秋华柳种群光合作用性能对人工调节水位消涨的响应
2023-04-18沈承叙陈芳清黄永文
沈承叙,陈芳清,吴 宇,吕 坤,黄永文
(1.湖北省三峡大学三峡地区生态保护与治理国际联合研究中心,湖北 宜昌 443002;2.湖北省正江环保科技有限公司,湖北 宜昌 443002)
【研究意义】秋华柳(SalixvariegataFranch)是杨柳科柳属多年生灌木,具有较好的耐淹与抗旱能力,是一种优良的固土护岸植物[1]。主要分布于我国西藏东部、云南北部、贵州、四川、甘肃东南部等地,生长于河岸带的沙土、卵石地和石缝间[2]。随着河流的水位消涨,河岸带植物每年都经历水淹与干旱的交替胁迫。河岸带水位消涨会影响河岸带植物光合作用与生长和发育[3-4]。当水位消涨格局发生显著变化时,河岸带植物光合性能以及生长发育会受到严重的胁迫与损害[5-6]。而光合作用是植物生产力和生物量形成的基础,对植物的生长、发育和繁殖有着决定性的影响[7]。因此,分析人工调节河流模式下水位消涨变化对秋华柳植株生长发育的影响具有重要科学意义。【前人研究进展】植物光合作用的强弱既与自身生理特性有关,还受到环境因子的影响。当生境地的光照、温度、水分以及CO2浓度等环境因子发生变化时[8-9],河岸带植物通过形态学(根系与枝叶数量与形态)[10-11]、生理生化(抗性酶、体内营养物质)组成与含量[12]以及生活史[13]等方面的变化来应对复杂的环境胁迫,在正常的水位涨落情况下,由于植物在长期进化中形成了较强的适应性能,河岸带植物的光合性能以及生长发育受影响程度是有限和可控的[14],光合生理会产生响应,通过调节光合速率、胞间二氧化碳浓度、气孔导度、光补偿点、光饱和点、暗呼吸速率和表观量子效率等光合作用生理特征以及光合色素含量的变化,以减少环境胁迫对自身的损害,尽可能维持正常的光合性能[15-16]。然而三峡工程以及葛洲坝对河流的调节,引起位处大坝下游的秋华柳种群生境地水生态环境大幅变化。其中首要的变化是秋季出露时间的推迟和夏季水淹时间的提早。此外,还引起河流秋季水位下降速率加快和枯水期水位大幅下降,并由此导致生境地土壤含水量的下降[17-18]。人工调节河流导致的水位消涨格局变化可能会影响植物的光合性能[19]。李强等[20]在对三峡库区狗牙根(Cynodondactylon)幼苗的光合生理特征进行研究时发现,大坝对库区水位消涨节律的改变,对该物种的光合性能产生了显著的负面影响。【本研究切入点】国内学者为了对库区消落带进行生态恢复,已从根系生长、光合生理特性、抗性生理、开花物候等方面对秋华柳的水淹性能及其耐淹机制进行了研究[21-23],该物种可以通过产生不定根、调节光合色素比例、维持较高光合速率、提高抗性酶含量等来适应河流原有的水淹节律,但是在水库改变水淹节律条件下,其光合生理是否受到影响不得而知[24-25]。【拟解决的关键问题】通过测定秋华柳在生境地出露初期(植株生长恢复期)以及整个生长期其它阶段的光合生理生态特征,揭示秋华柳光合性能对人工调节河流水位变化的响应,并结合人工调节河流水位消涨格局下生境地水生态环境的变化规律,为该物种的保护和利用提供科学依据。
1 材料与方法
1.1 样地生态环境概况
研究地点位于三峡大坝下游的湖北省枝江市顾家店镇关洲岛(30°15′54.83′′ N,111°34′14.91′′ E;图1),属于亚热带季风性湿润气候区,具有四季分明、冬冷夏热、冬干夏湿的气候特征。该地区雨水丰沛,年平均降水量1212 mm,最大日降雨量183.9 mm。全年积温较高,无霜期较长,约275 d。年平均气温16.7 ℃,极端最高气温 41.4 ℃,极端最低气温-15.6 ℃。生境地土壤类型为砾石和细沙。植被以灌木为主,群落的物种组成主要有:秋华柳(S.variegata)、狗牙根(C.dactylon)、苔草(Carextristachya)、灰绿藜(Chenopodiumglaucum)、蓄萹(Polygonumaviculare)、繁缕(Stellariamedia)等。秋华柳植株个体株高范围为10~20 cm,生长良好。
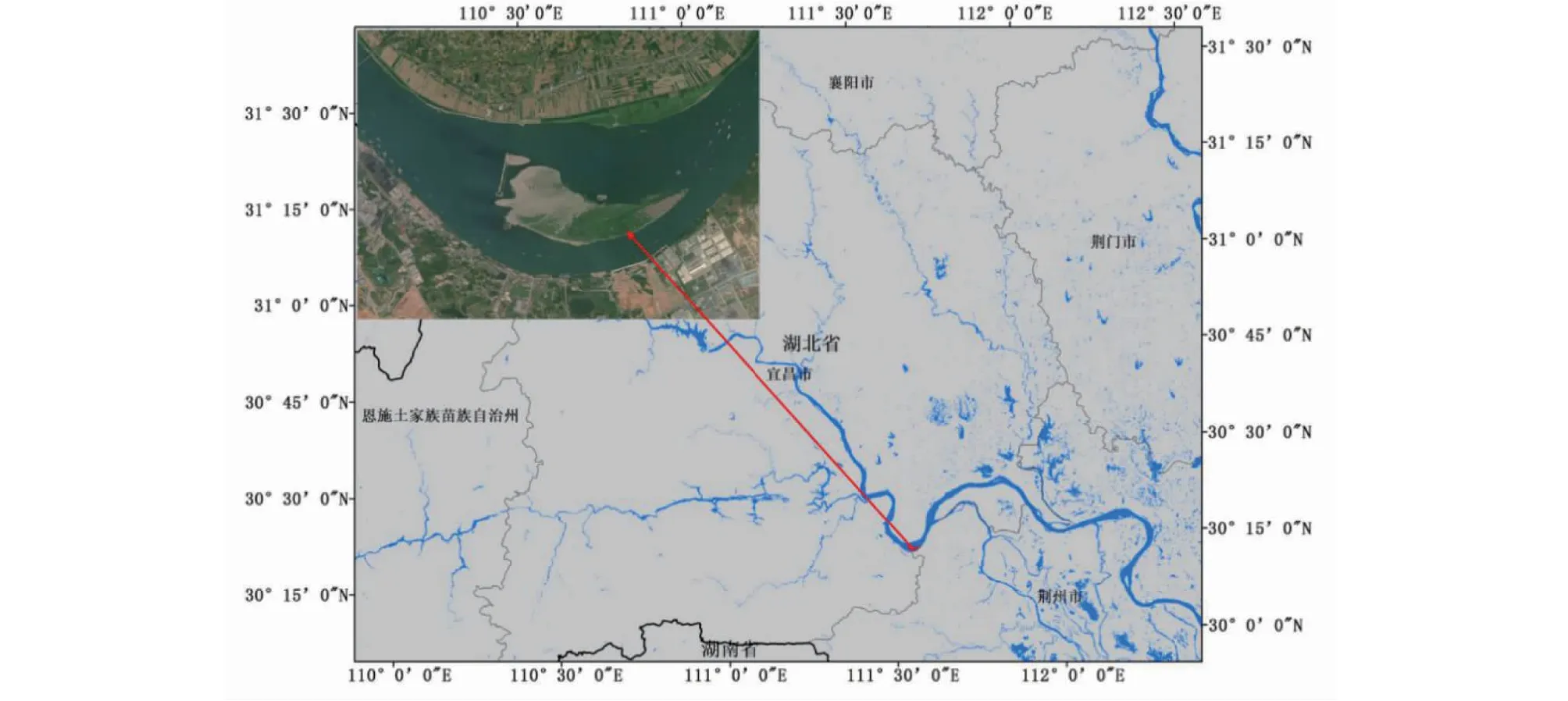
图1 秋华柳野外试验样地地理位置Fig.1 Geographical location of S.variegatafield experimental plot
1.2 样方设置
在研究区域内,在海拔梯度相近的消涨带选择长势良好的秋华柳种群,分别设置5个5 m×5 m的长期样方,每个样方间隔10 m,用于测定秋华柳植株在生长恢复期(出露初期)以及整个生长期其它阶段光合生理生态特征以及环境因子的变化。
1.3 水位变化、土壤含水量与温度的测定
通过资料收集和土壤样品采集与测定,获取秋华柳种群生境地水生态环境的变化资料(图2)。试验地的水位变化以长江水文网(www.cjh.com.cn)枝城段水位公告为依据,并在水位波动期每隔3 d进行一次现场踏查,核实样地的出露与淹没状况。样地出露时间为 2020年10月20日,淹没时间为2021年5月14日。最高温度与最低温度以宜昌气象局公告为依据。在出露初期(出露60 d内)每隔10 d以及整个生长期的前期、中期和后期(出露50、110和170 d),使用环刀在各样方内采集土壤样品(深度15 cm),带回实验室烘干称重,测量土壤含水量。一个样方每次分别随机采集土壤样品3份,整个实验共采集土壤样品120份。
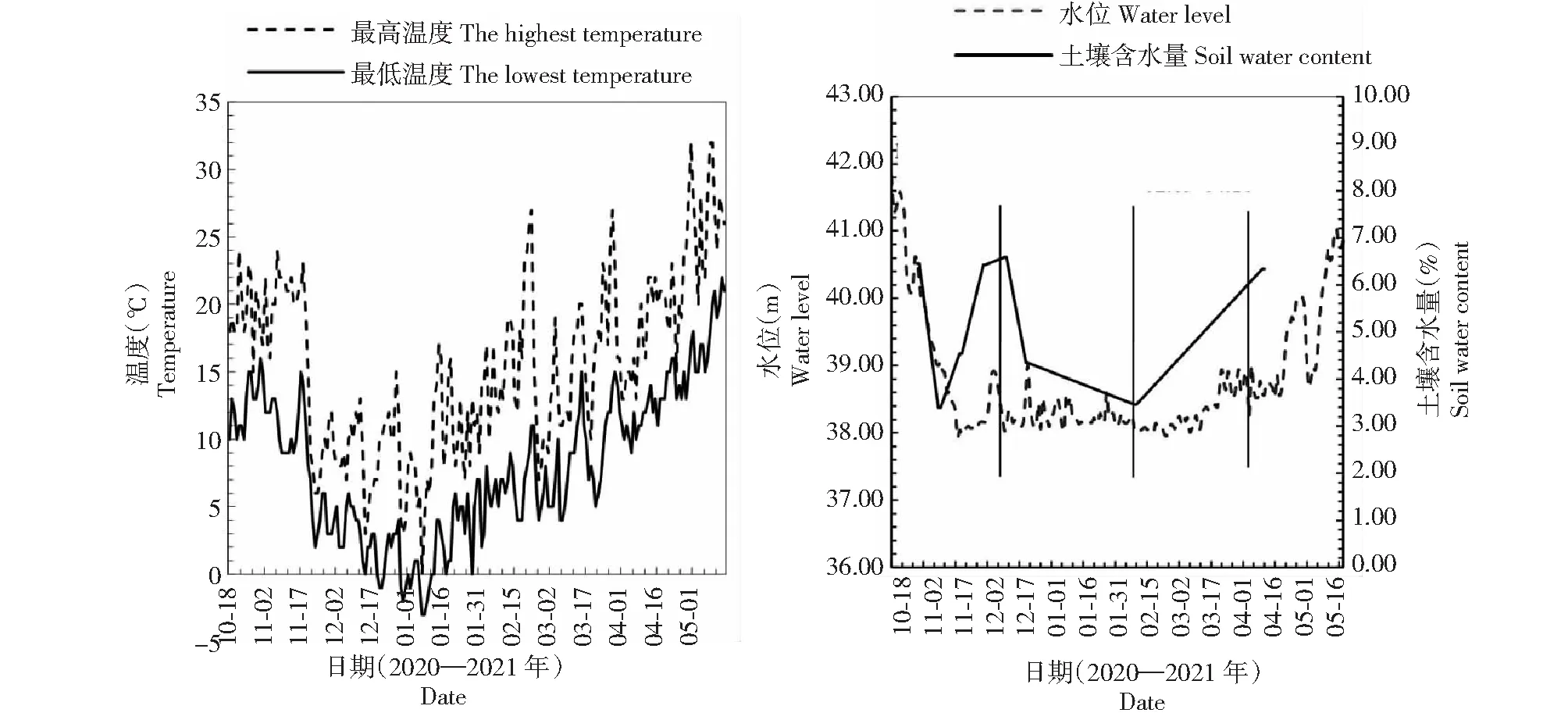
图2 样地温度、水位及土壤含水量在野外试验期间的变化Fig.2 Changes of temperature,water level and soil water content at the sample plot during field experiment
1.4 光合生理生态特征的测定
1.4.1 光合作用的测定 在种群植株整个生长期的前期(50 d)、中期(110 d)和后期(170 d)以及生长恢复阶段(出露60 d)每隔10 d的晴朗天,于每个样方随机选取 1 株大小相近、长势良好的秋华柳植株进行光合生理生态特征的测定。由于秋华柳叶片较小,其幼枝呈绿色,具备一定的光合性能,所以光合生理测定时选取植株一级枝条上部当年生的第 3个或第 4 个二级枝条(包括叶片)作为测试对象。用便携式光合仪(Li-6400)在饱和光强[1000 mol/(m2·s),采用标准红蓝光叶室控制光强]下分别测量枝叶的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间二氧化碳浓度(Ci)。测定时,叶室温度设为20 ℃,流速设为500 μmol/s。每个测定时间各5个重复。光合生理生态特征测定结束后,将所测定枝条剪下带回实验室,60 ℃烘 24 h 至恒重,称量干重(生物量)。用所测枝条的生物量代替叶面积作为测定单位以表征上述所测指标的大小[26-28]。
1.4.2 光响应曲线的测定与拟合 在1.4.1所述各光合作用测定时间和样地所标记的5株秋华柳上另外选择1个一级枝条上部的二级枝条进行光响应曲线的测定。利用红蓝光源分别测定各植物枝条在 2000、1500、1200、1000、500、200、100、50、25、0 μmol/(m2·s)等10个光合有效辐射通量密度下的净光合速率。测定时,叶室温度设为20 ℃,流速设为 500 μmol/s。测后将所测枝条剪下,带回实验室,称其鲜重,然后置于120 ℃烘箱杀青15 min,并在65 ℃烘箱烘7~8 h,烘至恒重,称其干重(生物量),并计算鲜干重比。再用Photosynthesis软件采用非直角双曲线模型拟合光合响应曲线,分别计算植物在恢复生长阶段以及整个生长阶段的最大净光合速率(Pn max)、光补偿点(Lcp)、暗呼吸速率(Rd)和表观量子效率(AQY)等光响应曲线参数。
1.4.3 光合色素的测定 在1.4.1所述各光合作用测定时间,从各样方中选择大小相近/长势良好的1株秋华柳植株,从其上部随机采取一些绿色小枝条,分别放入保鲜袋和冰盒中保存,带回实验室用于测定叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)等光合色素含量。每次5个重复。测定方法如下:分别将所采集的新鲜枝条样品各自称重 0.20 g,剪碎后置于研钵,加入少量石英砂和95%乙醇研磨至匀浆,再加 10 mL 95%乙醇继续研磨至组织变白。将浸泡液倒入25 mL容量瓶中,用95%乙醇进行定容。用95%乙醇作为空白对照,分别测定各溶液在波长为 665、649和 470 nm 处的吸光度(OD)值,每份样品测3次,取平均值。然后计算各样品的叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)含量(mg/L)[29]。
Chla=13.95×A665-6.88×A649
Chlb=24.96×A649-7.32×A665
Chlt(总叶绿素含量)=20.0×A649+8.02×A665
Car=[1000×A470-2.05×C(Chla)-114.8×C(Chlb)]/245
叶绿体色素含量(mg/g)=(C×提取液体积)/样品鲜重
1.5 数据处理
分别以秋华柳光合生理生态特征指标(净光合速率、气孔导度、蒸腾速率、胞间二氧化碳浓度)、光合色素指标(光合色素、Chla、Chlb和Car含量)为因变量,以取样时间为自变量,按生长恢复期和整个生长期进行单因素方差分析(One-way ANOVA),并采用Duncan多重比较分析各处理、水平间的差异显著性。同时进行秋华柳光合生理生态特征指标、光合色素含量与环境因子的Person相关性分析,揭示秋华柳光合作用生理特征与光合色素含量之间及其与环境因子之间的关系。数据分析采用SPSS Statistics 26.0软件进行。
2 结果与分析
2.1 秋华柳植株的光合生理生态特征在生长恢复期阶段的变化
从图3可知,秋华柳植株光合生理在生长恢复期随时间进程呈极显著的动态变化(净光合速率,F=9.360,P<0.05;胞间二氧化碳浓度,F=5.558,P<0.05;气孔导度,F=11.966,P<0.05;蒸腾速率,F=3.921,P<0.05)。植株净光合速率、气孔导度和蒸腾速率在所测定的时间范围内均呈先增加后减少的趋势,而胞间二氧化碳浓度呈先减少后增加的趋势。植株净光合速率、气孔导度和蒸腾速率均在植株出露50 d达到其最高峰值262.88 μmol/(mg·s)、2.62 mol/(mg·s)、25.68 mmol/(mg·s),分别较出露10 d增长413.62%、253.02%、138.95%;胞间二氧化碳在植株出露50 d达到最低值255 μmol/mol,较出露10 d下降20.56%。植株光合生理在夏季水淹后可迅速恢复,并有一段持续50 d左右的快速生长期。表明人工调节河流所带来的水环境变化对下游秋华柳的影响在该物种的适应范围内,秋华柳植株水淹出露后可快速恢复其光合作用性能。
光响应曲线特征值显示秋华柳植株的光合性能随着生长恢复进程显著增强(表1)。最大净光合速率(Pn max)、暗呼吸速率(Rd)、表观量子效率(AQY)随生长恢复进程呈不断增加的趋势,在出露50 d后达到最大值。其中Pnmax、Rd、AQY在出露30~50 d增长较快,分别增长196.63%、247.89%和202.04%。光补偿点(Lcp)则呈不断减少的变化趋势,以出露10~30 d的变化最大,与10 d相比,30 d时的光补偿点下降36.42%。
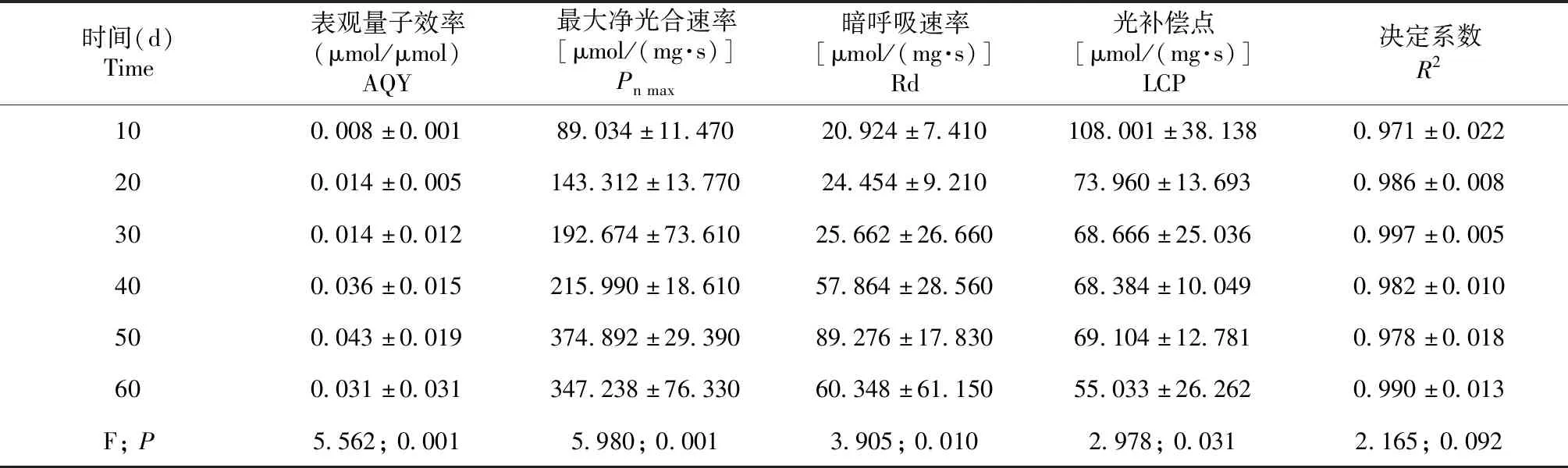
表1 秋华柳植株在生长恢复期的光响应曲线特征值Table 1 Characteristics of light-response curve of S.variegataplants during growth recovering period
2.2 秋华柳植株的光合生理生态特征在整个生长期的变化
从图4可知,秋华柳植株的净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率在整个生长期均呈V字形变化,其中净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率的变化均达到显著水平,中期净光合速率、气孔导度和蒸腾速率较前后期显著下降;中期胞间二氧化碳浓度较前后期显著上升(净光合速率,F=15.172,P<0.05;胞间二氧化碳浓度,F=5.468,P<0.05;蒸腾速率,F=6.209,P<0.05,气孔导度,F=5.650,P<0.05)。秋华柳植株的净光合速率、气孔导度及蒸腾速率均以生长前期最高262.88 μmol/(mg·s)、2.62 mol/(mg·s)、25.68 mmol/(mg·s),分别比中期高59.39%、54.58%、53.39%;秋华柳植株的胞间二氧化碳浓度以生长中期最高297 μmol/mol,比前期高16.47%。从整个生长期来看,夏季水淹后秋华柳植株的光合生理在生长前期最旺盛,而生长中期光合生理显著下降,表明生长中期的光合生理生长可能受到限制。
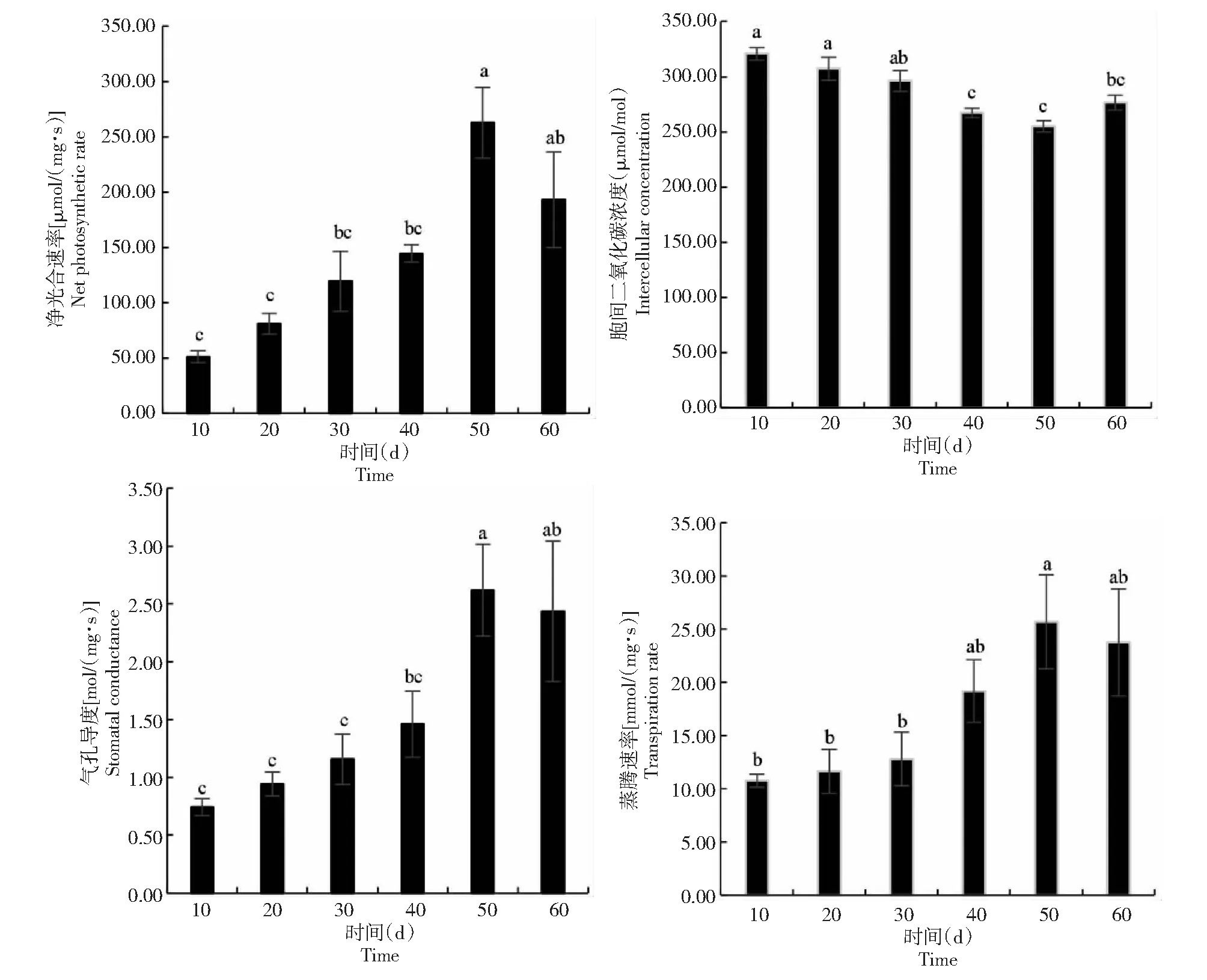
图中不同小写字母表示处理间的差异显著性 (P<0.05) ,下同。Different lowercase letters in the picture indicate significance of difference between treatments (P<0.05).The same as below.图3 秋华柳植株的光合生理生态特征在生长恢复期的变化Fig.3 Changes of S.variegataplants in photosynthetic physiology during the growth recovering period
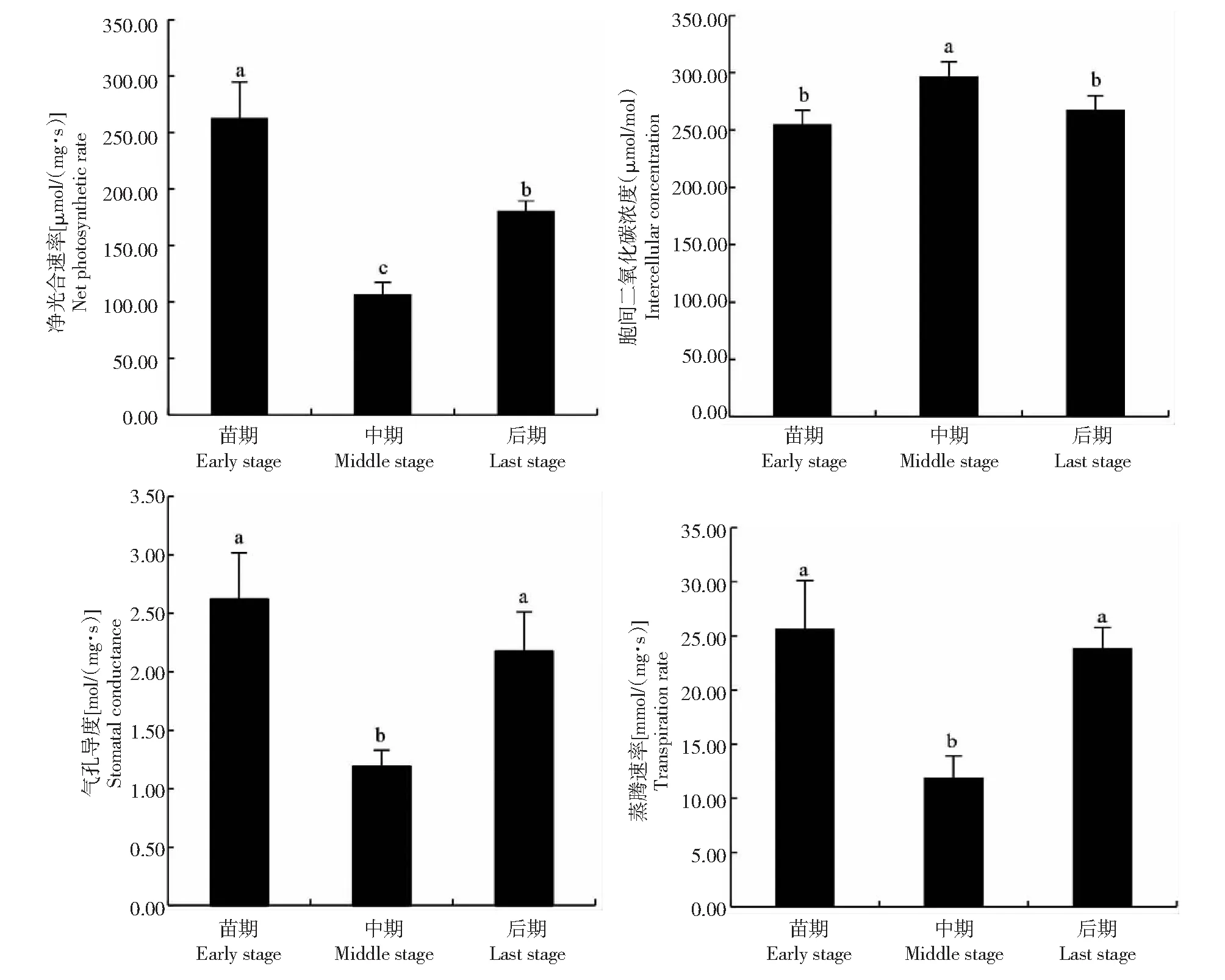
图4 秋华柳植株的光合生理生态特征在整个生长期的动态变化Fig.4 The dynamic changes in photosynthetic indices during the whole recovering growth periods of S.variegataplants
光响应曲线特征值显示秋华柳植株的光合性能在整个生长期有着较大的变化(表2)。Pn max、Rd、LCP、AQY均呈V字形的变化趋势。其中Pn max、AQY和LCP变化显著,而Rd变化不显著。生长前期的Pn max和AQY分别比生长中期高63.49%和76.36%;LCP比生长中期高10.17%。

表2 秋华柳植株整个生长期不同阶段的光响应曲线特征值Table 2 Characteristics of light response curve of S.variegataplants at different growth periods
2.3 秋华柳植株的光合色素在生长恢复期的变化
从图5可知,秋华柳植株的Chla和Chlb含量、Car含量、总叶绿素含量在生长恢复期不同时间之间存在显著差异(Chla,F=248.024,P<0.05;Chlb,F=21.626,P<0.05;Car,F=583.208,P<0.05;总叶绿素含量,F=258.680,P<0.05)。随着生长恢复进程的推进,秋华柳植株的Chla、Chlb、Car和总叶绿素含量整体呈上升趋势,在生长恢复50 d时达到最大值,分别比生长恢复10 d增长77.89%,42.37%、82.97%和66.50%。Chla与Chlb的比值在生长恢复期呈显著上升趋势(F=7.648,P<0.05),而总叶绿色素和Car的比值则呈显著下降趋势(F=14.746 ,P<0.05)。秋华柳植株的光合色素在夏季水淹后可快速恢复积累,并有一段持续50 d左右的快速积累期。

图5 秋华柳植株生长恢复期不同阶段光合色素的动态变化Fig.5 Changes of S.variegataplants in the photosynthetic pigment content during the recovering growth period
2.4 秋华柳植株的光合色素在整个生长期的变化
从图6可见,秋华柳植株的光合色素含量在整个生长期呈V字形变化,且中期较前后期显著下降(Chla,F=129.324,P<0.05;Chlb,F=57.683,P<0.05;Car,F=175.974,P<0.05;总叶绿素含量,F=140.208,P<0.05)。秋华柳植株的光合色素在水淹后整个生长期的变化趋势与光合生理生态特征变化一致,Chla、Chlb、Car、总叶绿素含量在生长前期最高,中期最低。秋华柳植株生长前期的Chla、Chlb、Car和总叶绿素含量分别比生长中期的高29.26%、34.16%、23.53%和30.61%。秋华柳植株的Chla与Chlb的比值在整个生长期有所上升,但变化不显著(F=2.735,P>0.05)。秋华柳植株的总叶绿素和Car的比值随时间进程呈下降趋势,后期较前期显著下降(F=28.161,P<0.05)。
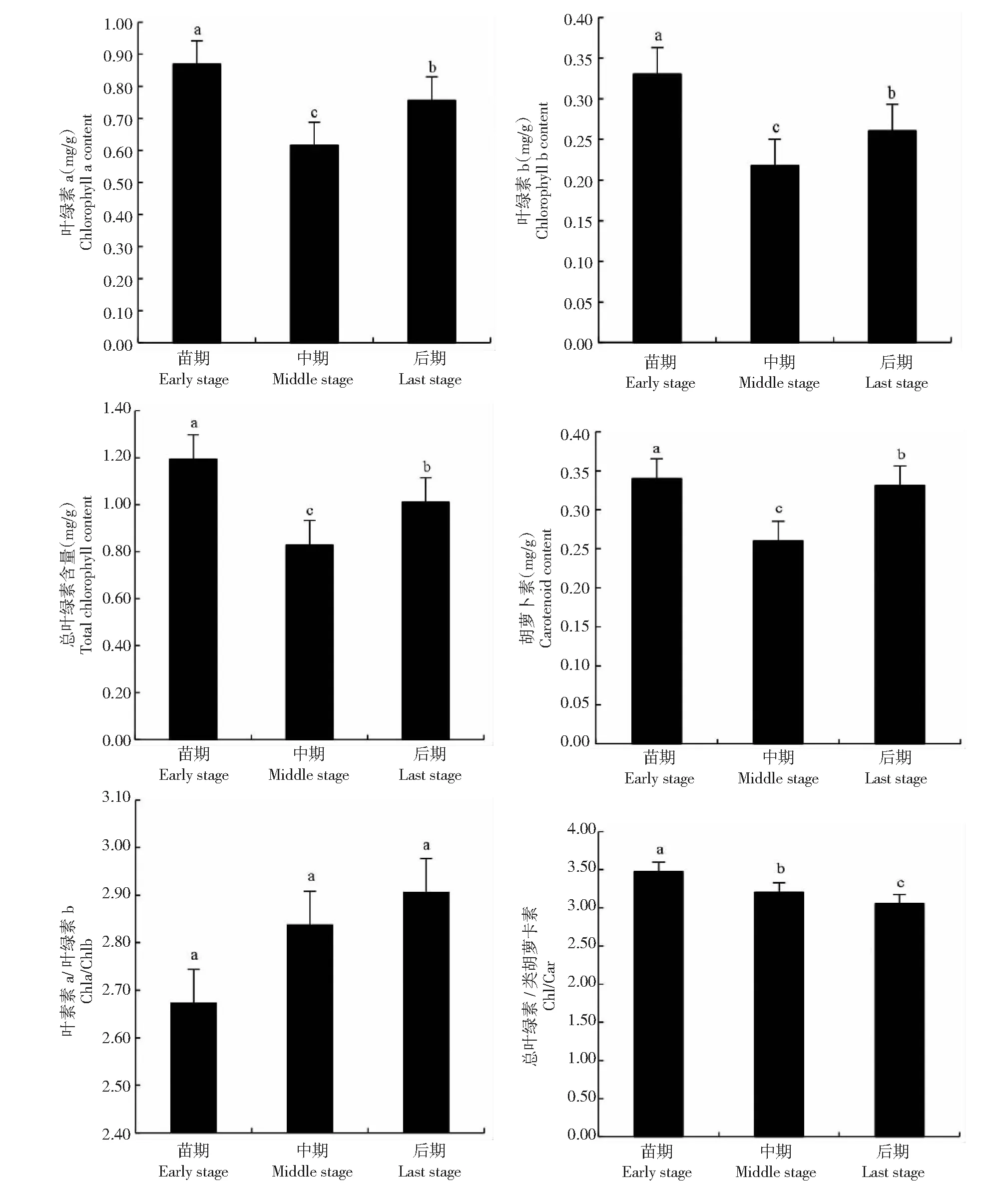
图6 秋华柳植株的光合色素在不同生长期的动态变化Fig.6 Changes of S.variegataplants in photosynthetic pigment contents during different growth periods
2.5 秋华柳植株生长期不同阶段光合作用性能的综合评价
将秋华柳植株的净光合速率、气孔导度、胞间二氧化碳、蒸腾速率、叶绿素a、叶绿素b、总叶绿素、类胡萝卜素8项指标进行标准化(Z-score)处理,数据间关联度较高,表明综合评价秋华柳的光合作用性能是可行的。运用主成分分析得到各指标载荷矩阵及贡献率。由表3可知,第一主成分以秋华柳植株的叶绿素a、叶绿素b、总叶绿素、类胡萝卜素载荷较高,主成分贡献率为74.836%,第二主成分则主要反映秋华柳植株的光合生理的信息,主成分贡献率为10.561%,2个主成分累计贡献率为85.397%,表明秋华柳植株的光合色素对光合作用性能的影响较大。

表3 主成分分析载荷矩阵及贡献率Table 3 Different index component load matrix and contribution rate
根据特征值及主成分载荷矩阵得出各指标系数,将系数与标准化的数据处理得到主成分解析表达式,将标准化后的原始数据带入主成分解析表达式,得到不同时期在2个主成分的得分,再对主成分进行加权可得到不同时期的综合得分公式。通过对不同时期各项指标各主成分得分及综合得分进行比较排序,最终得到主成分得分(表4),恢复生长前期秋华柳植株的光合作用性能在第一主成分得分、第二主成分得分与综合得分远远高于其他时期。

表4 整个生长期不同阶段的综合得分Table 4 Comprehensive scores of different stages in the whole growth period
2.6 光合特性和光合色素与环境因子间的相关性分析
秋华柳植株的光合生理与光合色素含量密切相关(表5)。其中净光合速率与Chla、Chlb、总叶绿素、Car含量之间呈极显著相关(P<0.01);气孔导度与Chla、Chlb、总叶绿素、Car含量之间呈显著相关(P<0.05);胞间二氧化碳浓度与Chla、总叶绿素含量呈显著相关(P<0.05),与Car含量呈极显著相关(P<0.01);蒸腾速率与Chla、Car含量呈极显著相关(P<0.01),与总叶绿素含量呈显著相关(P<0.05)。
秋华柳植株光合生理生态特征和光合色素变化与环境因子之间均存在一定相关性(表5)。其中净光合速率与土壤含水量呈极显著相关(P<0.01),蒸腾速率与土壤含水量、地下水位呈极显著相关(P<0.01),气孔导度和胞间二氧化碳浓度与土壤含水量呈显著相关 (P<0.05)。Chla、Chlb、总叶绿素、Car含量与土壤含水量均呈极显著相关(P<0.01),其中Car还与地下水位呈显著相关(P<0.05)。光合生理生态特征、光合色素与日照时长、平均温度之间无显著相关。

表5 光合生理生态特征、光合色素和环境因子间的相关性分析Table 5 Person correlation analysis between photosynthetic physiologic characteristics,photosynthetic pigments and environmental factors
3 讨 论
3.1 秋华柳植株在生长恢复期光合生理生态特征的变化
水淹是河岸带植物每年都要经历的环境胁迫。在水淹及其所引起的缺氧胁迫下,植物叶片的光合色素会发生分解,并由此引起光合色素含量和光合作用下降的变化[30]。由于胁迫期间植物仍需消耗大量的营养物质来维持其必需的生理活动,胁迫后的植株多出现营养物质匮乏的现象[32]。很多植物的光合生理在水淹后的生长恢复期常出现一个快速增长(补偿性生长)的过程,以及时补充植物生长发育所需的有机营养物质[33]。秋华柳植株在水淹出露后开始恢复生长,光合生理活动呈一个快速增长的过程,其净光合速率、气孔导度、蒸腾速率均随着生长恢复进程不断上升,在生长恢复期的后期(50 d)达到最大值。与此同时,秋华柳植株的Pn max、AQY、Rd等光响应曲线特征值也随着生长恢复进程不断上升,并在生长恢复期的后期(50 d)达到最大值。表明人工调节河流所带来的水环境变化对下游秋华柳的影响在该物种的适应范围内,经历长时间水淹胁迫的秋华柳植株在胁迫解除后也存在一个补偿性生长过程,植物通过快速恢复其光合作用性能,及时同化和合成有机营养物质来弥补胁迫期间的损耗,并满足恢复期枝叶快速生长的需要。关于水淹后植物光合生理的补偿性快速增加,Chen 等[34]在对狗牙根(Cynodondactylon)耐水淹机理的研究中也有报道,狗牙根在经历45 d的水淹胁迫后,经过7 d的恢复生长,其净光合速率显著高于未经过水淹胁迫的植株。
叶片光合色素承载着光能的捕获、分配、传递和转化的功能[35]。叶绿素两大组分间的比值(Chl a/b )和总量(Chl a+b )在一定程度上表征了整个光系统对光量子的捕获和利用状况[36]。类胡萝卜素的主要功能是清除体内活性氧和保护光合膜系统,其含量与在光合色素中的占比反映了植物对环境胁迫的抵抗性能[37]。植物在生长恢复期为了尽快恢复其光合性能,叶片的光合色素含量通常会发生快速增长[33]。Guan等[38]对疏花水柏枝的研究发现,植株的光合色素含量在水淹胁迫解除后都有一个快速增长的过程,伴随而来的是最大净光合速率的快速增长。本研究中,进入恢复生长的秋华柳植株,其总叶绿素、Chla、Chlb和Car含量均快速增长,并在出露50 d时达到最大值。这种变化与光合作用性能的变化趋势是相同的,由主成分分析结果可见,秋华柳植株光合作用性能在恢复期的增长得益于光合色素的合成与含量的增长。
3.2 秋华柳植株光合生理生态特征在整个生长期的变化
长江流域的河岸带植物每年在夏末至秋初随着洪水的消退而出露,然后进行生长恢复、开花结果,一直到次年的春末或夏初再次被洪水淹没。河岸带植物的生长期含盖整个冬季,但由于冬季气温偏低与少雨,其生长相对缓慢,秋季和春季温度相对较高是该地区河岸带植物快速及主要的生长期[39]。河岸带植物的光合生理性能通常也与植物生长呈相同的变化格局[42]。Guan等[38]对三峡大坝下游疏花水柏枝(Myricarialaxiflora)残存种群的光合生理性能进行了研究,发现该物种的光合生理性能在整个生长期呈V字形的变化,即以前期和后期高,中期最低,但是后期高于前期。本研究中秋华柳植株光合生理性能在整个生长期也呈V字形的变化规律,既以中期的光合生理性能最低,但其前期高于后期。主成分分析结果也显示,恢复生长前期下的秋华柳光合作用性能在第一主成分得分、第二主成分得分与综合得分远远高于其他时期。植株在水淹胁迫后出露初期的补偿性生长和冬季气温偏低对植物生长的影响被认为是植株光合生理性能呈V字形变化的主要原因[33,38]。
秋华柳植株光合色素含量在整个生长期的变化与光合生理性能的变化呈相同趋势,Chla、Chlb、总叶绿素与Car含量在出露初期升高,中期显著下降,后期又一次显著上升。前期研究测定了水淹对秋华柳光合生理生态特征的影响,正常水位下秋华柳的Chla、Chlb分别为1.474和0.485 mg/g[43],其值均高于本研究中植株的光合色素含量。植物可通过调整Chla/b和Chl与Car的比值来适应环境的变化以维持较高水平的光合生理生态特征[44]。正常情况下,植物叶片Chl a/b 和Chl/Car均约为3∶1[45]。本研究中秋华柳在整个生长期Chl a/b小于3∶1,Chl/Car大于3∶1。但随着植物出露时间的延长,Chl a/b逐渐增加,而Chl/Car逐渐减少。表明该物种植株在整个生长期大多数时间均处于逆境中,植物的光合作用性能随着出露时间的延长不断恢复。植物出露初期叶片的光合性能高于出露后期,而抗逆性能低于出露后期。出露初期植株叶片以相对较高的光合反应中心来提高光合性能,而出露后期则通过提高光捕获能力来维持相对高的光合性能。
3.3 人工调节河流对下游秋华柳植株光合生理生态特征的综合影响
水位消涨及其所引起的生态环境变化是影响河岸带植物生长发育的主要生态过程[30,46]。水位消涨会导致河岸带植物生境地的出露与水淹,并引起土壤含水量、土壤营养(有机质和养分)和土壤结构的变化,由此导致河岸带植物光合生理生态特征的变化[47-49]。本研究结果显示,秋华柳的光合性能与水位消涨及其所引起的环境因子变化密切相关。其中,土壤含水量与光合生理及光合色素之间呈显著相关,水位与蒸腾速率、类胡萝卜素之间呈显著相关。表明该物种的光合性能极易受水位变化及其所引起的土壤含水量变化的影响。
河流水位消涨格局的变化常常会对植物的光合生理产生严重影响,并由此影响植物的生长与发育[15,50]。大西洋沿海濒危物种粉花金鸡菊(Coreopsisrosea)的生长便因不合理的水位消涨格局而受到严重威胁[51]。三峡大坝与葛洲坝因防洪发电的需求对河流进行了调控,由此改变了其下游秋华柳生境地的水位消涨格局和生态环境。生境地的出露时间由9月初推迟到了10月下旬甚至11月中上旬,而水淹时间则由6月下旬提前到5月上旬,导致生境地水淹时间延长,生长期缩短。此外,人工调节河流还导致水位下降速率加大和枯水期水位显著下降,由此引起生境地土壤含水量全面降低。由于秋华柳的光合生理与水位消涨及其所引起的环境因子变化密切相关,水位涨落节律改变引起的环境因子变化,会较大程度影响植物的光合生理生态特征。如恢复初期水位急剧下降和枯水期的低水位引起的土壤含水量下降,使光合作用生理活动下降;生长期的缩短会减少植物光合作用的时间,减少有机物质的合成与贮藏。因此,三峡大坝与葛洲坝对河流水位消涨格局的改变会进一步影响秋华柳种群植株的生长和发育。
4 结 论
(1)秋华柳种群植株的光合作用性能在水淹后的生长恢复阶段有一个快速增长的过程,其净光合速率、气孔导度、蒸腾速率、表观量子效率均随生长恢复进程不断上升,在生长恢复期的后期(50 d)达到最大值,其后逐步下降。在水淹后的生长恢复阶段,秋华柳植株光合色素含量的变化与光合生理性能的变化呈相同趋势。
(2)在整个出露后的生长期,秋华柳植株的光合生理活动随生长期延伸而增强;在生长中期,秋华柳植株的光合生理特征指标显著下降,中期的光合生理性能最低,但植株前期光合生理性能高于后期光合生理性能。秋华柳植株光合色素含量在整个生长期的变化与光合生理性能的变化呈相同趋势。Chla、Chlb、总叶绿素与 Car 含量在出露初期升高,中期显著下降,后期又一次显著上升。
(3)三峡大坝及葛洲坝人工调节河流水位消涨格局引起的水生态环境变化,使秋华柳种群光合作用生理活动下降,进一步影响该种群的生长和发育。
