重金属镉对植物原初光能转化的影响
2023-04-14唐佳乐于志民李润唐
唐佳乐,于志民,李润唐,王 静
(湛江科技学院,广东 湛江 524094)
土壤重金属污染是世界各国面临的重大生态问题,重金属镉是中国土壤的首要重金属污染物。镉作为一种生命非必需元素,具有极强的生理毒性,能够通过离子取代、诱导氧化应激等途径破坏信号传导通路及细胞生物膜结构,影响光合作用、呼吸作用等重要生理代谢过程,最终抑制植物的生长与发育,甚至造成植物死亡。光合作用效率降低是影响植物生长的根本原因,光合作用是光能被生物组织捕获、储存、转化的过程,光合能量最终用于驱动细胞新陈代谢[1]。高等植物的光合作用主要可以分为2 个阶段,光反应阶段和碳反应阶段,其中光反应阶段(即原初光能转化过程)主要涉及植物对光合能量的捕获、传递、转化及光合电子传递等重要过程。随着叶绿素荧光动力学及激光共聚焦荧光寿命成像显微技术的发展,植物光合原初能量转化过程得到了有效的监测[2,3]。本研究针对植物光合能量的捕获、传递及转化过程对重金属镉的响应展开论述,旨在揭示重金属镉对叶片光合能量捕获、传递及转化的影响机理,为减缓重金属镉的胁迫,提高植物光能吸收能力和光能传递效率,促进光能转化提供理论依据。
1 重金属镉对植物叶绿素及类囊体膜结构的影响
1.1 重金属镉抑制叶绿素合成、促进叶绿素降解
高等植物中光合色素主要包括叶绿素(Chl a、Chl b)及类胡萝卜素。叶绿素除捕获、传递光能之外,还在主体捕光色素蛋白复合体(LHC Ⅱb)三聚体形成过程中具有重要作用[4],少数特殊Chl a 分子还具有光敏化特性,参与光化学反应(电荷分离)。重金属镉对光合色素影响的研究结果较为一致[5-8],即镉胁迫下植物叶绿素含量显著减少,约3周龄的菠菜幼苗经90 mg∕kg Cd2+处理10 d 后,叶绿素含量下降68%[9]。即使是较低浓度的镉胁迫也会导致植物叶绿素含量的降低,灯芯草经1 μmol∕L Cd2+处理15 d 后叶绿素含量下降达27.3%[10]。
叶绿素生物合成的受阻是造成叶片内叶绿素含量降低的主要原因,高等植物中叶绿素合成最少需要15 个过程并涉及15 种不同的生物合成酶[11],重金属镉通过与相关酶活性位点的巯基(-SH)结合[12]或下调叶绿素合成酶相关基因的表达[8],干扰5-氨基酮戊酸脱水酶(ALAD)、胆红素原酶(PBGase)、尿卟啉原脱羧酶(UROD)、NADPH 原叶绿素酸酯氧化还原酶(POR)等酶的活性,使叶绿素合成受阻。此外,重金属镉引起的缺铁也会抑制叶绿素的生物合成[13]。
重金属镉会通过促进叶绿素的降解以减少叶绿素的含量,通过荧光谱带观测到镉胁迫下菠菜叶片内存在大量叶绿素降解产物[9],在另一项研究中也观察到大量与叶绿素降解途径相关的基因(ACD2、NYE1、SGR)的表达量显著上调[8]。有研究表明,重金属镉对叶绿素中心原子Mg2+具有较强取代作用,仅需30 min 就可将80%的Chl a 转化为缺镁叶绿素[14],从而降低叶片内有效叶绿素的含量。
因此,重金属镉通过抑制叶绿素合成、促进叶绿素有降解及离子取代等途径降低叶片内叶绿素含量及有效叶绿素含量,进而影响叶片对光能的捕获及转化效率。
1.2 重金属镉影响叶绿素的组成
叶绿素组成也直接影响光系统Ⅱ捕光色素蛋白复合体(LHCⅡb)的结构及功能。要形成稳定的LHCⅡb 三聚体,Chl a ∕Chl b 只能在一定范围内变化,且必须保证每个三聚体中至少有一个Chl a 分子,Chl b 的合成对LHCⅡb 三聚体的含量也有一定的影响[15]。
不同的植物中Chl a 与Chl b 对重金属镉的敏感性有所差异,有研究表明桑树、辣椒等植物在Pb、Cd胁迫下Chl a ∕Chl b 显著下降[16,17]。将辣椒幼苗分别置于含0、100 μmol∕L Cd2+的MS 培养基中,Chl a∕Chl b 由3.40 下降为2.87。因此,在这些植物中Chl a对重金属镉胁迫可能更为敏感。但有研究提出,Chl b 较Chl a 对重金属更加敏感,重金属镉胁迫下龙葵及茄子Chl a∕Chl b 较非胁迫状态下更大[18]。Chl a 与Chl b 对重金属镉的敏感性差异可能与胁迫强度及植物种类有关,需要进一步探讨。
1.3 重金属镉对类囊体膜成分与超微结构的影响
类囊体是由色素、脂类物质及蛋白质组成的膜系统,是叶绿体中执行光能吸收与转化的场所。因此,类囊体膜结构及功能的完整性是植物进行光合作用的基础。研究表明,进入叶绿体的镉离子有80%积累在类囊体膜中[19],造成类囊体膜结构的解体。
类囊体膜结构的受损与膜脂过氧化有关,各类生物及非生物胁迫通常会导致膜脂过氧化的最终产物丙二醛(MDA)含量的增加。组成类囊体膜的脂类物质主要包括磷脂及糖基甘油酯,二者均富含多不饱和脂肪酸[4]。重金属镉可以通过脂氧合酶(LOX)介导多不饱和脂肪酸的氧化,产生自由基,进而导致类囊体膜的破坏,如番茄幼苗经100 μmol∕L Cd2+胁迫10 d 后,叶片内脂氧合酶(LOX)活性显著增加,同时MGDG(单半乳糖基甘油二脂)、DGDC(双半乳糖基甘油二脂)及PG(磷脂酰甘油)含量显著下降,类囊体膜严重受损且基粒解体[20]。
植物PSII 光合放氧中心及LHCⅡb 富含PG,而PG 分子中的反式十六碳-烯酸(trans-16:1)对保持LHCⅡb 寡聚体结构起重要作用[4]。当PG 分子中trans-16:1 含量降低时,LHCⅡb 的聚集度下降,LHCⅡb 单体数量增加,导致LHCⅡb 捕获的光合能量无法有效地传递至反应中心,捕光效率下降[21]。而trans-16:1 含量极易受到外界环境的影响。因此,重金属镉是否影响PG 分子中trans-16:1含量及trans-16:1 含量对LHCⅡb 结构的影响机制值得进一步的研究。
2 重金属镉对植物PSⅡ光合能量捕获及传递的影响
2.1 PSⅡ光合能量捕获及传递过程
植物PSⅡ光合能量捕获、传递及转化过程如图1 所示。激发能从PSⅡ捕光天线系统到反应中心存在2 条途径,①由主体外周捕光天线复合体(LHCⅡb)中的色素分子吸收光量子,通过辅助外周捕光天线CP26 将激发能传递到内周捕光天线CP43 中的色素分子,最后再传递至反应中心的P680,进行稳定的光化学反应;②由主体外周捕光天线复合体(LHCⅡb)中的色素分子吸收光量子,通过辅助外周捕光天线CP29 及CP24 将激发能传递到内周捕光天线CP47 中的色素分子,最后再传递至反应中心的P680,进行稳定的光化学反应。
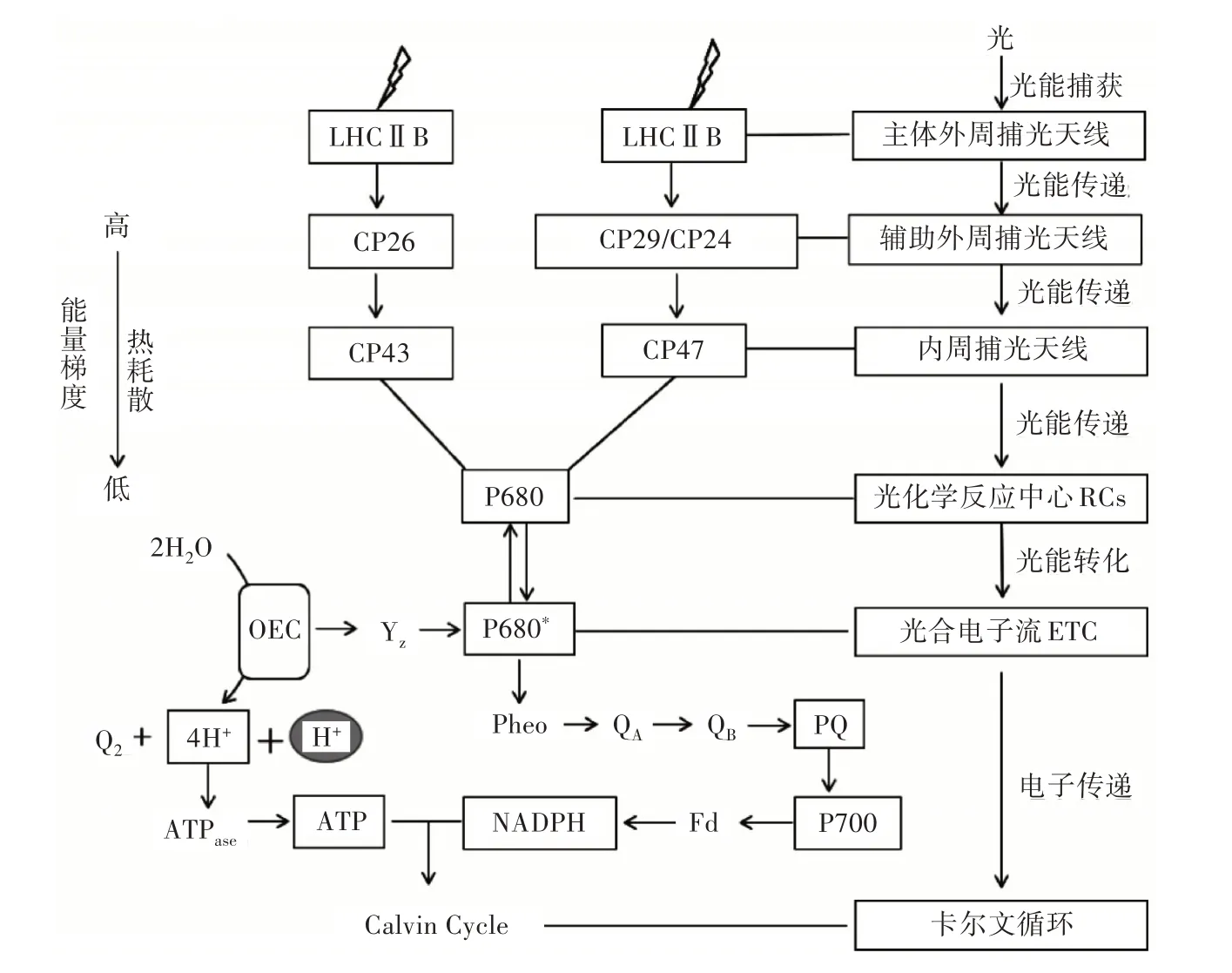
图1 植物对光能的吸收、捕获及转化过程
2.2 重金属镉对植物PSⅡ光合能量捕获的影响
2.2.1 植物PSⅡ光合能量捕获过程对重金属镉的响应 光合作用始于捕光系统对光能的捕获,光合能量的捕获效率直接影响光化学反应效率。重金属镉对植物光能捕获效率的影响尚未得出一致的结论。
大量研究表明重金属镉会促进叶片单位活性反应中心(RC)对光能的捕获,但单位面积叶片捕获的光能在镉胁迫下降低。如25 mg∕L Cd2+胁迫下梭鱼草叶片单位活性反应中心吸收的光能较对照增加32.97%[22],水稻[23]、烟草[24]、番茄[25]等植物中也呈现类似结果;单位面积叶片捕获光能在镉胁迫下降低的原因主要表现为单位活性中心数量(RC∕CS0)的减少,如杂交狼尾草(Hybrid pennisetum)在镉胁迫(c(Cd2+)=10~100 mg∕kg)下有18%~23%的光反应活性中心转化为非活性中心[26],最终导致单位面积上吸收的光能减少[27]。
有研究表明,鸡爪槭处于高浓度镉胁迫(c(Cd2+)>200 mg∕kg)时,其ABS∕RC 值较对照有所降低[28],而另一项研究中发现梭鱼草叶片在镉胁迫下ABS∕CSm较对照增加28.53%,但单位面积热耗散的能量(DI0∕CS0)也响应增加了80.16%[22],这可能是不同的研究材料所处环境差异所致。
2.2.2 重金属镉对植物PS Ⅱ光合能量捕获的影响机制 主体外周捕光天线系统(LHCⅡb)结合叶片中约45%的叶绿素,起主要的光能捕获功能[4]。因此,重金属镉对植物光能捕获效率的影响在本质上是重金属镉对LHCⅡb 结构及功能的影响。
在植物体内LHCⅡb 主要以三聚体的形式存在,也有单体和寡聚体形式的LHCⅡb存在,LHCⅡb寡聚体和单体中不但色素吸收的光能减少,且从类胡罗卜素到Chl b 及从Chl b 到Chl a 的能量传递效率均低于三聚体[4]。因此,LHCⅡb 三聚体结构的形成是高效光能捕获的前提。
重金属镉会降低LHCⅡb 的聚集度[29]。重金属镉可能通过降低磷脂酰甘油PG 及反式十六碳-烯酸(trans-16:1)的含量影响LHCⅡb 的聚集度,进而降低LHCⅡb 对光能的捕获效率。
光合色素对LHCⅡb 的聚集度也具有一定的影响,叶绿素b 在维持LHCIIb 结构稳定中具重要作用[30,31]。拟南芥中Chl a ∕Chl b 由2.85 下降为2.65时,其外周捕光色素蛋白复合体含量增加20%[15],而在缺失Chl b 的莱茵衣藻突变体中,PSⅡ天线的叶绿素从320 分子降到了95 分子[32]。此外,要形成稳定的LHCⅡb 三聚体,至少要保证每个三聚体中有一个Chl a[33]。因此,重金属镉可以通过影响叶绿素的含量及组成来影响LHCⅡb 的结构,进而影响LHCⅡb 对光能的捕获效率。
此外,重金属镉可以通过影响LHCⅡb 状态的转变来影响LHCⅡb 对光能的捕获效率。当植物处于胁迫条件下时,捕光系统捕获的光能无法被及时消耗,植物叶片可能通过LHCⅡ的磷酸化引起的状态转换及LHCⅡ的可逆脱离降低PSⅡ捕光系统有效吸收截面,从而抑制捕光复合体对光能的吸收[34]。
在镉胁迫下植物叶片单位活性中心对光能捕获的增加的同时常观察到LHC Ⅱb 表观尺寸的增大[35]。在PS Ⅱ-LHC Ⅱ超级复合体周围存在一定数量的含Chl a 较少的辅助捕光色素蛋白单体(CP29、CP26、CP24)[36]及部分没有结合或松散结合的LHCⅡb 三聚体,当植物处于逆境胁迫时,叶绿素含量的降低及光反应活性中心的部分失活导致叶片对光能的捕获能力下降,PS Ⅱ-LHC Ⅱ超级复合体将结合更多的单体捕光色素蛋白及游离的LHCⅡ三聚体,以增强叶片的捕光能力[37]。
重金属镉会通过改变LHCⅡb 结构及功能来降低植物叶片光能捕获效率,而单个LHCⅡb 对镉胁迫的响应存在差异,可能与镉胁迫程度及植物种类有关。
2.3 重金属镉对植物PS Ⅱ光合能量传递的影响
主体外周捕光复合体LHC Ⅱb 捕获光量子后通过微量LHCⅡ(CP29、CP26、CP24)将激发能传递给内周捕光复合体(CP43、CP47),而内周捕光复合体将进一步激发能传递到反应中心D1∕D1 异二聚体上引起电荷分离(P680→P680*),因此,光合能量的传递效率直接影响着光化学反应的正常进行。
重金属镉会抑制光合能量向PSⅡ反应中心的传递。在正常生理状态下,植物的最大光化学效率Fv∕Fm较为稳定,一般为0.80~0.85[37],而在干旱、高温、高盐、强光等胁迫下常表现一定程度的下降,镉胁迫下水稻[38,39]、辣椒[40]、龙葵[18]、芥菜[41]等植物中均发现Fv∕Fm随镉胁迫程度加深而降低,其中不乏一些镉超富集植物(龙葵及芥菜)。由此,PSⅡ反应中心对激发能的捕获效率较易受到镉胁迫的抑制。此外,有研究表明镉胁迫下光合能量分配到光化学反应中心的比例(TRo)降低,而热耗散比例(DIo)显著提高[39,42],即镉胁迫抑制了光合能量的传递过程,导致外周捕光复合体捕获的光合能量无法正常传递到光化学反应中心[39],使更多的激发能在天线系统中热耗散的形式消耗,以减少过剩激发能对光系统的伤害[39,42]。
重金属镉抑制光合能量传递的机制尚不清楚,理论上天线色素吸收的光子有95%~99%的能量可以传递至反应中心,而天线色素间的能量传递属于物理现象(荧光共振能量转移(FRET),并不容易受到外界环境的干扰,因此,较为可能的原因是进行光合能量传递的载体受到镉胁迫的抑制,通过蓝色非变性凝胶电泳(BN-PAGE)技术及蛋白质组技术发现镉胁迫下鱼腥藻[35](Anabaena)、烟草[43]等PSⅡ复合体CP43 及CP47 蛋白含量显著降低,进而抑制了激发能向光化学反应中心的传递。
3 重金属镉对植物PSⅡ光合能量转化的影响
3.1 植物PSⅡ光合能量捕获效率对重金属镉的响应
当激发能传递到PS Ⅱ光化学反应中心特殊叶绿素分子P680(P),产生激发态P680*,而P680*具有极强的还原性,将在3ps 内将1 个电子传递给缺镁叶绿素(Pheo)而产生激发态P+·Pheo-,通过这一过程,光合能量由激发能转化为氧化还原能[1]。理论上光能的传递效率超过95%,即被吸收的光量子都能够参与原初光物理反应,但每个光子仅有25%的能量可以被转化为化学能,其余能量转化为热能,而在实际情况中C3 植物PSⅡ能量转化效率大约为4.3%[44],此外,PS Ⅱ能量转化效率还会受到各类环境因素的限制。
重金属镉会抑制PSⅡ反应中心将吸收的激发能转化为氧化还原能。在一定浓度的镉胁迫下常发现PSⅡ实际光化学效率的下降,如超过0.2 mmol∕L的Cd2+胁迫会使部分品种水稻实际光化学效率(ΦPSⅡ)及光化学猝灭系数(qP)显著下降[38,39],超富集植物龙葵及芥菜等植物在较低浓度(约25~40 μmol∕L)镉胁迫下ΦPSⅡ及qP 即受到抑制[41,45],但有研究表明部分品种水稻在镉胁迫(0.2 mmol∕L)下依然能保持较高的实际光化学效率[38],甚至在低浓度(0.1 mmol∕L)下略有上升[39]。植物叶片PSⅡ反应中心将激发能转化为氧化还原能的过程对重金属镉较为敏感,但不同种类及品种间存在一定差异。
3.2 重金属镉对植物PS Ⅱ光合能量转化的影响机制
3.2.1 重金属镉通过降低光合能量捕获、传递效率及PSⅡ 反应中心数量影响光合能量转化效率 有学者提出重金属镉引起PSⅡ激发能转化效率下降的原因可能是重金属镉通过抑制激发能向PSⅡ反应中心的传递,同时使部分PSⅡ反应中心数量减少,导致PSⅡ反应中心对激发能转化效率的下降[46]。但在另一项研究中发现镉胁迫下拟柱孢藻单位活性反应中心捕获的用于电子传递的能量(ET0∕RC)较非胁迫条件下下降8.33%,而单位活性反应中心RC 捕获的能量(TR0∕RC)增加6.04%[27],由此可以推测PSⅡ活性反应中心的数量及激发能捕获及传递效率的降低对于光合能量转化效率的影响主要表现为量的变化,这并不能解释镉胁迫下PSⅡ反应中心对激发能转化能力下降的质的变化。
3.2.2 重金属镉通过抑制光合电子传递降低PS Ⅱ光合能量转化效率 PSⅡ活性反应中心对激发能转化的实质是传递至PSⅡ光化学反应中心的激发能使P680 发生电荷分离产生阴阳离子自由基对(P680+·Pheo-)。Pheo-及P680+能否正常得失电子、P680及Pheo 结构的完整性是电荷分离能够正常且持续进行的关键,其中光合电子的得失构成了光合电子传递链,且尚没有证据表明重金属镉会直接引起P680及Pheo 结构的变化[47],因此光合电子传递过程受阻可能时是光合能量转化效率下降的主要原因。PSII 的光合电子传递易受到重金属镉的抑制[48,49],在一定浓度的镉胁迫下,水稻[38,39]、大豆[50]、菜豆[51]及龙葵[45]等植物PSⅡ光合电子传递速率均表现出不同程度的下降。
重金属镉分别通过影响PSⅡ受体侧及供体侧抑制光合电子的传递过程[36,47,52,53],重金属镉不同的作用位点对PSⅡ光化学反应中心存在不同的破坏机制。当重金属镉使P680受体侧电子传递受阻时,P680+·Pheo-发生电荷重组,产生三线态P680(3P680),随后3P680将电子传递给O2反应生成单线态氧1O2,而1O2通过直接攻击P680使PS Ⅱ光化学反应中心失去电荷分离能力,最终引起D1蛋白的裂解,导致光合能量转化效率下降[54]。而当重金属镉使P680供体侧电子传递受阻时,P680+无法接受电子,导致P680+积累,引起β-Car 和Chl a 等色素分子的氧化,并进一步导致D1 蛋白构象的变化及其自身水解,最终引起光合能量转化效率下降[54]。
3.2.3 重金属镉通过影响PS Ⅱ光化学反应中心结构降低PS Ⅱ光合能量转化效率 PS Ⅱ光化学反应中心由D1-D2-Cyt b559所组成,其结构的完整性也是影响光合能量转化的重要影响因素。其中D1、D2蛋白共同组成异源二聚体通过与P680、Pheo、质体醌(QA及QB)结合,构成了原初光反应和电子传递的关键场所。重金属镉能够直接或间接引起D1、D2蛋白含量的降低,如水稻、小麦等作物在镉胁迫下D1蛋白显著下降[55,56],部分水稻品种D2蛋白含量也受到镉胁迫的影响[55],但D1蛋白对重金属镉更为敏感。活性氧(ROS)在重金属镉引起D1、D2蛋白降解的过程中可能起重要作用。
Cyt b559 是一种完整的膜蛋白,由α、β 2 个亚基组成,在植物体内存在2 种电势态(高电势态和低电势态),Cyt b559 在PS Ⅱ光化学反应中心的生物合成及组装方面发挥重要作用[57]。有研究表明Cyt b559 从低电势态到高电势态的转变是放氧复合体组装到PS Ⅱ光化学反应中心的关键步骤之一[58],而重金属镉会通过Cd2+与Ca2+位点竞争结合等多种途径导致Cyt b559 由高电位向低电位形式转变[59,60],进而影响PS Ⅱ光化学反应中心结构的完整性。此外,也有研究发现Cyt b559 由高电势态转向低电势态时PS Ⅱ光化学反应中心放氧速率下降[61],虽然Cyt b559 不直接参与光合放氧过程,但可以通过高低电势态转化过程中蛋白构象的改变影响放氧复合物的活性及锰簇组装活化等过程[62]。
4 小结
重金属镉可以通过降低叶绿素含量、改变叶绿素的组成、破坏类囊体膜结构等途径影响植物原初光能转化的生理结构基础。此外,重金属镉会通过改变主体外周捕光复合体LHC Ⅱb 及内周捕光复合体(CP43、CP47)结构和功能来降低植物叶片光能捕获、传递效率,并通过抑制光合电子传递链及改变Cyt b559 电势态破坏PS Ⅱ光化学反应中心而降低植物对光能的转化效率。
类囊体膜的膜脂和蛋白质复合体的组成及分子结构在维持光合系统的超分子结构、原初光能转化中起重要作用,关于重金属镉对植物类囊体膜脂及蛋白质含量的影响已有较多的研究成果,但对构成膜脂和蛋白质的脂肪酸及氨基酸的变化研究较少,而这些小分子物质往往发挥某些重要的生理功能,如PG 分子中的trans-16:1。因此,重金属镉对脂肪酸及氨基酸等小分子代谢物质的影响及这些小分子物质如何影响PSⅡ-LHCⅡb 的组装及稳定性值得进一步的研究,其中重金属镉对LHCⅡb 三聚体结构的影响与光合能量的捕获与传递效率有直接联系,应该成为研究的重点课题。
重金属镉除了影响蛋白质含量以外,更多的影响可能来自于镉对蛋白质构象的改变,如重金属镉胁迫下D1蛋白的水解可能与蛋白质构象的变化密切相关,因此有必要从蛋白质水平上解析重金属镉对PSⅡ-LHCⅡb 中关键蛋白各级结构的影响。
植物对光合能量的捕获、传递及转化在很大程度上与PSⅡ-LHCⅡb 的分子空间结构、分子间距离及起分子间的相互作用密切相关。在光能捕获及传递过程中多涉及激发能在各色素分子间的传递(如Chl b 到Chl a、Chl a 到Chl a 及β-Car 到Chl a 等),即使在PSⅡ光化学反应中心,也存在各辅助色素分子与反应中心P680的能量传递,因此,进一步探究重金属镉对PSⅡ-LHCⅡb 分子空间结构的影响,可以从分子水平上探究镉胁迫对光合能量传递的影响。
在未来的研究中,将从代谢组、蛋白质结构及分子空间构型等水平,探究重金属镉对植物光合能量捕获、传递及转化过程的影响,阐明镉胁迫下植物光合作用的光物理特征及光化学反应的变化机制,能够为提高镉胁迫下植物的光能传递、转化效率、减少激发能损耗,提高光合效率提供理论依据。
