脂质对猪肠道健康影响的研究进展
2023-03-27侯伟晓张思雨郑鑫明
■侯伟晓 张思雨 郑鑫明 张 琳
(华南农业大学,广东广州 510642)
猪肠道作为吸收营养的主要部位,对仔猪和育肥猪的生长和发育尤为重要。仔猪早期胃肠道发育不成熟,容易受到感染,从而导致腹泻、生长缓慢、成活率下降,给畜牧业造成巨大损失。实际生产中广泛使用抗生素等化学试剂提高仔猪免疫力,但抗生素滥用导致许多问题,如猪肠道菌群紊乱、畜产品抗生素残留、养殖场环境污染等,限制了畜牧业的健康发展。
脂质包含的种类较多,多以健康、绿色为特性,如长、中、短链脂肪酸等属营养物质,植物精油等脂类衍生物属非营养物质。饲粮中添加中链脂肪酸可提高仔猪免疫力;添加植物精油可改善肠道健康,提高免疫力和调节肠道菌群结构。脂质由于残留少、毒副作用少、安全性高可作为功能型替抗产品,有益于猪肠道健康,促进生长发育。
本文介绍了脂质的分类及其在肠道内的代谢过程,重点阐述了脂质对肠道健康的作用,以期为脂质在猪生产中应用提供参考依据。
1 脂质及其分类
脂质是一类天然的大分子物质,包括脂肪、蜡、类固醇等。脂质被定义为疏水性或双亲性小分子。通常饮食中脂质的主要成分是三酰甘油(triacylglycerol,TG),TG 由一个甘油骨架连接三个脂肪酸分子组成,作为膳食脂肪中主要能量来源。膳食脂肪中还含有磷脂(phospholipids,PL)和一些少量的固醇,如胆固醇。磷脂在肠腔内主要成分是磷脂酰胆碱(phosphatidylcholine,PC)。食物中的固醇类主要为胆固醇(主要来自动物)和β-谷甾醇(主要来自植物)。胃肠道中的PL 和胆固醇不仅仅来自于食物,也来源于胆汁。除饮食中脂肪外,脂质还包括部分植物精油和植物提取物以及肠道菌群产物短链脂肪酸(short chain fatty acids,SCFAs)。植物提取物如肉桂醛、月桂酸等;SCFAs主要包括乙酸、丙酸、丁酸,主要由肠道中不易消化的单糖和淀粉多糖被微生物发酵所得;早期报道丁酸主要为结肠上皮细胞提供能量(Roediger 等,1982)。
2 脂质在肠道内的代谢过程
2.1 脂质在胃肠道内的消化吸收
胃肠道中可以消化吸收95%的膳食脂质,TG 在胃脂肪酶作用下继续分解为脂肪酸(fatty acids,FA)和二酰甘油(diacylglycerol,DAG)。食糜中的脂肪进入十二指肠后与胆汁酸和胰腺分泌的胰液混合消化,在胰脂肪酶的作用下分解为2-单酰甘油(2-monoacyglycerol,2-MAG)和FA;膳食磷脂被胰腺磷脂酶A2(pancreatic phospholipase,PLA2)消化分解为溶血磷脂酰胆碱(lysophosphatidylcholine,LPC)和FA;胆固醇被羧基酯水解酶酶解释放游离胆固醇和FA;游离的FA、PC、2-MAG 和胆固醇被整合到脂质微粒上,在小肠肠腔中被吸收到肠上皮细胞内。
2.2 脂质在肠上皮细胞中的转运
FA和其他脂质成分都通过被动运输和主动运输被肠上皮细胞吸收,脂质成分通过磷脂双分子层进入肠上皮细胞。白细胞分化抗原36(cluster of differentiation 36,CD36)在肠上皮细胞中有较高表达(Lobo等,2001),并能影响乳糜微粒的释放,是FA 主要转运载体之一(Nassir等,2007)。脂肪酸转运蛋白家族4(fatty acid transport protein 4,FATP4)在转运FA 过程中发挥重要作用,是肠上皮细胞摄取脂肪酸的主要转运载体(Stahl等,1999)。
SCFAs 主要由单羧酸转运载体(monocarboxylate transporters,MCTs)和钠依赖性单羧酸转运载体1(sodium-dependent monocarboxylate transporter 1,SMCT1)或被动扩散进入结肠上皮细胞(Stumpff等,2018)。
尼曼-皮克C1 型类似蛋白-1(niemann-pick-c1-like 1,NPC1L1)在介导胆固醇的吸收中发挥重要作用(Altmann 等,2004),三磷酸腺苷结合转运蛋白(adenosine triphosphate-binding cassette transporter,ABC)中ABCG5和ABCG8作为胆固醇外排转运体负责胆固醇的排出(Lee 等,2001;Berge 等,2000;Lu 等,2001)。B 类Ⅰ型清道夫受体(scavenger receptor class B typeⅠ,SR-BⅠ)在肠道中有促进胆固醇摄取的作用(Bietrix等,2006)。
2.3 脂质在肠上皮细胞内的代谢
在肠上皮细胞内,单酰甘油(MAG)和长链脂肪酸(long chain fatty acids,LCFA)在内质网(endoplasmic reticulum,ER)膜上重新合成TG(Kayden等,1967),再形成乳糜微粒进入淋巴循环中,供给机体组织脂肪酸。中链脂肪酸(medium chain fatty acid,MCFA)与白蛋白结合进入门静脉循环,在肝脏内直接氧化供能或重新形成极低密度脂蛋白(very-low-density lipoprotein,VLDL)(Babayan,1987)。SCFAs 则进入结肠上皮细胞的柠檬酸循环产生能量(Schönfeld 等,2016),其余的SCFAs则进入到肝门静脉循环中。
肠上皮细胞内TG 合成主要通过MAG 通路,即MAG 和脂酰辅酶A 在酰基转移酶(monoacylglycerol acyltransferases,MGATs)作 用 下 形 成DAG(Coleman等,1986;Yen 等,2008),随后DAG 在二酰甘油酰基转移酶(diacylglycerol acyltransferase, DGAT)作用下合成TG;还可通过甘油磷脂(glycerol-3-phosphate,G3P)途径合成DAG,进而合成TG;TG 以脂滴形式储存在肠上皮细胞内。肠上皮细胞内游离的胆固醇被酰基辅酶A 胆固醇酰基转移酶(acyl-coenzyme A:cholesterol acyltransferases,ACAT)酯化为胆固醇酯,随后被纳入乳糜微粒核心(Nguyen 等,2012)。胞内LPC 被溶血磷脂酰胆碱酰基转移酶(lysophosphatidylcholine acyltransferase,LPCAT)酯化为PC,LPCAT3 在乳糜微粒分泌中起关键作用(Rong等,2015)。
3 脂质对猪肠道健康的作用
脂质在猪肠道健康方向发挥着诸多作用:调控采食和内分泌,促进肠道发育,提高消化吸收能力和免疫力;改善肠道菌群结构和降低氧化应激损伤,其中关于短链脂肪酸中丁酸的研究较多;植物提取物中月桂酸、牛至属脂类中萜类衍生物具有良好的抗菌能力,可作为饲料中替抗物使用(Lillehoj 等,2018)。脂质对肠道有益调控作用见表1。
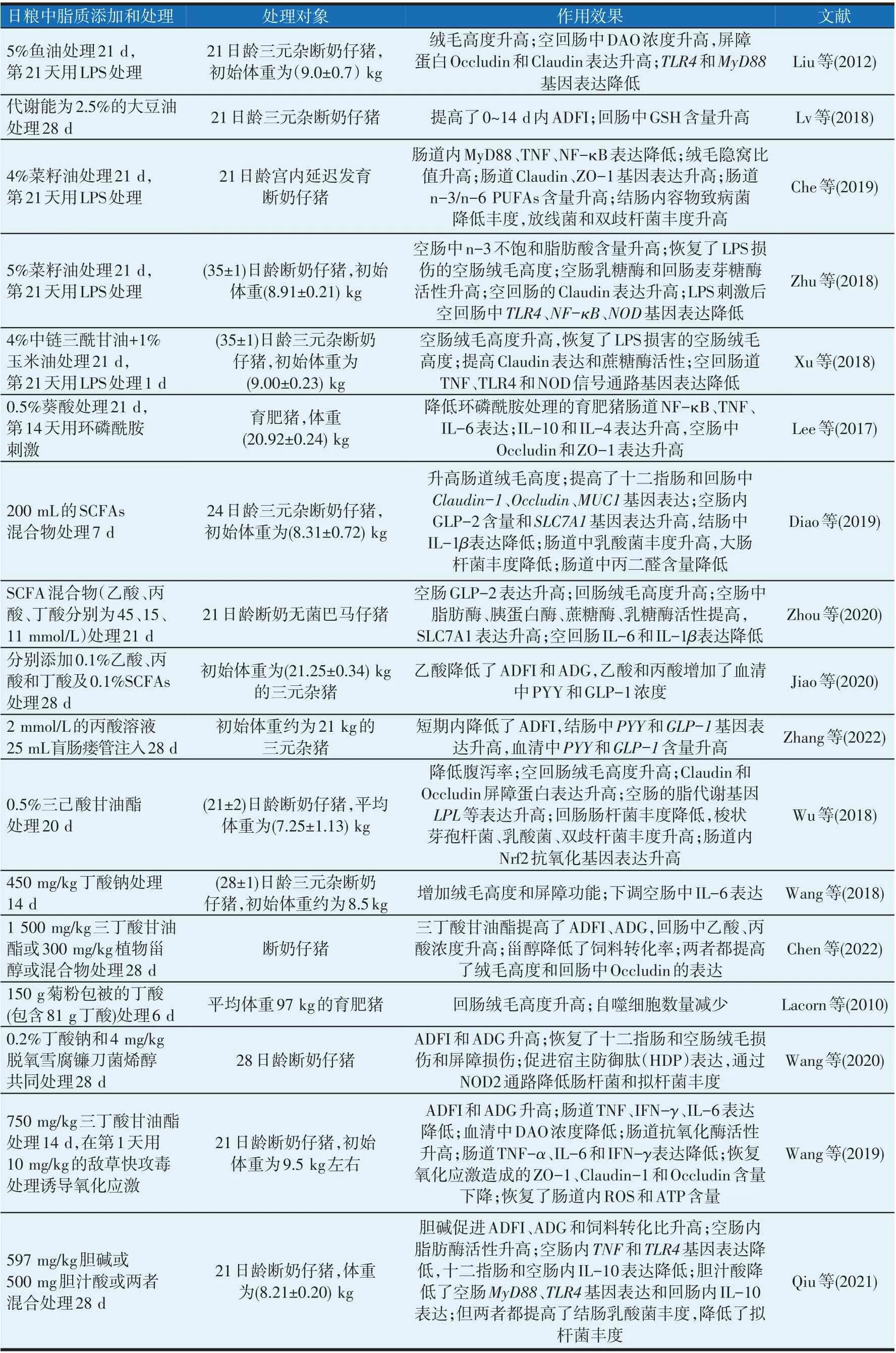
表1 脂质对猪肠道作用
3.1 脂质对肠道发育的影响
肠上皮细胞的两个主要功能是吸收营养和防御外界环境干扰,肠上皮细胞可分泌消化酶,表面有微绒毛结构,即刷状缘。在空肠和回肠肠段中,肠上皮组成凸状绒毛隐窝结构,绒毛向肠腔内凸出,扩大营养物质吸收面积,肠绒毛上覆盖单层的肠上皮细胞,肠上皮细胞具有快速更新优势,4~5 d 内即可完全更新一次(Johnson,1988;Williamson,1978)。通常以肠绒毛高度、隐窝深度以及绒毛高度与隐窝深度比值作为肠道形态发育指标。
n-3 多不饱和脂肪酸(n-3 polyunsaturated fatty acids,n-3 PUFAs)和单不饱和脂肪酸有益于猪肠道发育。饲料中添加100 g/kg 鱼油处理母猪26 d,可提高后代断奶仔猪回肠的绒毛高度与隐窝深度比值(Leonard 等,2011)。鱼油含40%的二十碳五烯酸(eicosapentaenoic acid,EPA)和25%的二十二碳六烯酸(docosahexaenoic acid,DHA)。Liu等(2012)研究也发现,含5%鱼油的饲粮处理断奶仔猪21 d,显著提高空肠和回肠中绒毛高度。
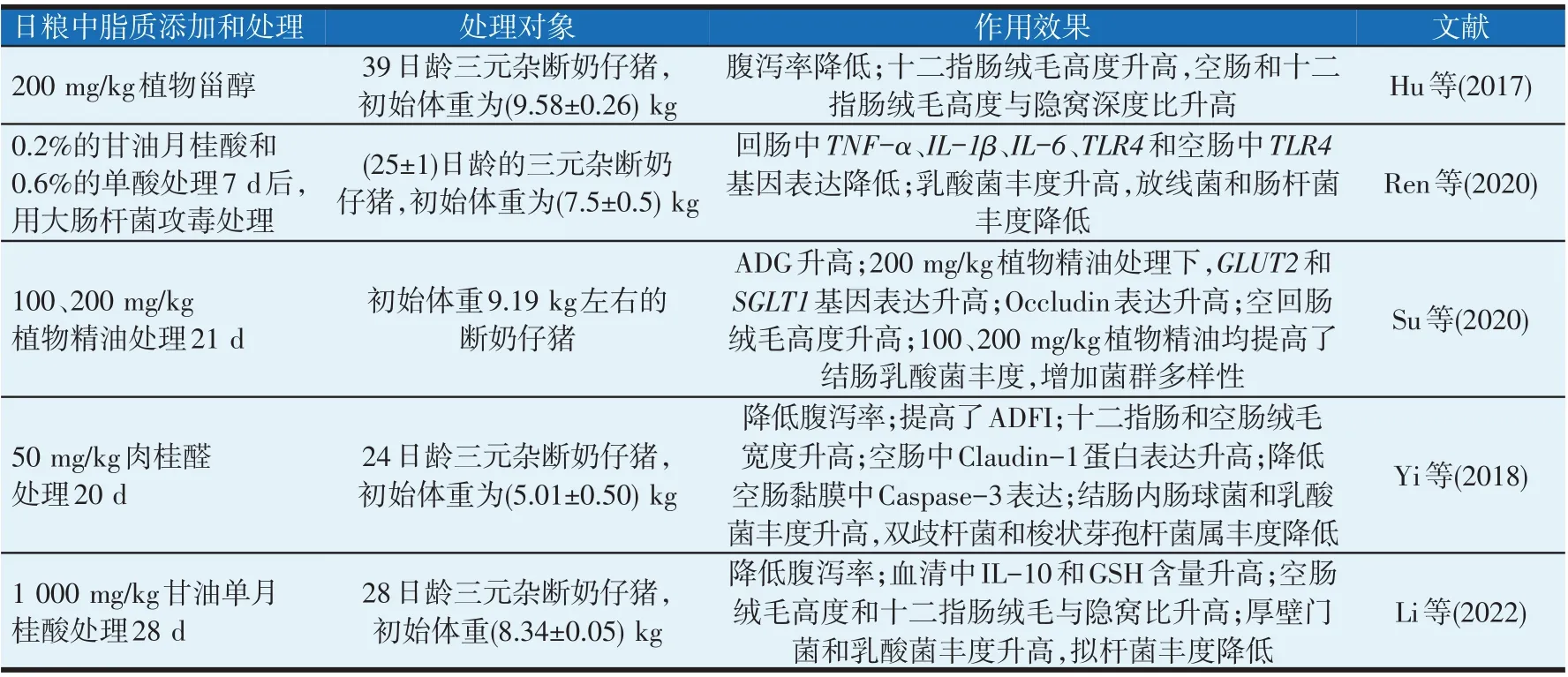
表1(续)脂质对猪肠道作用
MCFA 和SCFAs 能保护肠道绒毛结构。日粮中分别添加0.175%(De Keyser 等,2019)和4%(Xu 等,2018)的中链三酰甘油饲喂不同日龄断奶仔猪21 d后可恢复受脂多糖(lipopolysaccharide,LPS)损伤后的肠绒毛高度。有研究给断奶仔猪灌胃200 mL 的SCFAs(含乙酸40.08 mmol/L、丙酸15.41 mmol/L 和丁酸9.78 mmol/L)持续7 d,显著提高了十二指肠、空肠和回肠的绒毛高度(Diao 等,2019);然而在给无菌断奶仔猪口服25 mL/kg体重的SCFAs(乙酸、丙酸、丁酸分别45、15、11 mmol/L)21 d 却仅有增加回肠绒毛高度的趋势(Zhou 等2020),可能是受肠道菌群结构的影响所致。
丁酸可提高猪肠道绒毛高度。如日粮添加0.2%的丁酸钠饲喂27 kg左右的育肥猪70 d,可提高肠道绒毛高度(Wang等,2020);在28日龄断奶仔猪日粮中添加450 mg/kg 丁酸钠处理2 周,可提高肠道绒毛高度(Wang等,2019);用含290 g/kg丁酸的日粮饲喂75 kg左右的育肥三元杂猪5 d,空肠绒毛高度升高(Claus等,2007);日粮中添加150 g/d 菊粉(含81 g 丁酸)饲喂育肥猪(约97 kg)可促进回肠绒毛高度增加(Lacorn等,2010);此外,有研究表明0.2~0.4 mmol/L的丁酸钠可促进IPEC-J2细胞的增殖(Yang等,2020)。
植物精油中萜类等物质有益于肠道发育。如饲粮中添加中0.2%的SCFAs 和有机酸混合物饲喂断奶仔猪42 d 后可提高回肠绒毛高度(Li 等,2018);用含50 mg/kg肉桂精油的日粮处理断奶仔猪20 d,提高了肠道绒毛高度(Yi等,2018);日粮中单独添加甲酸、月桂酸、牛至油、植物甾醇同样有促进猪肠道发育的效果(Ren等,2020;Hu等,2017)。
3.2 脂质对猪采食调控和肠道内分泌代谢功能的影响
肠道作为摄入能量的主要器官,肠道中存在分泌型细胞如肠分泌细胞、杯状细胞和潘氏细胞。肠分泌细胞分泌胆囊收缩素(cholecystokinin,CCK)、胰高血糖素样肽(glucagon-like peptide,GLP)和胃肠肽类激素酪酪肽(peptide YY,PYY)(Gribble 等,2016),PYY和GLP1 可调控动物的食欲,GLP-2 则可促进肠道发育(Steinert 等,2017),脂肪酸营养可通过调控肠道分泌和促进猪肠道发育增加ADFI。
日粮中添加2.5%的代谢能大豆油(富含PUFA)处理断奶仔猪28 d,与添加2.5%的代谢能谷氨酰胺相比,在14 d处理期内明显提高ADFI、ADG及饲料转化率(Lv等,2018),提示饲粮中油脂气味影响采食。
SCFAs 具有刺激性气味,可调控食欲,并能提高猪空肠中GLP-2 的表达。用200 mL 的SCFAs(乙酸40.08 mmol/L、丙酸15.41 mmol/L、丁酸9.78 mmol/L)灌胃初始体重为(8.31±0.06)kg的断奶仔猪7 d,降低了ADFI 和血清瘦素含量(Jiao 等,2018);给断奶仔猪口服SCFAs处理21 d后,提高了空肠中GLP-2的表达(Zhou 等,2020)。饲粮中添加混合0.1%的SCFAs(乙酸、丙酸、丁酸比例为3∶1∶1)处理初始体重为(21.25±0.34)kg 的三元杂猪28 d,血清中PYY 和GLP-1 的含量升高,结肠中PYY 的mRNA 表达增加,ADFI和ADG降低(Jiao等,2020),仅添加0.1%的丙酸饲粮也提高了血清中PYY 和GLP-1 的含量及结肠中PYY 的mRNA 表达,但没有影响猪的采食;另有研究在体重为16 kg 左右的三元杂猪盲肠瘘管中注入2 mmol/L 的丙酸钠25 mL,持续28 d,发现丙酸仅能短期降低ADFI,血清中PYY和GLP-1含量升高,结肠中PYY 表达升高(Zhang 等,2022),这项研究补充了丙酸对采食的影响。
丁酸通过调控PYY 和促进肠道发育进而提高猪的食欲。如日粮中添加1 500 mg/kg三丁酸甘油酯处理断奶仔猪28 d,提高了ADFI 和ADG(Chen 等,2022);用含0.2%丁酸钠的日粮饲喂育肥猪70 d,提高了ADFI 和ADG,并降低了饲料转化比(Sun 等,2020);在体重为28.7~41.9 kg 的育肥猪饮食前1 h 口服含0.01 mol/L 丁酸钠的饮水持续28 d,很少影响育肥猪的采食量和肠道内分泌功能(Val-Laillet 等,2018)。日粮中添加0.2%的丁酸钠和0.75%的三丁酸甘油酯分别对应处理脱氧雪腐镰刀菌刺激和氧化应激状态下的断奶仔猪,有提高ADFI 作用(Wang 等,2020;Wang 等,2019)。丁酸钠还降低了脱氧雪腐镰刀菌攻毒仔猪血清中PYY 含量,恢复了ADFI。丁酸直接充当肠上皮细胞的能量来源,促进了肠道发育,可能是因为提高了酶活性进而增加了ADFI。
饲粮中添加597 mg/kg 胆碱饲喂断奶仔猪28 d,提高了ADFI、ADG和饲料利用率,肠道内脂质消化能力提高(Qiu等,2021)。此外还有研究表明,饲喂添加橄榄油提取物500 mg/kg的日粮可恢复LPS刺激断奶仔猪下降的ADFI(Liehr等,2017),用含50 mg/kg肉桂精油的饲粮处理断奶仔猪20 d,提高了ADFI(Yi 等,2018),可能与下丘脑神经肽Y 的分泌有关。关于脂质如何响应肠脑轴调控饮食还需有更多研究。
3.3 脂质对猪肠道营养吸收的影响
断奶应激和肠道疾病会造成仔猪肠道内活性酶下降,减少肠道对营养物质的吸收,影响到仔猪的生长状况和免疫力。脂质可提高肠道内消化酶的活性,增加机体能量吸收利用。
PUFAs 和MCFA 均可提高断奶仔猪肠道中消化酶活性。日粮添加5%的亚麻籽油饲喂断奶仔猪21 d,可提高空肠乳糖酶和回肠麦芽糖酶的活性(Zhu 等,2018);添加4%的中链三酰甘油饲喂断奶仔猪21 d,空回肠中蔗糖酶和麦芽糖酶活性升高(Xu等,2018);用含中链三酰甘油的日粮处理断奶仔猪14 d,可提高空回肠中脂肪酶活性(Cui等,2022)。
SCFAs 可提高猪肠道的消化吸收能力。Zhou 等(2020)选用无菌巴马猪,口服灌胃断奶仔猪SCFAs(乙酸、丙酸、丁酸分别45、15、11 mmol/L),持续21 d,可提高空肠内胰脂肪酶、乳糖酶和蔗糖酶活性和氨基酸转运蛋白溶质载体家族7成员1(solute carrier family7 member1,SLC7A1)的基因表达,改善小肠内氨基酸稳态;给初始体重为(8.31±0.72)kg 的断奶仔猪口服灌胃200 mL 的SCFAs 持续7 d,提高了空肠内SLC7A1的基因表达(Diao等,2019)。提示SCFA可提高猪肠道氨基酸利用能力。
丁酸可提高猪肠道吸收能力和肠道内糖脂转运代谢(Claus 等,2007)。饲粮中添加0.5%的三己酸甘油酯处理断奶仔猪21 d,可促进空肠中脂蛋白脂肪酶(lipoprotein lipase,LPL)、过氧化物酶体增殖激活受体(peroxisome proliferator-activated receptor gamma,PPARG)、磷酸烯醇丙酮酸羧基酶1(phosphoenolpyruvate carboxykinase 1,PCK1)基因表达,降低脂肪酸合酶(fatty acid synthase,FASN)表达,影响肠道内糖脂代谢和吸收(Wu等,2018)。
日粮中添加胆碱597 mg/kg 处理断奶仔猪28 d,提高了空肠内脂肪酶活性(Qiu等,2021)。用含固醇类的乳汁饲喂7日龄的仔猪21 d,提高了仔猪回肠食糜中胆固醇的消化率(Babawale等,2018)。
植物精油中月桂酸和百里香酚可提高猪肠道消化功能。日粮添加1 000 mg/kg甘油单月桂酸处理断奶仔猪28 d,粗脂肪、粗蛋白、干物质的肠道表观消化率升高(Li 等,2022);用含200 mg/kg 植物精油(13.5%的百里香酚和4.5%的肉桂醛)的日粮处理断奶仔猪21 d,空肠内蔗糖酶和乳糖酶活性提高,钠-葡萄糖转运体2(glucose transporter 2,GLUT2)和钠-葡萄糖转运载体1(sodium/glucose cotransporter 1,SGLT1)基因表达升高(Su 等,2020);用添加甘油单月桂酸的日粮处理断奶仔猪14 d,空肠和回肠中脂肪酶活性升高(Cui等,2022)。
3.4 脂质对猪肠道屏障功能的影响
肠上皮细胞之间形成的物理屏障主要由细胞间的紧密连接蛋白和黏着小带构成,屏障蛋白包括紧密连接蛋白(zonula occludens-1,ZO-1)、Occludin 和Claudin;α-catenin 和β-catenin 则 形 成 黏 着 小 带(Buckley等,2018)。G氏细胞和潘氏细胞分泌黏蛋白(mucin,MMU)、抗菌肽(antimicrobial protein,AMP)和三叶因子3(trefoil factor 3,TFF3)等有助于肠道化学屏障形成(Johansson等,2016)。并且仔猪断奶后肠道中ZO-1和Claudin-1表达明显下降(Wang等,2016);肠上皮屏障在防御细菌和病原体侵入时可起到对肠道保护的作用。
PUFAs有益于保持肠道屏障功能。Liu等(2012)研究发现,饲粮中添加5%的鱼油处理断奶仔猪21 d,显著提高了屏障蛋白Occludin 和Claudin-1 的表达。随后发现含5%亚麻籽油的饲粮也可提高断奶仔猪空回肠中Claudin 蛋白表达(Zhu 等,2018)。12.5 μg/mL的DHA 能保护IPEC-J1 细胞屏障功能完整性(Xiao等,2020),可能与Caspase3/8通路抑制炎症有关。
MCFA 有益于肠道屏障功能。饲粮中添加4%中链三酰甘油处理断奶仔猪21 d,屏障蛋白Occludin的表达升高(Xu 等,2018)。并且1 mmol/L 辛酸和0.5 mmol/L 壬酸可通过调控组蛋白去乙酰化酶(HDAC)活性提高IPEC-J2 细胞中防御素(beta-defensin,pBD)蛋白表达(Wang等,2018)。
短链脂肪酸可提高猪回肠中Occludin、Claudin表达(Diao等,2017)。口服灌胃断奶仔猪200 mL的SCFAs持续7 d,十二指肠和回肠内Occludin 和Claudin-1的基因表达升高(Diao等,2019)。
丁酸可通过丝裂原激活蛋白激酶类(mitogen-activated protein kinases,MAPK)和丝氨酸/苏氨酸激酶 一磷酸腺苷活化蛋白激酶(serine/threonine kinase adenosine monophosphate-activated protein kinase,AMPK)通路增强猪肠道屏障功能。丁酸可能通过核苷酸结合寡聚化结构域蛋白2(nucleotide-binding oligomerization,NOD2)通路调节防御肽表达(Wang等,2020);用含0.2%丁酸的日粮饲喂断奶仔猪10 d,可通过抑制组蛋白去乙酰化酶活性,提高受大肠杆菌感染仔猪的回肠pBD2和pBD3蛋白表达(Xiong等,2016)。另有研究表明,丁酸在猪肠道上皮细胞IPEC-J2 中调控Toll样受体2(toll-like receptors,TLR2)和表皮生长因子受体(epidermal growth factor receptor,EFGR)的表达,进而提高pBD2 和pBD3 表达(Dou 等,2020),与HDAC的表达无关。丁酸可通过核因子kappa B(nuclear factor kappa B,NF-κB)和MAPK 通路提高猪空肠中ZO-1、Occludin、Claudin蛋白表达(Tian等,2022)。在细胞水平上,1 mmol/L丁酸钠在过氧化氢诱导损伤的IPEC-J2 细胞中通过AMPK 通路恢复ZO-1、Occludin和Claudin 蛋白表达(Li 等,2022)。丁酸调控猪肠道屏障功能的内在机制还需深入研究。
植物精油类中肉桂醛、百里香酚、月桂酸、甲酸等都能保护猪肠道的屏障功能。日粮中添加0.6%的甘油月桂酸和0.2%的甲酸混合饲喂断奶仔猪,空肠Claudin基因表达升高(Ren 等,2020);饲粮中添加植物精油(13.5%的百里香酚和4.5%的肉桂醛)饲喂断奶仔猪21 d,回肠中Occludin 蛋白表达升高(Su 等,2020)。Pu 等(2018)报道,用含3 000 mg/kg 苯甲酸、400 mg/kg 凝结芽孢杆菌和400 mg/kg 牛至油的饲粮处理初始体重为(7.64±0.46)kg 的断奶仔猪26 d 后,可恢复接种产毒素大肠杆菌刺激后的仔猪肠道ZO-1、Occludin 和MMU 的表达量。植物精油对猪肠道屏障的有益作用可能与其本身的抗菌效应有关,深层次分子机制还需再挖掘。
3.5 脂质对猪肠道免疫功能的影响
肠道长期暴露于食物中抗原、微生物以及病原体中,拥有先天免疫系统和适应性免疫系统,用于防御微生物和病原体等的感染。免疫反应发生的场所包括肠上皮细胞层、肠道固有层、黏膜下层,这些部位都包含大量派尔集合淋巴结,由上而下形成完整的检测保护系统。肠上皮细胞层通常作为主要发生免疫反应的场所,肠上皮细胞拥有先天免疫机制形成保护网络,通过模式识别受体(pattern recognition receptors,PRRs)识别入侵的微生物和病原体,启动免疫反应,保护肠道和肠道微生物免受伤害(Takeuchi 等,2010)。PRRs 包括Toll 样受体(toll-like receptors,TLRs)、核苷酸寡聚受体(nucleotide-binding oligomerization-like receptors,NLRs)和骨髓分化初级反应蛋白(myeloid differentiation primary response protein,MyD88)等。此外,肠道微生物及其产物SCFAs 在肠道免疫功能上已有报道,LCFA 在免疫方面研究也有报道(Haghikia 等,2015),但在猪肠道上的研究还未系统整理。
PUFAs 可通过TLR4 和NOD2 通路降低断奶仔猪的炎症因子表达。饲粮中添加4%亚麻籽油处理宫内生长受限的断奶仔猪,降低了肠道内MyD88、肿瘤坏死因子(tumor necrosis factor,TNF)和白介素-10(interleukin,IL-10)基因表达(Che 等,2019);用含5%亚麻籽油的日粮处理断奶仔猪,可通过TLR4 和NOD2通路可降低LPS 攻毒后肠道内MyD88 和NF-κB 表达(Zhu 等,2018)。12.5 μg/mL 的DHA 可降低IPEC-J1细胞内Caspase3 和Caspase8 的表达(Xiao 等,2020)。提示PUFAs可提高猪肠道免疫力。
MCFA 可提高猪肠道免疫能力。用含4%中链三酰甘油的饲粮处理断奶仔猪,再用LPS 处理后,降低了断奶仔猪空肠和回肠中TLR4、NOD2的基因表达(Xu 等,2018),有效减弱LPS 引起的肠道炎症反应。在体外试验中,用0.5 mmol/L 葵酸处理IPEC-J2 细胞24 h,降低了环磷酰胺刺激后IPEC-J2细胞内TNF-α、NF-κB、干扰素γ(interferon,IFN-γ)的基因表达(Lee等,2017)。提示MCFA 提高猪肠道免疫力可能依赖于NF-κB信号通路。
SCFAs 也可提高猪肠道免疫能力。口服灌胃断奶仔猪200 mL的SCFAs 处理7 d,结肠中白细胞介素1β(interleukin beta,IL-1β)表达降低(Diao 等,2019);Zhou 等(2020)对无菌断奶仔猪口服灌胃SCFAs 处理21 d,空回肠中IL-1β、白细胞介素6(interleukin,IL-6)表达降低;用含丙酸钠饮食处理初始重为20 kg 左右的三元杂猪28 d,结肠中NF-κB 和白细胞介素18(interleukin,IL-18)表达升高(Zhang 等,2018)。提示SCFAs有助于维持肠道免疫稳态。
丁酸在猪肠道免疫功能的发挥中具有重要作用。Xu等(2016)给刚出生仔猪口服丁酸钠7 d,回肠中IL-6、白细胞介素8(interleukin-8, IL-8)的表达降低,转化生长因子(transforming growth factor-β,TGF-β)和IL-10 表达升高;丁酸钠能有效降低断奶仔猪肠道中TNF-α和IL-6的表达(Wang等,2018;Wang等,2019;Tian等,2022),表明丁酸钠在仔猪肠道免疫中发挥重要作用。丁酸还可通过抑制NF-κB/MAPK 通路基因实现抗菌效果(Tian等,2022)。
固醇类物质也有抗肠道炎症效果。饲粮中添加200 mg/kg的植物甾醇降低了断奶仔猪十二指肠中肠道黏膜内分泌型免疫球蛋白A(secretory immunoglobulin A,sIgA)浓度,增强了免疫能力(Hu等,2017)。
有机酸和植物精油在猪的短、中、长期处理中表现出改善肠道免疫功能的作用。Ren等(2020)发现饲粮中添加0.6%的单酸和0.2%的甘油单月桂酸酯处理断奶仔猪7 d后,减轻了大肠杆菌引起的肠道炎症,通过NF-κB p65 和MAPK p38 的磷酸化通路降低了回肠中TNF-α、IL-1β、IL-6和TLR4的基因表达;用含苯甲酸、凝结芽孢杆菌、牛至油混合物的日粮处理断奶仔猪,降低了产肠毒素大肠杆菌攻毒后仔猪肠道中TLR4、NOD2、TNF-α、IL-1β的基因表达,提高了内分泌型免疫球蛋白含量(Pu等,2018)。
3.6 脂质对肠道菌群结构的影响
肠道菌群对宿主健康有许多益处,如发酵不易消化的碳水化合物产生SCFAs,为宿主提供能量,参与肠道糖异生过程(De Vadder等,2014);影响肠道中营养物质的消化代谢(Sonnenburg 等,2016);参与黏膜免疫的成熟过程(Thaiss 等,2016)。仔猪肠道菌群经常受到母体遗传、饮食、环境的影响,仔猪的肠道菌群在哺乳期由母乳塑造成一个以乳酸菌为主导的稳定状态(Frese 等,2015),创造一个厌氧环境,有利于拟杆菌、乳酸菌、双歧杆菌、梭状芽孢杆菌的生长(Petri等,2010)。断奶期会造成肠道菌群失调和菌群多样性减少;通常肠道菌群失调的标志是专性厌氧菌的减少,如乳酸杆菌减少和菌群多样性降低。肠道菌群多样性减少有助于细菌的生长。
PUFAs有益于猪肠道内菌群多样性增加、有益菌丰度提高、有害菌丰度降低。在仔猪断奶前饲喂脂质混合物、发酵乳杆菌,可增加肠道中梭状芽孢杆菌的丰度,提高盲肠中SCFAs 浓度水平,改善肠道分泌功能和调节稳态(Lemaire等,2018)。有报道表明,高油酸饮食可提高猪肠道中普氏菌属丰度(López-García等,2021)。饲粮中添加6%的棕榈油处理断奶仔猪28 d,可降低十二指肠和空肠中厚壁菌门丰度,提高小肠中放线菌属丰度,降低肠道对脂肪吸收,而添加7.5%的封装棕榈油(含80%棕榈油)可降低小肠中放线菌属丰度,提高肠道对脂肪的吸收(Yang 等,2020)。提示不同脂肪形式可通过菌群影响肠道吸收能力。
SCFAs有益于肠道菌群稳定,提升乳酸菌等有益菌丰度,降低大肠杆菌等有害菌丰度。口服灌胃断奶仔猪200 mL的SCFAs持续7 d,提高了回肠和结肠食糜中乳酸菌丰度,降低了大肠杆菌丰度(Diao等,2019),提高了消化能力;育肥猪通过瘘管注射300 mL混合SCFAs(乙酸、丙酸、丁酸分别61.84、18.62、12.55 mmol/L)处理14 d,回肠中芽孢杆菌、乳酸杆菌和双歧杆菌丰度升高,大肠杆菌丰度降低,并降低了肠道中的pH,抑制了肠道中病原体和细菌的生长(Diao等,2017);Zhang等(2018)报道,用丙酸钠(2 mol/L,pH 为5.8)通过瘘管处理育肥猪28 d,拟杆菌属和普氏菌属丰度升高,结肠中厚壁菌门丰度降低。以上结果表明SCFAs 处理有益于健康菌群生长并稳定肠道的微生物系统。
磷脂类可提高猪肠道中有益菌群丰度。饲粮中分别添加胆碱597 mg/kg 和胆汁酸500 mg/kg 处理断奶仔猪28 d,提高了结肠丁酸浓度(分别提高29.1%、20.9%)和乳酸菌属丰度(分别提高42.3%、39.7%)(Qiu 等,2021)。用含5%的溶血磷脂饲粮处理母猪,处理时间从妊娠期至产仔后18 d,增加了后代仔猪空肠中厚壁菌门与拟杆菌门的比例(1.0∶3.5),肠道免疫能力提高,有益菌的丰度升高(Jang等,2020);
有机酸和植物精油类物质可保持肠道菌群健康,促进SCFA 产生,增加菌群多样性和有益菌群稳态。Li 等(2018)研究发现,用含有机酸(0.2%丁酸、SCFAs、MCFA、有机酸、酚类的混合物)的饲粮处理断奶仔猪21 d,可明显提高普氏菌属丰度,提高肠道的免疫稳态和肠道碳水化合物消化能力。然而单独的有机酸处理是否有此效果还有待探讨。Ren 等(2020)报道,饲粮中添加0.6%的单酸和0.2%的甘油单月桂酸处理断奶仔猪7 d,回肠中淀粉乳杆菌属和唾液乳杆菌属的相对丰度升高,放线杆菌属的丰度降低,有益于猪肠道的免疫稳态。含1 000 mg/kg甘油月桂酸的日粮也可提高断奶仔猪肠道中厚壁菌门与拟杆菌门比例,提高乳酸菌丰度(Li等,2022)。
饲粮中脂质不仅有益于猪肠道功能,同时可改变其菌群结构,肠道菌群与肠道功能又密切相关,脂质是否是通过改变猪肠道菌群而影响肠道功能,哪些菌群直接影响肠道功能或直接作用于肠道功能,还需进一步研究确定。
3.7 脂质对肠道氧化应激的影响
氧化应激通常与多种疾病有关,肠道氧化应激与仔猪断奶时的肠道屏障功能损伤、消化吸收能力减弱密切相关(Wang 等,2016)。氧化应激由两种机制(Forman 等,2021)产生,第一种是活性氧(reactive oxygen species,ROS)的产生,如羟自由基(·OH)、过氧亚硝基阴离子(ONOO-)和次氯酸(HOCl)等,可直接氧化大分子物质,如氧化脂质、结构蛋白、酶和核酸,导致细胞功能异常和死亡;第二种是氧化还原信号通路紊乱。机体依赖于抗氧化酶提供底物,消除活性氧和修复氧化应激损伤提高抗氧化能力。抗氧化酶包括超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GPX-Px)、谷氨酸-半胱氨酸连接酶(glutamate-cysteine ligase,GCL)、谷氨酸-半胱氨酸连接酶合成酶催化亚基(glutamate-cysteine ligase synthetase catalytic subunit,GLCL)和硫还原蛋白(thioredoxin,TRX)等;核因子E2相关因子(nuclear factor E2-related factor,Nrf2)与亲电反应元件调控抗氧化应激酶产生和抵抗氧化应激(Pickering等,2012)。
长中短链脂肪酸均可降低猪肠道氧化应激状态。用含大豆油(占代谢能2.5%)的饲料处理断奶仔猪28 d,可提高回肠中谷胱甘肽(glutathione,GSH)含量(Lv等,2018);分别用12.5 μg/mL的EPA和DHA对脱氧雪腐镰刀菌烯醇攻毒的猪肠上皮细胞处理24 h,可降低细胞内ROS 含量(Xiao 等,2020);0.5 mmol/L葵酸处理环磷酰胺攻毒的IPEC-J2细胞24 h,降低了细胞内过氧化氢和丙二醛含量和GLCL基因表达(Lee等,2017),丙二醛为脂质过氧化产物。口服灌胃断奶仔猪200 mL的SCFAs持续7 d,空肠和结肠中丙二醛含量降低(Diao等,2019);饲粮中添加750 mg/kg三丁酸甘油酯处理氧化应激状态的断奶仔猪14 d,仔猪肠黏膜内SOD 含量和GPX-1、GPX-4基因表达升高,丙二醛和ROS含量降低(Wang等,2019);1 mmol/L丁酸钠可通过AMPK 通路降低IPEC-J2细胞的ROS含量,抵抗氧化应激(Li等,2022)。
植物精油类中的甘油月桂酸和百里香酚可提高猪的氧化应激能力。用含200 mg/kg植物精油(13.5%的百里香酚+4.5%的肉桂醛)的饲粮处理断奶仔猪14 d,空回肠内GSH 含量升高,丙二醛浓度降低(Su 等,2018),并且50 μmol/L 的百里香酚处理IPEC-J2细胞1 h 降低了LPS 刺激后的IPEC-J2 细胞内ROS 含量(Omonijo 等,2019);2.5~10 μg/mL剂量的牛至油可通过Nrf2 通路降低过氧化氢诱导应激损伤的IPEC-J2细胞内ROS 和丙二醛含量,提高了SOD的基因表达(Zou等,2016),有效减弱肠道氧化应激。
4 总结展望
脂质具有促进消化吸收、保护肠道屏障、提高肠道免疫力、调节肠道菌群结构等功能,有益于肠道健康,在猪养殖业中具有极大的应用前景,可用于饲料添加剂和抗生素应用的替代。然而,目前关于脂质的研究仍存在不足之处,如脂质在肠道中的作用位点、作用机制尚不明确;不同种类、不同成分的脂质之间可能存在相互协同作用;脂质的添加剂量、脂质对不同阶段猪的影响、植物精油的代谢途径等问题还需进一步深入研究证实。在精准营养调控方面,如何有效利用脂质维护肠道健康也将是未来的挑战。
