比叶面积和比叶质量等指标间均存在显著差异。与CK相比,SL处理树番茄幼苗的叶片各参数无显著变化;SM处理的比叶质量显著降低11.7%,而叶鲜重、叶干重和单叶面积分别显著增加48.20%、33.54%和29.19%;SH处理树番茄幼苗的比叶质量显著降低16.70%,而叶长、叶宽、叶鲜重、叶干重、单叶面积和比叶面积分别显著增加19.69%、26.80%、55.85%、32.25%、46.99% 和16.52%。综上所述,SH处理树番茄幼苗的变化最显著。
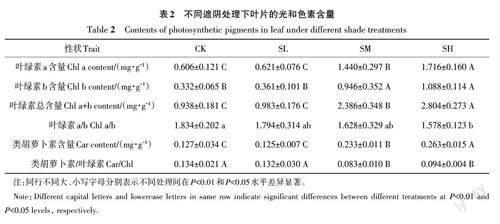
2.2 遮阴对树番茄幼苗叶片光合色素的影响
从表2可知,树番茄幼苗的类胡萝卜素、叶绿素 a、叶绿素 b和叶绿素总含量及叶绿素a/b值、类胡萝卜素/叶绿素值在不同处理间存在差异。随着光照强度的逐渐减弱,幼苗叶片的叶绿素 a、叶绿素 b、叶绿素a+b及类胡萝卜素含量均逐渐增加;而叶绿素a/b 值逐渐降低;类胡萝卜素/叶绿素值先降低再小幅上升。与CK相比,SL处理树番茄幼苗叶片光合色素含量无显著变化;而SM和SH处理树番茄幼苗的光合色素含量均显著增加,其中,SH处理最高。
2.3 遮阴对树番茄幼苗叶片非结构性碳水化合物的影响
如图1所示,不同处理间树番茄幼苗叶片的可溶性糖、淀粉和非结构性碳水化合物(NSC)含量存在显著差异。随着遮阴程度的加重,叶片可溶性糖、淀粉和NSC含量呈逐渐下降趋势。与CK相比,SL处理幼苗叶片的可溶性糖、淀粉和NSC含量分别降低13.78%、2.50%和14.45%,但差异不显著;SM处理幼苗叶片的可溶性糖、淀粉和NSC含量分别显著降低40.59%、33.73%和41.29%;SH处理幼苗叶片的可溶性糖、淀粉和NSC含量分别显著降低54.25%、48.75%和54.71%。
2.4 遮阴对树番茄幼苗丙二醛和渗透物质的影响
如图2所示,不同处理树番茄幼苗的丙二醛(MDA)、可溶性蛋白(SP)和游离脯氨酸(Pro)含量存在显著差异。随着遮阴程度的加深,树番茄幼苗叶片的MDA、Pro 和SP 含量先降低后升高,在SM处理达到最小值。与CK相比,SM和SH处理幼苗叶片的MDA含量显著降低;SM处理幼苗叶片的SP含量显著降低;遮阴处理幼苗叶片的Pro含量极显著降低,其中,SM处理最低。 SM处理的MDA含量较CK显著降低19.80%,SP和Pro含量较CK和SH处理分别显著降低21.43%、30.42%和35.04%、15.94%。
2.5 遮阴对树番茄幼苗酶促保护系统的影响
由图3可知,树番茄幼苗叶片的SOD、POD和CAT 活性随光照强度的减弱先降低后升高,均在在SM处理时达到最小值。SM处理下,幼苗叶片的SOD活性较CK处理极显著降低43.00%,较SH处理极显著降低58.09%;POD活性较 CK处理显著降低27.79%,与SH处理差异不显著;CAT活性较CK和SH处理分别显著降低38.49%和40.99%。由此表明,过强或过弱的光照都会使树番茄幼苗SOD、POD 和CAT 活性提高,以缓解胁迫造成的伤害,SM处理更有利于缓解光照胁迫。
2.6 树番茄幼苗光适应性的综合评价
2.6.1 主成分分析 经主成分分析,共提取出3个主成分因子,其累计贡献率达99.09%(表3)。第1主成分的贡献率为43.97%,其主要影响因子包括叶片含水率、MDA、淀粉、可溶性糖、NSC、叶绿素a含量、叶绿素b含量、叶绿素a/b、总叶绿素含量、类胡萝卜素和类胡萝卜素/叶绿素11个指标,主要反映了遮阴处理对叶绿素和光合产物的影响;第2主成分的贡献率为30.97%,主要作用因子包括叶鲜重、叶干重、叶长、叶宽、叶形指数、叶面积、比叶面积和比叶质量8个指标,主要反映了遮阴处理对叶片生长指标的影响;第3主成分的贡献率为24.15%,主要作用因子包括游离脯氨酸、CAT、POD、SOD和可溶性蛋白5个指标,主要反映了遮阴处理对抗氧化酶系统和渗透调节物质的影响。综上所述,树番茄幼苗叶片的MDA、淀粉、SS、NSC、叶绿素a含量、叶绿素b含量、叶绿素a/b、叶绿素总量、类胡萝卜素、叶鲜重、叶干重、叶长、叶面积、比叶面积、POD 等指标的荷载量较大,可作为树番茄耐阴性的鉴定指标。
对3个主成分的各指标间进行相关分析,删减相关性较高的重复性指标[20],结果表明,叶绿素总量、叶绿素a/b、MDA和NSC作为第1主成分的代表性指标(表4);叶鲜重和比叶面积作为第2主成分的代表性指标(表5);Pro和SOD作为第3主成分的代表性指标(表6)。因此,选择MDA、NSC、叶绿素总量、叶绿素a/b、叶鲜重、比叶面积、Pro含量和SOD活性共8个指标进行树番茄幼苗在不同遮阴处理下的表现进行综合评价。
2.6.2 隶属函数综合分析 依据隶属均值可划分植物抗逆性为4 个等级[21],隶属值0.6~1 为高抗型,0.4~0.6为中抗型,0.2~0.4为低抗型,小于0.2为不抗型。利用筛选出来的8个指标计算树番茄在不同遮阴处理下的隶属函数值,结果(表7)表明,树番茄幼苗平均隶属值为0.485,即树番茄幼苗的光胁迫抗性为中抗型。对不同处理树番茄幼苗的隶属函数均值進行排序,表现为SM>SH>SL>CK,表明SM处理最适宜树番茄幼苗的生长,强光或弱光都会对其造成胁迫,影响树番茄幼苗的生长发育。
3 讨论
3.1 树番茄幼苗叶片性状对遮阴的响应
植物叶片性状与植物的生长对策和植物利用资源的能力密切相关,且直接影响到植物的基本行为和功能[22]。叶片结构是植物叶片理化特征的基础,能够客观展现植物对外部环境的适应性[23]。本研究结果显示,树番茄叶片的叶长、叶宽、叶面积、比叶面积和比叶质量等指标在不同遮阴环境下存在显著差异。其中,叶形指数和比叶质量显著降低,叶长、叶宽、叶鲜重、叶干重、叶面积和比叶面积显著增加。此外,与CK相比,轻度遮阴处理下树番茄幼苗的叶片各参数变化较小;中度和重度遮阴环境下树番茄幼苗的叶形指数和比叶质量减小,叶长、叶宽、叶鲜重、叶干重、单叶面积和比叶面积增大,说明树番茄幼苗通过调整叶片形状和增大叶面积来减小光辐射距离,同时增强叶片对弱光的捕获能力,从而提高光能利用效率。
CK处理下,树番茄幼苗叶片有向内卷曲的趋势,而遮阴处理后叶片呈舒张状态,推测可能是树番茄幼苗对强光的一种避光机制,适当的遮阴有利于保护叶片免受强光伤害。中度和重度遮阴环境下,树番茄幼苗的叶面积、比叶面积和比叶质量变化较大,说明树番茄幼苗主要通过增大叶面积及增加干物质积累来响应光环境的变化,与马天光等[24]研究结果一致。
3.2 树番茄幼苗在不同遮阴环境下的生理调控机制
光合色素是植物进行光合作用的重要基础物质,其含量在一定程度上反映了植物光合能力的高低[25]。本研究发现,树番茄幼苗叶片的叶绿素a、叶绿素b和类胡萝卜素含量及叶绿素a/b在不同处理间差异显著。其中,叶绿素a、叶绿素b、叶绿素总量及类胡萝卜素含量均随着遮阴强度的增加而升高,说明树番茄幼苗通过增加色素含量来捕获更多的光能。叶绿素a/b是衡量植物耐阴性的重要指标[26]。通常情况下,植物为适应低光环境会降低叶绿素a/b。Bertamini等[27]研究发现,重度遮阴下植株叶绿素a/b值降低,增强对蓝紫光的利用,从而适应阴暗环境。本研究也发现,随着遮阴强度的增加,叶绿素a/b逐渐降低,由此表明,叶绿素a/b值降低可能是树番茄幼苗对低光环境的一种适应机制。类胡萝卜素/叶绿素可作为光伤害敏感指数[28],其比值越高,植物的光合作用越低。本研究表明,随着遮阴程度的加深,类胡萝卜素/叶绿素值先降低后升高,在中度遮阴处理达到最小值,说明过强或过弱的光照都会对树番茄幼苗造成光胁迫。
非结构性碳水化合物(NSC)主要指糖类和淀粉,是参与植株生命代谢的重要物质[29],也是光合作用的主要产物。在弱光环境下,植物光合作用所需要的光强条件不足,合成的碳水化合物减少。本研究表明,随着遮阴程度的加深,树番茄幼苗叶片单位质量内的可溶性糖、淀粉和NSC含量逐渐减少,且正常光照下叶片单位质量的NSC含量显著高于中度和重度遮阴处理,但中度和重度遮阴处理下的叶片干重分别是正常光照处理的1.5和1.48倍,说明弱光环境下树番茄幼苗叶片的NSC可能主要用于叶片的生长发育,从而提高叶片干物质的积累,与黄河腾等[30]对木奶果的研究结果略有不同,可能是2种植物的耐阴性差异造成。
膜脂过氧化程度可用MDA含量来衡量[31],而脯氨酸(Pro)和可溶性蛋白作为渗透调节物质其含量的增加通常可以缓解植物在胁迫中受到的损伤[32]。本研究表明,树番茄幼苗叶片中MDA、Pro和SP含量均随着遮阴程度的加深先降低后升高,其中,SM处理的MDA、Pro和SP含量最低,表明过强或过弱的光照都会使幼苗叶片细胞膜受到损害,适度的遮阴可以对细胞膜起到保护作用,与郭品湘等[6]研究结果一致。
SOD、POD和CAT是抗氧化酶保护系统中的3种酶,其中SOD处于核心地位,能有效地清除自由基,高SOD、POD 和CAT 活性可以提高植物抗逆性[33]。本研究结果表明,树番茄幼苗叶片SOD、POD 和CAT活性随着遮阴程度的加量先降低后升高,其中,在中度遮阴处理时达到最小值;SOD和CAT活性在重度遮阴处理时达到最大值,说明过强或过弱的光照都会激活树番茄幼苗的SOD、POD和CAT活性以缓解胁迫对植物造成的损伤,而中度遮阴处理下抗氧化酶活性最低,推测该处理下的光照强度更有利于树番茄幼苗的生长发育。
3.3 树番茄幼苗光适应性综合评价
通过对树番茄幼苗叶片生长指标和理化指标等24个指标进行主成分分析,筛选出8个指标用于综合评价,结果表明,中度遮阴处理最适宜树番茄幼苗的生长,强光或弱光都会限制其生长发育。且树番茄幼苗平均隶属值为0.485,表明树番茄幼苗比较耐阴,因此,今后树番茄在热区栽培、林下推广种植以及园林应用中应选择适当的半阴环境,以50%遮阴强度较为适宜。
参考文献
[1] 李小琴,张凤良,杨湉,等.遮阴对濒危植物风吹楠幼苗叶形
态和光合参数的影[J].植生理学报,2019,55(1):80-90.
LI X Q, ZHANG F L, YANG T, et al .. Effect of shading on leaf
morphology and photosynthetic parameters in endangered
Horsfieldia glabra seedlings [J]. Plant Physiol. J., 2019, 55(1):
80-90.
[2] CLELAND R E, MELIS A, NEALE P J. Mechanism of
photoinhibition: photochemical reaction center inactivation in
system II of chloroplasts [J]. PHS Res., 1986, 9(1-2):79-88.
[3] 何欣,張攀伟,丁传雨,等.弱光下硝铵比对小白菜氮吸收和
碳氮分配的影响[J].土壤学报, 2009, 46 (3):452-458.
HE X, ZHANG P W, DING C Y, et al .. Effects of nitrate/
ammonium ratio on nitrate absorption and distribution of
carbon and nitrogen in pakchoi growing under low light
intensity [J]. Acta Pedol. Sin., 2009, 46(3):452-458.
[4] DAI Y, SHEN Z, LIU Y, et al .. Effects of shade treatments on
the photosynthetic capacity, chlorophyll fluorescence, and
chlorophyll content of Tetrastigma hemsleyanum Diels et
Gilg [J]. Environ. Experim. Bot., 2009, 65(2-3):177-182.
[5] JALEEL C A, RIADH K, GOPI R, et al .. Antioxidant defense
responses: physiological plasticity in higher plants under
abiotic constraints [J]. Acta Physiol. Plantarum., 2009, 31(3):
427-436.
[6] 郭品湘,尹婷,粟春青,等.遮阴对双色木番茄幼苗生理特性
的影响[J].森林与环境学报, 2020, 40(1):76-82.
GUO P X, YIN T, SU C Q, et al .. Effects of shade on the
physiological characteristics of Solanum wrightii seedlings [J].
J. For. Environ., 2020, 40(1):76-82.
[7] 中国科学院中国植物编辑委员会.中国植物志[M].北京:科
学出版社,1978:141.
[8] 张东华,汪庆平.具有开发前景的热带果蔬植物——树番茄
[J].资源开发与市场,1999,14(5): 209-210.
[9] KOU M C, YEN J H, HONG J T, et al .. Cyphomandra Betacea
Sendt. phenolics protect ldl from oxidation and PC12 cells
from oxidative stress [J]. LWT Food Sci. Technol. 2009, 42(2):
458-463.
[10] JOSHI U J, GADGE A S, DMELLO P, et al .. Antiinflammatory,
antioxidant and anticancer activity of quercetin
and its analogues [J]. Int. J. Res. Pharma Biomed. Sci., 2011, 2:
1756-1766.
[11] MUTALIB M A, RAHMAT A, ALI F, et al .. Nutritional
compositions and antiproliferative activities of different solvent
fractions from ethanol extract of Cyphomandra betacea
(Tamarillo) fruit [J]. Malays. J. Med. Sci., 2017, 24(5):19-32.
[12] LIM T K. Solanum Betaceam. In: Edible Medicinal and Non-
Medicinal Plants [M]. Dordrecht: Springer, 2013:326-332.
[13] 郭碧瑜,周偉华,叶青莲,等.树番茄的生物学特性及栽培技
术[J].广东农业科学,2007, 34(12):102-103.
GUO B Y, ZHOU W H, YE Q L, et al .. Biological characteristics
and cultivation techniques of Cyphomandra betacea [J].
Guangdong Agric. Sci., 2007, 34(12):102-103.
[14] 董琼,何祯,徐云鹏,等.不同基质对树番茄穴盘育苗效果的
影响[J].种子,2012, 31(5):50-53.
DONG Q, HE Z, XU Y P, et al .. Effect of different basal
fertilizers on Cyphomandra betacea Plug seeding breeding [J].
Seed, 2012, 31(5):50-53.
[15] 黄科文,李嘉宇,李祉钰,等.喷施脱落酸对树番茄幼苗生理
特性及镉吸收的影响[J]. 土壤通报, 2020,51(1):207-213.
HUANG K W, LI J Y, LI Z Y, et al .. Effects of spraying
abscisic acid on physiological characteristics and cadmium
uptake of Cyphomandra Beracea seedlings [J]. J. Soil Sci.,
2020, 51(1):207-213.
[16] 董琼,李世民,高尚杰,等.不同种源树番茄果实品质比较及
综合分析[J].食品与发酵工业,2022,48(4):266-273.
DONG Q, LI S M, GAO S J, et al .. Comparison and
comprehensive analysis of fruit quality of Cyphomandra
betacea from different provenance [J]. Food Ferment. Ind.,
2022,48(4):266-273.
[17] CHEN X, FEDRIZZI B, KILMARTIN P A, et al .. Free and
glycosidic volatiles in tamarillo (Solanum betaceum Cav. syn.
SendtCyphomandra betacea.) juices prepared from three cultivars
grown in New Zealand [J/OL]. J. Agric. Food Chem., 2021,69(15):
837[2021-03-10]. https://doi.org/10.1021/acs.jafc.1c00837.
[18] 路文静,李奕松.植物生理学实验教程[M].北京:中国林业出
版社,2011:130-132.
LU W J, LI Y S. Experimental Course of Plant Physiology [M].
Beijing: China Forestry Association, 2011:130-132.
[19] 黄溦溦,张念念,胡庭兴,等高温胁迫对不同种源希蒙得木叶
片生理特性的影响[J].生态学报, 2011,31(23):62-70.
HUANG W W, ZHANG N N, HU T X, et al .. Effects of hightemperature
stress on physiological characteristics of leaves of
Simmondsia Chinensis seedlings from different provenances [J].
Acta Ecol. Sin., 2011, 31(23):7047-7055.
[20] 张辉,赵秋红.基于主成分分析基本原理的经济指标的筛选
方法[J].山东财政学院学报, 2013(2):52-61.
ZHANG H, ZHAO Q H. An economic indicator screening method
based on fundamental principle of principal components analysis
[J]. J. Shandong Univ. Fin. Econ., 2013(2):52-61.
[21] 刘卓.不同苜蓿品种耐盐性、抗旱性比较的研究[D].长春:
吉林农业大学,2008.
LIU Z. Study and appraisal of salt tolerance drought resistance
capacity of different alfalfa cultivars [D]. Changchun: Jilin
Agricultural University, 2008.
[22] REICH P B, BUSCHENA C T, JOELKER M G, et al .. Variation
in growth rate and ecophysiology among 34 grassland and savanna
species under contrasting N supply: a test of functional group
differences [J]. New Phytol., 2010, 157(3):617-631.
[23] ACKERLY D, KNIGHT C, WEISS S, et al .. Leaf size, specific
leaf area and microhabitat distribution of chaparral woody
plants: contrasting patterns in species level and community
level analyses [J]. Oecologia, 2002, 130(3):449-457.
[24] 马天光,李向义,林丽莎,等.遮阴对骆驼刺叶性状和水分生
理的影响[J].生态学报,2018,38(23):8466-8474.
MA T G, LI X Y, LIN L S, et al .. The effects of shade on leaf
traits and water physiological characteristics in Alhagi
sparsifolia [J]. Acta Ecol. Sin., 2018, 38(23):8466-8474.
[25] 张斌斌,姜卫兵,翁忙玲,等.遮阴对红叶桃叶片光合生理的
影响[J].园艺学报,2010,37(8):1287-1294.
ZHANG B B, JIANG W B, WENG M L, et al .. Effects of
shading on photosynthetic characteristics of red-leaf peach [J].
Acta Hortic. Sin., 2010, 37(8):1287-1294.
[26] 劉忆文,董彦娜,白靖怡,等.加拿大美女樱的耐荫性[J].江苏
农业学报,2017,33(6):1438-1440.
LIU Y W, DONG Y N, BAI J Y, et al .. Study on the shade
tolerance of Verbenr hybrida [J]. Jiangsu J. Agric. Sci., 2017, 33
(6):1438-1440.
[27] BERTAMINI M, MUTHUCHELIAN K, NEDUNCHEZHIAN
N. Shade effect alters leaf pigments and photosynthetic
responses in Norway spruce (Picea abies L.) grown under field
conditions [J]. Photosynthetica, 2006, 44(2):227-234.
[28] WILLEKENS H, CAMP W V, MONTAGU M V, et al .. Ozone,
sulfur dioxide, and ultraviolet b have similar effects on mRNA
accumulation of antioxidant genes in Nicotiana plumbaginifolia
L. [J]. Plant Physiol., 1994, 106(3):1007-1014.
[29] 潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物
的生理生态学研究进展[J].植物学通报,2002,19(1):30-38.
PANG Q M, HAN X G, BAI Y F, et al .. Advances in
physiology and ecology studies on stored non-structure
carbohydrates in plants [J]. Bull. Bot., 2002,19(1):30-38.
[30] 黄河腾,黄剑坚,陈杰,等.不同遮阴环境下木奶果幼苗生长
与生理生化的响应[J].生态学杂志, 2020,39(5):1538-1547.
HUANG H T, HUANH J J, CHEN J, et al .. Growth,
physiological and biochemical response of Baccaurea ramiflora
Lour. seedlings to different shading environments [J]. Chin. J.
Ecol., 2020, 39(5):1538-1547.
[31] 赵子豪,宋琦,李利,等.南方四季杨雌雄幼苗对镉胁迫光合
生理响应的差异[J].森林与环境学报,2019,39(2):201-207.
ZHAO Z H, SONG Q, LI L, et al .. Difference in photosynthesis
and physiological response of male and female Populus
deltoides nigrato Cd stress [J]. J. For. Environ., 2019, 39(2):
201-207.
[32] 唐钢梁,李向义,林丽莎,等.骆驼刺在不同遮阴下的水分状
况变化及其生理响应[J].植物生態学报,2013,37(4):354-364.
TANG G L, LI X Y, LIN L S, et al .. Change of different shading
on moisture conditions and the physiological response in
Alhagi sparsifolia [J]. J. Plant Ecol., 2013, 37(4):354-364.
[33] 苍晶,李唯.植物生理学[M].北京:高等教育出版社,2017:356.
CANG J, LI W. Plant Physiology [M]. Beijing: Higher
Education Press, 2017:356.
(责任编辑:张冬玲)
