Regulation of Reproduction in Delayed Gametophyte of Saccharina japonica (Laminariales, Phaeophyceae):Effects of Light Intensity, Quality and Photoperiod
2023-03-17WANGXiaoLIANGZhouruiLIUFuliZHANGPengyanYUANYanminLIUYiWANGWenjunandSUNXiutao
WANG Xiao, LIANG Zhourui, LIU Fuli, ZHANG PengyanYUAN Yanmin,LIU YiWANG Wenjunand SUN Xiutao
Regulation of Reproduction in Delayed Gametophyte of(Laminariales, Phaeophyceae):Effects of Light Intensity, Quality and Photoperiod
WANG Xiao1), #, LIANG Zhourui1),#, LIU Fuli2), *, ZHANG Pengyan1), YUAN Yanmin1),LIU Yi1), WANG Wenjun1), and SUN Xiutao1)
1),,,,266071,2),,,,266003,
gametophytes can survive a long period under unfavorable environmental conditions, while they also delay in growth and/or reproduction. Although the reproduction in delayed gametophyte ofwas known to be strong- ly influenced by light intensity, light quality, and photoperiod, no previous studies have evaluated their interactive effects on game- togenesis. To evaluate these effects, we used an orthogonal experiment to expose delayed gametophytes ofto different light intensities, light qualities, and photoperiods for 12 days. The results showed that changes in light intensity rather than light qua- lity and photoperiod significantly affected the relative growth rates of the delayed gametophytes. Blue light had the greatest promo- tion on reproduction rate.The optimal light conditions in the early vegetative growth phase in gametogenesis induction for the delay- ed gametophytes were at 60–80μmol photons m−2s−1with daylength of 12 or 16 hours under white or blue light. When the delayed gametophytes were maintained in a constant light condition from delayed state to gametogenesis, the beneficial photoperiods for vegetative growth and reproductive rate were both 16L (16 hours of light): 8D (8 hours of dark). However, when the delayedgametophytes achieve the optimal growth state during the first 6 days and then they were cultured at different light conditions for the following 6 days, the reproduction rate increased as the daylength decreased and attained a peak value in group of 8L:16Dphotoperiod, indicating that photoperiod adjustment at the transition period is crucial in the gametogenesis induction process of de- layed gametophyte of
; light intensity; light quality; photoperiod;gametogenesis; vegetative growth
1 Introduction
, which has been cultured with a large-scale in Northeast Asian countries, is one of the im- portant economic seaweeds (Tseng, 2001; Buschmann., 2017; Liu., 2017). It exhibits an obligate heteromor- phic life history that alternates between two generations ofmacroscopic sporophyte and microscopic gametophyte (Lü- ning, 1990; Schiel and Foster, 2006). In general, the micro- scopic gametophyte generation can be divided into three stages from the liberation of the zoospores to the forma-tion of zygotes in 10–13 days, including gametophyte for- mation stage (about 2–3d), gametophyte vegetative growth stage (about 6d), and gametophyte reproduction stage (2–3d) (Lüning, 1980; Tseng, 1987).Some studies have de- monstrated that gametophyte ofcan survive du-ring extended periods of poor environmental quality (., unsuitable irradiance and temperature, and poor nutrients), delaying growth and/or reproduction (Fang., 1978; Lü- ning, 1980; Yang., 2007). Thus, the delayed gameto- phytes can be considered as ‘seed bank’ and has evolved as an effective way for long-term preservation of thegermplasm (Carney., 2006; Yang., 2007).Moreover, the delayed gametophytes are the parental re- sources of thevariety and hybrid breeding, which have been often used in the commercial production of sporeling-raising of(Cui., 2017). Thereis a trade-off between the vegetative growth and reproduc- tion of kelp gametophyte, and the induction of gametoge- nesis showed a contrasting pattern to gametophyte vege- tative growth (Izquierdo., 2002; Bartsch., 2008; Mohring., 2013; Karasov., 2017; Liu., 2017). For example, the optimum temperature range for gameto- phyte vegetative growth is wider than that for reproduction, and the minimal demand of light intensity and nutrient le- vel for vegetative growth is lower than that for reproduc- tion (Lüning and Neushul, 1978; Lüning, 1980; Martins., 2017; Wang., 2020). Many studies have report- ed that the transition between a vegetative growth phase and the reproduction phase in seaweed is a way to control the life cycle of seaweed (Cock., 2014; Charrier., 2017; Martins., 2017; Ratcliff., 2017).
Whether the kelp gametophyte reproduce sexually or de- lay development can be substantially influenced by abiotic factors such as light (., light intensity, photoperiod, and light quality), temperature, and nutrient availability (Lü- ning and Neushul, 1978; Xu., 2009; Sui., 2011; Martins., 2017; Ratcliff., 2017). Gametogenesis was successfully induced by broad light intensity gradients(Lüning, 1980; Lee and Brinkhuis, 1988). Optimal light in- tensity for gametogenesis was different between kelp spe- cies, which is closely related to their geographical distribu- tion (Lüning and Neushul, 1978; Lüning, 1980). Gameto- genesis could not be induced under extremely low light in- tensity (., 2–4.5μmolphotonsm−2s−1) in most cases inthe kelp species (Bartsch., 2008). Studies have shown that blue light is an inductive condition necessary for the reproduction of some kelps (Lüning and Dring, 1972; Lü- ning and Neushul, 1978; Lüning, 1980). Only a small amount of gametophyte can reproduce at red light (Lü-ning and Dring, 1972). In a general way, long photoperiodis conducive to the vegetative growth of gametophytes, whe- reas optimal photoperiod for gametogenesis was different between kelp species. For example, long photoperiod was conducive to the gametogenesis of(Mar-tins., 2017), while short photoperiod was beneficial to the gametogenesis of(Zhang., 2008)Hence, light intensity, photoperiod, and light quality all areprime candidates as abiotic filters potentially capable of in-fluencing the growth and reproduction of kelp gametophy- tes.
Delayed development of gametophytes is likely impor- tant for the recruitment of some kelp sporophyte (Carney and Edwards, 2006, 2010; Carney, 2011). It was reported that the meiospores releasing from sori of kelp sporophyte show a higher fecundity than pre-cultivated red-light grown gametophytes with delayed development (Izquierdo., 2002). Carney and Edwards (2010) found that delayed ga- metophytes can produce sporophytes 30% faster than ga- metophytes that had never been delayed once development is resumed. These results indicated that the optimal condi- tions for the reproduction of delayed gametophytes might be different from general gametophytes without delaying. How the terrestrial plants that form long-lived seed or seed- ling banks during unfavorable periods resume development when resources are renewed has been studied extensively (Grime, 2001; Makana and Thomas, 2005). However, the effects of environmental factors on the vegetative growth phase, reproduction phase, and the transition between these two phases of delayed gametophytes ofremain to be further researched.
Previous studies have commonly examined the effects of a single factor of light (., light intensity), or dual fac- tors of light (., light intensity and light quality, or light intensity and photoperiod) on the vegetative growth and reproduction of kelp gametophytes (Choi., 2005; Ebb- ing., 2020). However, to the best of our knowledge, the combined effect of all light intensity, light quality, and photoperiod has never been investigated in an orthogonal design for the reproduction of delayed gametophytes. In addition, considering that the induction of gametogenesis showed a contrasting pattern to vegetative growth of game- tophyte, culture conditions of light during the transition pe- riod may need to be further determined. Here, we design- ed an experiment to evaluate how the interaction of three environmental factors related to light influences the vege- tative growth and reproduction of delayed gametophytes of, aiming to seek the optimal light variables for commercial production of sporeling-raising of.
2 Materials and Methods
2.1 Gametophyte Culture
Mature sporophyte of‘Yudai No. 1’ strain, which had a prolonged harvest period with higher yield, was collected from afarm in Dalian city, Chi- na, in July 2018. Sporophyte blade in wet condition was put in sterile and sealable plastic bagsand trans- ported to the laboratory in a cooler (<10℃) in dark within 4h. The sample was rinsed three times in plastic tray with filtered (0.22μm pore size) and autoclaved seawater to re- move any attached fouling organisms. Zoospore release wasinduced by immersing sori tissue in filtered seawater at 10℃. The zoospores attached to the glass microscope slidesand germinated there. Female and male gametophytes werepicked up separately when their gender was distinguishable. The selected gametophytes were transferred into NaNO3(4mgL−1NO3−-N) and KH2PO4(0.4mgL−1PO43−-P) en- riched seawater, and cultured to a desirable amount of bio- mass. Then, the vegetative gametophytes were maintained in f/2 nutrient solution in artificial seawater without ironat low red light intensity (2–4μmolphotonsm−2s−1) under a 12h:12h light/dark (L:D) cycle at 4℃ for at least 1 year to obtain reproduction-delayedgametophytes.
2.2 Experiment 1
Previous researches have reported that gametogenesis in- duction for most kelp gametophytes performed well in the light intensity range of 20–80μmolphotonsm−2s−1within daylength of 8–16h under white or blue light (Akiyama,1965; Hsiao and Druehl, 1971; Choi., 2005; Sui., 2011). Therefore, based on the previous studies, an ortho- gonal experiment (experiment 1) was made to determine the tentative ranges of light intensity, light quality, and photo- period in gametogenesis induction for the delayedgametophytes, and the experimental levels were 3 light intensities (40, 50, and 60μmolphotonsm−2s−1), 3 light qua-lity (white, red, and blue light), and 3 photoperiods (L:D= 8:16, 12:12, 16:8). An orthogonal array with totally 9 treat- ments was conducted as shown in Table 1. Three biologi- cal replicates were carried out for each treatment.

Table 1 Orthogonal design in gametogenesis induction for the delayed S. japonica gametophytes
The female and male delayed gametophytes were mix- ed (fresh weight ratio of female:male=2:1) and smashed by blender, then filtered with 200-mesh sieve. A total of 27 petri dishes (9cm in diameter) were used for the above 9 treatments, each containing a glass microscope slide and en- riched seawater (supplemented with 4mgL−1NO3−-N and 0.4mgL−1PO43−-P). The petri dishes were inoculated with the fragmented gametophytes (40–100μm) to make a set- tlement density of 15–20 fragmented gametophytes in a field of view of 100× magnification and cultured at 10℃for 12 days. The seawater was renewed with 50% fresh ma-terial every three days. The gametophytes were observedusing the microscope (Nikon E200) at the beginning of cul- ture and then on days 2, 4, 6, 8, 10, and 12 during the cul- ture, and ten fields of view (100× magnification) were se- lected randomly and photographed for each microscope slide in each observation.
From the above microscopic observations, we found thatthe oogonium did not appear until the 8th day of culture.Therefore, we divided the process of gametogenesis induc- tion into two phases: early vegetative growth phase (repre-sented by Phase I, when delayed gametophytes enhance theirnumber of cells and increase their cell volume, which is fromthe beginning to the 6th day of culture) and reproductivephase (represented by Phase II, when gametophytes form gametangium and gametes, finally resulting in sporophyte recruitment).
The photographs were analyzed using ImageJ software (developed by Wayne Rasband, National Institutes of Health,Bethesda, MD, USA) to respectively measure the femaleand male vegetative gametophytes’ sizes on the 6th day of culture. Then, the number of vegetative gametophyte (N1), oogonium (N2), eggs or zygotes (N3), and sporophytes (N4) were counted on the 12th day of culture.
The relative growth rates () of the female and male vegetative gametophytes were calculated using the cell area, respectively. The formula was as follows:
,
where2and1were the gametophyte cell area at2and1respectively.
Reproduction percentage ofgametophyte was calculated by the formula:

2.3 Experiment 2
The results of the experiment 1 showed that the growth rate of delayedgametophyte was the lowest un-der photoperiod of 8L:16D and 40μmolphotonsm−2s−1, andincreased with the increase of light time and light intensity in the Phase I, suggesting that longer daylength and high- er light intensity may be more conducive to the growth rate in the Phase I. Therefore, in order to obtain the optimum light intensity and photoperiod in the Phase I for the delay-edgametophytes, more treatments with various light intensities (60, 80, and 100μmolphotonsm−2s−1) and different photoperiods (12L:12D, 16L:8D, and 24L:0D) un- der white light were further conducted. Three replicates were carried out for each treatment. The other culture conditionsandcalculation method were the same as those in Sec- tion 2.2.
2.4 Experiment 3
In the experiment 1, thegametophytes were maintained under a constant light condition from delayed state to gametogenesis during the 12-day gametogenesis in- duction process in each treatment. However, the previous studies showed that the light need between the early vege- tative growth phase and the subsequent reproductive phasewas different during gametogenesis induction process (Bar- tsch., 2008; Mohring., 2013; Liu., 2017). Thus, culture conditions of light in the transition period be- tween Phase I and Phase II were changed in this experimentdesign. Based on the results of the experiment 1, the game- tophytes were cultured at relatively appropriate light condi- tions (., light intensity of 60μmolphotonsm−2s−1, photo- period of 12L:12D, and white light) for 6 days to make thedelayedgametophytes to achieve the optimal growth state in Phase I. Then, these delayed gametophytes were cultured at different light conditions for the follow- ing 6 days (Phase II). These experimental conditions were as follows: 1) three light intensities (60, 80, and 100μmolphotonsm−2s−1) with photoperiod of 12L:12D under white light; 2) three light qualities (white, red, and blue light) with photoperiod of 12L:12D at light intensity of 80μmolpho- tonsm−2s−1; 3) four photoperiods (8L:16D, 12L:12D, 16L: 8D, 24L:0D) with light intensity of 80μmolphotonsm−2s−1under white light. Three replicates were carried out for each treatment. The other culture conditions and reproduction rate calculation method were the same as those in Section 2.2.
2.5 Statistical Analysis
Data were analyzed using the SPSS 19.0 statistical soft-ware packages. All values are presented as the means±stan- dard deviation (mean±SD). Statistical variance analyses were performed using one-way ANOVA (Analysis of Va- riance) and compared with Duncan Multiple Comparisons Test. The statistical significance was set at<0.05.
3 Results
3.1 Microscopic Observation of Delayed Gametophytes During the Gametogenesis Induction
The morphological changes of delayedgame- tophytes during the gametogenesis induction processwereshowed in Fig.1. During the stage of Phase I (from the be- ginning to the 6th day of culture), the cell number (from 2–3 to 7–9) and cell volume of the fragmented gametophy-tes increased as showed in Figs.1c and 1d. The color of cy- toplasm deepened and cytoplasmic inclusions became more homogeneous. The width of the female gametophyte cells increased from 9.3μm to 15.2μm. In the stage of Phase II, the color of cytoplasm became deeper and the female ga- metophyte developed a single large oogonium which pro- duced one egg (Figs.1e and 1f). The eggs were fertilized, producing the fertilized zygotes as shown in Fig.1g. The zygotes germinated and developed into young sporophy- tes (Fig.1h).

Fig.1 Microphoto of delayed S. japonica gametophyte during the gametogenesis induction process at 60μmolphotonsm−2s−1under a 12h:12h white light/dark cycle at 10℃. Scale bar=25μm. (a), fragment of delayed female gametophyte; (b), frag-ment of delayed male gametophyte; (c), vegetative female gametophyte; (d), vegetative male gametophyte; (e), developing oogonium; (f), discharged egg attached to oogonium; (g), mature oogonium and zygote; (h), sporelings.
3.2 RGR of Delayed Gametophyte in Phase I in Experiment 1
ANOVA showed that bothof female and male ga- tophytes in Phase I were affected significantly by light in- nsity (<0.05) rather than light quality and photoperiod. Moreover, according to the max-min values in Table 2 and Table 3, it was found that the light intensity had a greater impact on, followed by light quality. As Table 2 and Table 3 show, bothof female and male gametophytesincreased with the increase of light intensity in Phase I, reaching a peak value at 60μmolphotonsm−2s−1under whiteorblue light. When gametophytes were cultured at 40μmolphotonsm−2s−1and with a photoperiod of 8L:16D, bothof female and male gametophytes were significantly de- eased (<0.05).
3.3 RGR of Delayed Gametophytes in Phase I in Experiment 2
As Fig.2 shows, bothof female and male game- phytes reached a peak value at 80μmolphotonsm−2s−1un- der photoperiod of 16L:8D on the 6th day of culture. Un- der photoperiod of 12L:12D, bothof female and male gametophytes decreased significantly (<0.05) at 100μmolphotonsm−2s−1when compared with groups of 60 and 80μmolphotonsm−2s−1. There was no significant difference inof female gametophyte among the treatment groups of 24L:0Dphotoperiod (>0.05). Significant decreases inof 24L:0Dphotoperiod groupswere observed as com-pared with 16L:8Dphotoperiod groups (<0.05) when cul- tured at the same light intensity.
3.4 Reproduction Rate of Delayed Gametophyte in Phase II in Experiment 1
ANOVA results showed that light intensity, light quality, and photoperiod all had a significant impact on the repro- duction rate of delayed female gametophyte (<0.05). Ac-cording to the max-min values in Table 4, it was found that the light quality had a greater impact on reproduction rate, followed by photoperiod. As Table 4 shows, the reproduc- tion rate of female gametophyte increased with the increase of daylength. The reproduction rate of gametophyte reach- ed a peak value when cultured at blue light, while it was zero under red light. Meanwhile, the reproduction rate of game- tophyte was higher at 60μmolphotonsm−2s−1than those at 40 and 50μmolphotonsm−2s−1.
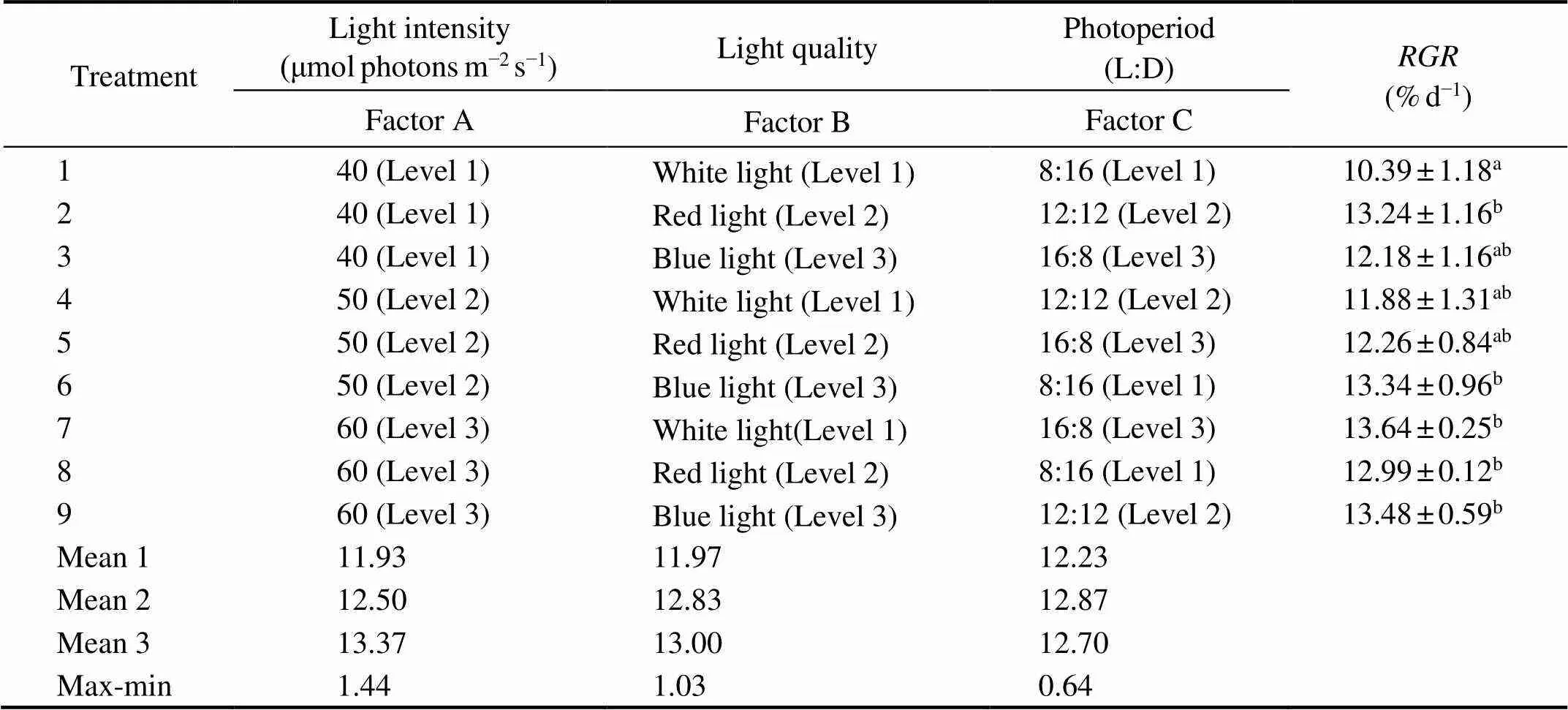
Table 2 RGR of female S. japonicagametophyte on the 6th day of culture in the orthogonal experiment
Notes: The different letters in the superscript ofvalues represent significant differences (<0.05). Data ofare the mean±SD (=30). Mean(=1, 2, 3) is defined as mean value of the indexes of levelfor factor(=A, B, C). Max-min is defined as the range between the maximum mean value and minimum mean value in the column of the corresponding factor.

Table 3 RGR of male S. japonica gametophyte on the 6th day of culture in the orthogonal experiment
Notes: Same as those in Table 2.
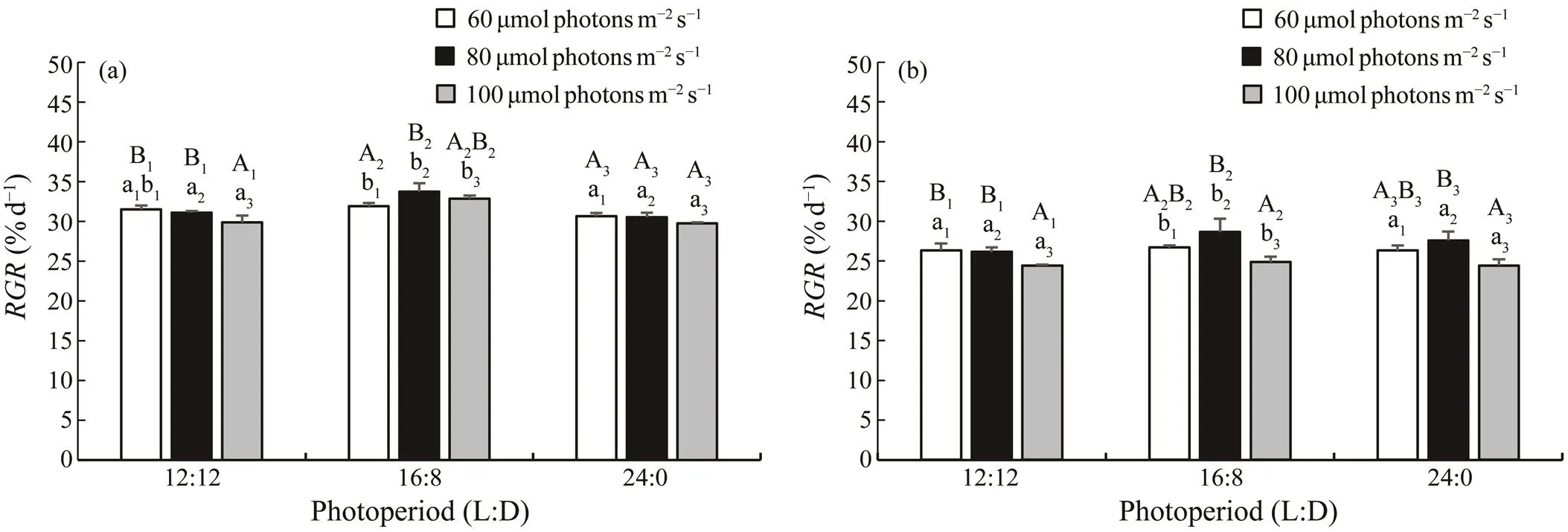
Fig.2 RGR of female (a) and male (b) S. japonicagametophytes at different light intensities and photoperiods under whitelight on the 6th day of culture. The groups with the same subscript of uppercase letters were compared, and the groups withthe same subscript of lowercase letters were also compared. The different letters represent significant differences (P<0.05). Data are the mean±SD (n=30).
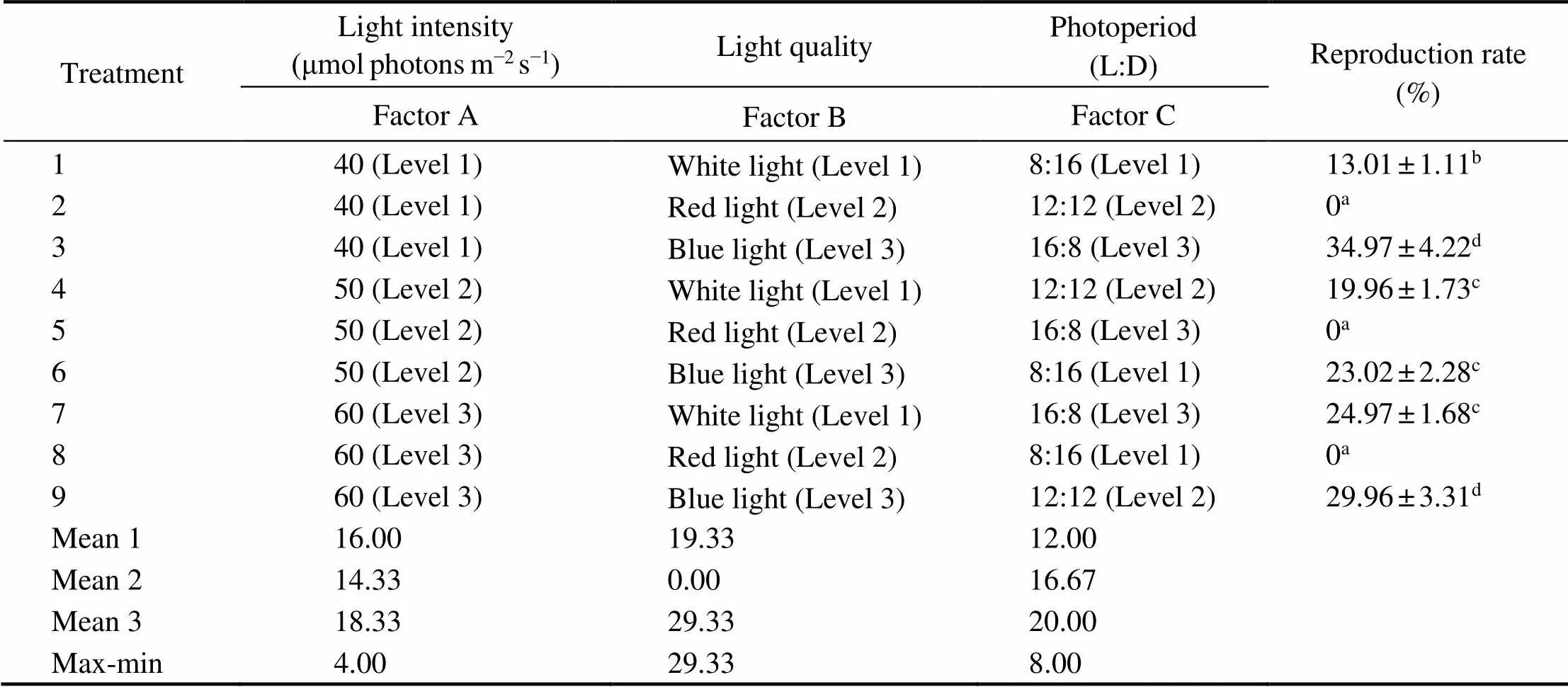
Table 4 Reproduction rate of delayed female gametophyte of S. japonica on the 12th day of culture in the orthogonal experiment
Notes: Same as those in Table 2.
3.5 Reproduction Rate of Gametophyte in Experiment 3
As Fig.3a shows, the reproduction rate of the group withlight intensity of 100μmolphotonsm−2s−1was significant- ly lower than those of the groups with light intensities of 60 and 80μmolphotonsm−2s−1(<0.05). In addition, there was no significant difference in reproduction rate between the treatment groups with light intensities of 60 and 80μmol photonsm−2s−1(>0.05). As Fig.3b shows,the gameto- phyte cultured at blue light had significantly higher repro- duction rate than that cultured at white light (<0.05). Moreover, the reproduction rate of gametophyte cultured at red light decreased significantly compared to that cul- tured at blue light or white light (<0.05). As Fig.3c shows,the reproduction rate increased as the daylength decreas- ed and attained a peak value in group of 8L:16Dphotope-riod.

Fig.3 Reproduction rate ofgametophyte at different light intensity (a), light quality (b), and photoperiod (c) on the 12th day of culture. Different letters above the bars indicate significant differences (<0.05). Data are the mean±SD (=30).
4 Discussion
The technology of sporeling-raising using gametophyte clones has been developed in some kelp species such as,andre- cently (Li., 2002; Westermeier., 2006; Zhang., 2008; Xu., 2009), which needs two basic prerequisitesfor application in commercial production: 1) enough bio-mass of gametophyte accumulated by rapid vegetative growth; 2) efficient induction of gametogenesis by culture condition regulation (Zhang., 2008). The delayed ga- metophytes can remain viable for up to many years dur- ing conditions that are not conducive for sexual reproduc- tion, and keep highly sensitive to changes in environment
quality (Edwards, 2000; Carney and Edwards, 2006; Zhao., 2016). How environmental factors regulate the delay and resumption of microscopic development was one of the scientific mysteries that are necessary to investigate for sea- weed (Hoffmann and Santelices, 1991). Moreover, the me-chanism that regulates the trade-off between vegetative growth and reproduction in response to the changes of en- vironmental conditions is critical to understand how seaweedintegrate environmental signals and adjust its life cycle ac- cordingly (Suda and Mikami, 2020).
Light exerts major impacts on the growth, photosynthe- tic activity, biochemical processes, and reproduction in sea- weed, and seaweed have evolved to deal with the environ- mental fluctuations by regulating metabolism, changes in pigment concentration, enzyme activity, and so on (Kim., 2011; Bischof and Rautenberger, 2012). As a generic abio- tic factor controlling gametogenesis, light has been the fo- cus in many studies in regulating the delay and resumptionof microscopicdevelopment. For example, the delayedgametophytes grown under low light levels and short-day photoperiods could resume growth when trans- ferred to higher light and longer photoperiods (Edwards,2000). The developments of most macroalgae at microsco- pic stages were able to be delayed for at least 60 days and resume growth due to increasing light intensity and day- length (Santelices., 2002). The interaction of tempe- rature and light intensity can make the delayed gametophy- tes ofwith more than one year of vegetative growth reproduce sexually reliably (Ebbing., 2021).
According to Table 2 and Table 3, we found thatlight in- tensity rather than light quality and photoperiod significant- ly affected theof the delayed gametophytes, indicat- ing that light intensity is the primary factor that determineswhether the delayedgametophytes can grow ra- pidly in the early vegetative growth phase of the process of gametogenesis induction. According to Fig.2,of ga- metophytes reached a peak value when the light intensity was at about 80μmolphotonsm−2s−1, which was consistent with the findings of previous studies in other kelp species (Yang., 2002; Ebbing., 2020). Combining the re-sults of Table 2, Table 3 and Fig.2, we proposed that the op- timal light conditions in the early vegetative growth phase of gametogenesis induction for the delayedga- metophytes were 60–80μmolphotonsm−2s−1, 12L:12D or 16L:8D, and white light or blue light. Once sufficient bio- mass has accumulated, the initiation of reproductive struc- tures development of gametophytes can be triggered (Rat- cliff., 2017). It has been proved that blue light is a ne-cessary factor for sexual reproduction ingame- tophytes, and the gametophytes will delay in a vegetative state without blue light (Lüning and Dring, 1972; Lüning, 1981). The light quality results of this study (Table 4) show- ed that blue light had the greatest promotion effect on re- production rate ofgametophyte as comparedto white light and red light, and red light inhibited gameto- genesis, which is consistent with the previous reports (Mi- zuta., 2007; Sui., 2011).
In the commercial production of sporeling-raising ofbased on gametophyte, the delayed gametophytesmight first undergo large-scale vegetative growth for a pe-riod of time, then the culture condition was changed to makethem enter the transition period, and enter the gametoge-nesis stage afterwards. Therefore, it is crucial to understandthe culture conditions of light during the transition periodbetween the vegetative growth phase and reproduction phase.In the present study, we changed the light conditions in the transition period between Phase I and Phase II in the ex- periment of Section 2.4. It was interesting to find that the gametophytes which had received an inductive signal du- ring white light cultivation were able to become fertile eventransferred to non-inductive red light conditions (Fig.3),which was different from that in the orthogonal experimentwith reproduction rate being zero in group of red light.Phy- tohormones act as vital switchers in the regulation of va- rious aspects of development in plant (Weyers and Pater- son, 2001;Saidi and Hajibarat, 2021). Mizuta. (2007) found thatlight quality influences internal IAA metabolismin the sporophytic stage of. In Laminariales plants, the IAA content is higher in younger tissues or the vegetative parts (Williams, 1949; Kai., 2006). There- fore, we hypothesized that the interrelationship between light quality and internal IAA metabolism may also exist in the gametophytic stage,., blue/white light probably reduced IAA content in the gametophyte thus promoting gameto- genesis while IAA content was probably at a high level un- der red light. Phytohormone is regarded as an enduring mes-sage, which possibly remains active long after the immedi- ate and possibly temporary stimulus has disappeared (Wey- ers and Paterson, 2001). This may be the reason why game- togenesis can still occur in some degree when the gameto- phytes are transferred from white light (cultured for 6 days)to red light (Fig.3). Pearson. (2019) found that a switchin culture irradiance from red to white light activated a core set of genes, involving rapid activation of ribosome bioge-nesis, as well as transcription- and translation-related path-ways. These genes and pathways probably can provide foun- dation for the future understanding of the inhibition of red light on gametophyte reproduction.
Thegametophytes were generally maintain-ed under a constant light condition from delayed state to ga-metogenesis in the whole gametogenesis induction process in some previous studies, which obtained different photo- period conditions for gametogenesis. For example, Hsiaoand Druehl (1971) found that long period of light (16L:8D) was beneficial to the reproduction of, but continuous light can significantly reduce the re- productive rate. Martins. (2017) also found that the reproductive rate ofgametophytes under long pe-riod of light (16L:8D) was significantly higher than that under short term of light (8L:16D). However, some studies have found that shortening light time was beneficial to the transition from vegetative growth to reproductive develop- ment forgametophytes (Zhang., 2008; Xu., 2009). Moreover, short daylength has been applied to commercial production of sporeling-raising ofbased on gametophyte (Xu., 2009).
When thegametophytes were maintained un- der a constant light condition from delayed state to game- togenesis in the study (Tables 2, 3, 4), the beneficial pho- toperiod condition for vegetative growth and reproductive rate was both at photoperiod of 16L:8D. We hypothesized that rapid vegetative growth may promote the gametogene-sis since the delayed gametophyte need to undergo vegeta- tive growth before gametogenesis. However, it was worth emphasizing here that, when the delayedgame- tophytes achieve the optimal growth state in Phase I (first 6 days) and then they were cultured at different light con- ditions for the following 6 days, the reproduction rate in- creased as the daylength decreased and attained a peak va-lue in group of 8L:16Dphotoperiod as showed in Fig.3, which was different from that in the orthogonal experimentwith reproduction rate increasing as the daylength increased (Table 4). Our results suggest that light time shortening at the transition period from vegetative growth phase to re- production phase is crucial in the gametogenesis induction process of delayed gametophyte of
5 Conclusions
The light intensity had the greatest impact onof de- layed gametophytes of, followed by light qua- lity.reached a peak value at 60–80μmolphotonsm−2s−1under white/blue light. The light quality had the great- est impact on reproduction rate of delayed gametophyte of, followed by photoperiod. The gametophyte re- production rate was considerably higher when cultured at the blue/white light than that at the red light. Rapid vegeta- tive growth may promote the gametogenesis. When the de- layedgametophytes achieve the optimal growthstate in vegetative growth phase, short daylength was bene-ficial for them during the transition from vegetative growth phase to reproduction phase.
Acknowledgements
This study was financially supported by the National KeyR&D Program of China (Nos. 2018YFD0900305, 2018YFD0901500), the China Agriculture Research System of MOF and MARA, Central Public-interest Scientific InstitutionBasal Research Fund CAFS (No. 2020TD27), and the‘Young Talent of Fishery Sciences’ project from Laboratory for Marine Fisheries and Aquaculture (No. 2018-MFS-T12).
Akiyama, K., 1965. Studies of ecology and culture of(Harv.) Sur. II. Environmental factors affecting the growing and maturation of gamtophyte., 25: 143-170.
Bartsch, I., Wiencke,C., Bischof,K., Buchholz, C. M., Buck, B. H., and Eggert, A.,2008. The genus: Recent insights and developments.,43(1): 1-86, https://doi.org/10.1080/09670260701711376.
Bischof, K., and Rautenberger, R., 2012. Seaweed responses to environmental stress: Reactive oxygen and antioxidative strate-gies. In:.Wiencke, C., and Bischof, K., eds., Springer, Berlin, 109-132.
Buschmann, A. H., Camus, C., Infante, J., Neori, A., Israel, Á., Hernández-González, M. C.,.,2017. Seaweed production: Overview of the global state of exploitation, farming and emerg- ing research activity., 52:391-406, https://doi.org/10.1080/09670262.2017.1365175.
Carney, L. T., 2011.A multispecies laboratory assessment of ra- pid sporophyte recruitment from delayed kelp gametophytes., 47: 244-251, https://doi.org/10.1111/j.1529-8817.2011.00957.x.
Carney, L. T., and Edwards, M. S., 2006. Cryptic processes in the sea: A review of delayed development in the microscopic lifestages of marine macroalgae., 21(2): 161-168, https://doi.org/10.4490/ ALGAE.2006.21.2.161.
Carney, L. T., and Edwards, M. S.,2010. Role of nutrient fluctu- ations and delayed development in gametophyte reproduction by(Phaeophyceae) in southern California., 46: 987-996, https://doi.org/10.1111/j.1529-8817.2010.00882.x.
Charrier, B., Abreu, M. H., Araujo, R., Bruhn, A., Coates, J. C., De Clerck, O.,., 2017. Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture., 216(4): 967-975, https://doi.org/10.1111/nph. 14728.
Choi, H. G., Kim, Y. S., Lee, S. J., Park, E. J., and Nam, K. W., 2005. Effects of daylength, irradiance and settlement density on the growth and reproduction ofgame- tophytes., 17: 423-430, https:// doi.org/10.1007/s10811-005-0432-2.
Cock, J. M., Godfroy, O., Macaisne, N., Peters, A. F., and Coel- ho, S. M., 2014. Evolution and regulation of complex life cy- cles: A brown algal perspective., 17: 1-6, https://doi.org/10.1016/j.pbi.2013.09.004.
Cui, C., Li, Y., Liu, Y., Li, X., Luo, S., and Zhang, Z., 2017. De- termination of genetic diversity amonggermplasm using ISSR and RAPD markers.,340: 76-86, https://doi.org/10.1016/j.crvi.2016.11.005.
Ebbing, A., Pierik, R., Bouma, T., Kromkamp, J. C., and Timmer-mans, K., 2020.How light and biomass density influence the reproduction of delayedgametophytes (Phaeophyceae)., 56: 709-718, https://doi.org/10.1111/jpy.12976.
Ebbing, A., Pierik, R., Fivash, G., van de Loosdrecht, N. C. J., Bouma, T., Kromkamp, J. C.,., 2021. The role of seasona- lity in reproduction of multiannual delayed gametophytes of., 57: 1580-1589, https://doi.org/10.1111/jpy.13191.
Edwards, M. S., 2000. The role of alternate life-history stages of a marine macroalga: A seed bank analogue?, 81: 2404- 2415, https://doi.org/10.1890/0012-9658(2000)081.
Fang, Z. X., Ou,Y. L., Cui,J. J., and Dai, J. X.,1978. Success in culturing clones of the gametophytes of., 23:115-116 (in Chinese with English abstract).
Grime, J. P., 2001.. Wiley, Chichester, UK, 417pp.
Hoffmann, A. J., and Santelices, B., 1991. Banks of algal micro- scopic forms–Hypotheses on their functioning and compari- sons with seed banks., 79: 185- 194,https://doi.org/10.3354/meps079185.
Hsiao, S. I. C., and Druehl, L. D., 1971. Environmental control of gametogenesis in. I. The effects of light and culture media.,49: 1503-1508, https://doi.org/10.1139/b71-211.
Izquierdo, J. L., Pérez-Ruzafa, I. M., and Gallardo, T., 2002. Ef- fect of temperature and photon fluence rate on gametophytes and young sporophytes ofPylaie., 55: 285-292, https://doi.org/10.1007/ s10152-001-0087-6.
Kai, T., Nimura, K., Yasui, H., and Mizuta, H., 2006. Regulation of sorus formation by auxin in Laminariales sporophyte., 18: 95-101, https://doi.org/10.1007/ s10811-005-9020-8.
Karasov, T. L., Chae, E., Herman, J. J., and Bergelson, J.,2017. Mechanisms to mitigate the trade-off between growth and de- fense.,29: 666-680, https://doi.org/10.1105/tpc.16.00931.
Kim, J. H., Kang, E. J., Park, M. G., Lee, B. G., and Kim, K. Y., 2011. Effects of temperature and irradiance on photosynthesis and growth of a green-tide-forming species () in the Yellow Sea., 23: 421-432, https:// doi.org/10.1007/s10811-010-9590-y.
Lee, J. A., and Brinkhuis, B. H., 1988. Seasonal light and tempe- rature interaction effects on development of(Phaeophyta) gametophytes and juvenile sporophytes., 24: 181-191, https://doi.org/10.1111/j. 1529-8817.1988.tb04232.x.
Li, D. P., Lu, Y. H., and Wu, C. Y., 2002. The past and present status ofgenetic breeding and sporophyte culturetechnology., 37: 1-3 (in Chinese with Eng-lish abstract).
Liu, X., Bogaert, K., Engelen, A. H., Leliaert, F., Roleda, M. Y., and De Clerck, O.,2017.Seaweed reproductive biology: En- vironmental and genetic controls., 60: 89-108, https://doi.org/10.1515/bot-2016-0091.
Lüning, K., 1980. Critical levels of light and temperature regula- ting the gametogenesis of threespecies (Phaeophy-ceae).,16: 1-15, https://doi.org/10.1111/j.1529-8817.1980.tb02992.x.
Lüning, K., and Dring, M. J.,1972. Reproduction induced by blue light in female gametophytes of,104: 252-256, https://doi.org/10.1007/BF00387080.
Lüning, K., 1990.. John Wiley and Sons Inc., New York, 527pp, https://doi.org/10.1002/aqc.3270010208.
Lüning, K., and Neushul, M., 1978. Light and temperature de- mands for growth and reproduction ofgametophy- tes in southern and central California., 45: 297-309, https://doi.org/10.1007/BF00391816.
Makana, J. R., and Thomas, S. C., 2005. Effects of light gaps and litter removal on the seedling performance of six African tim- ber species., 37: 227-237, https://doi.org/10.1111/j. 1744-7429.2005.00030.x.
Martins, N., Tanttu, H., Pearson, G. A., Serrao, E. A., and Bartsch, I., 2017. Interactions of daylength, temperature and nutrients affect thresholds for life stage transitions in the kelp(Phaeophyceae)., 60: 109-121, https://doi.org/10.1515/bot-2016-0094.
Mizuta,H., Kai, T., Tabuchi, K., and Yasui, H., 2007. Effects of light quality on the reproduction and morphology of sporophy- tes of(Phaeophyceae).,38: 1323-1329, https://doi.org/10.1111/j.1365-2109.2007.01809.x.
Mohring,M. B., Kendrick, G. A., Wernberg, T., Rule, M. J., and Vanderklift, M. A.,2013. Environmental influences on kelpperformance across the reproductive period: An ecological trade-off between gametophyte survival and growth?,8: e65310, https://doi.org/10.1371/journal.pone.0065310.
Pearson, G. A., Martins, N., Madeira, P., Serro, E. A., and Bar-tsch, I.,2019. Sex-dependent and -independent transcriptional changes during haploid phase gametogenesis in the sugar kelp., 14: e0219723, https://doi.org/10.1371/journal.pone.0219723.
Ratcliff, J. J., Soler-Vila, A., Hanniffy, D., Johnson, M. P., and Edwards, M. D.,2017. Optimisation of kelp () gametophyte growth and gametogenesis: Effects of photo- period and culture media.,29: 1957-1966, https://doi.org/10.1007/s10811-017-1070-1.
Saidi, A., and Hajibarat, Z., 2021. Phytohormones: Plant switch- ers in developmental and growth stages in potato., 19: 89.
Santelices, B., Aedo, D., and Hoffmann, A., 2002. Banks of mi- croscopic forms and survival to darkness of propagules and mi- croscopic stages of macroalgae., 75: 547-555.
Schiel,D. R., and Foster, M. S.,2006. The population biology of large brown seaweeds: Ecological consequences of multiphaselife histories in dynamic coastal environments.,37: 343-372, https://doi.org/10.1146/annurev.ecolsys.37.091305.110251.
Suda,M., and Mikami, K.,2020. Reproductive responses to wounding and heat stress in gametophytic thalli of the red alga.,7: 394, https://doi.org/10.3389/fmars.2020.00394.
Sui, X. W., Ren, W., Yan, W. H., Yan, W. H., Liu, S. P., and Yang, N.,2011. Effects of light wavelength on growth and reproduc- tion of the gametophytes ofAresch, 35: 33-36(in Chinese with English abstract).
Tseng, C. K., 1987.mariculture in China.. Food and Agriculture Organisa- tion of the United Nations, Rome. http://www.fao.org/3/X5819E/x5819e00.htm.
Tseng, C. K.,2001. Algal biotechnology industries and research activities in China., 13: 375-380, https://doi.org/10.1023/A:1017972812576.
Wang, X., Liu, F. L., Liang,Z. R., Zhang, P. X., Yuan, Y. M., Wang, W. J.,., 2020. Effects of iron on the growth and de-velopment of gametophyte clones in.,9: 1052-1061(in Chinese with English abstract).
Westermeier, R., Patiño, D., Piel, M. I., Maier, I., and Mueller, D.G., 2006. A new approach to kelp mariculture in Chile: Produc-tion of free-floating sporophyte seedlings from gametophyte cul- tures ofand., 37: 164-171.
Weyers, J., and Paterson, N. W., 2001. Plant hormones and the control of physiological processes., 152: 375- 407, https://doi.org/10.1046/j.0028-646X.2001.00281.x.
Williams, L. G., 1949. Growth-regulating substances in., 110: 169, https://doi.org/10.1126/science. 110.2850.169.
Xu, B., Zhang, Q., S., Qu, S. C., Cong, Y. Z., and Tang, X. X., 2009.Introduction of a seedling production and method suing ve- getative gametophytes to the commercial farming ofin China., 21: 171-178, https:// doi.org/10.1007/s10811-008-9347-z.
Yang, G. P., Li, X. J., Cong, Y. Z., Qu, S. C., Li, Z. L., Zhang, Z. Z.,,2007. Trends in the cloning methodology ofgametophytes: Research and application., 37: 569-572 (in Chinese with English abstract).
Yang, Y. X., Luo,S. J., Xie, Z. J., and Zhang, Z. Z., 2002. The ef- fect of light intensity on growth of gametophytic clone of., 3: 114-116(in Chinese with English abstract).
Zhang, Q. S., Qu, S. C., Cong, Y. Z., Luo, S. J., and Tang, X. X., 2008. High throughput culture and gametogenesis induction ofgametophyte clones., 20: 205-211, https://doi.org/10.1007/s10811-007- 9220-5.
Zhao, X. B., Pang, S. J., Liu, F., Shan, T. F., Li, J., Gao, S. Q.,., 2016. Intraspecific crossing ofusing dis- tantly related unialgal gametophytes benefits kelp farming by improving blade quality and productivity at Sanggou Bay, Chi-na., 28: 449-455, https://doi.org/ 10.1007/s10811-015-0597-2.
(September 18, 2021; revised November 16, 2021; accepted May 27, 2022)
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2023
#The two authors contributed equally to this work.
Corresponding author. E-mail: liufuli@ouc.edu.cn
(Edited by Qiu Yantao)
杂志排行

Journal of Ocean University of China的其它文章
- The Subduction Structure Beneath the New Britain Island Arc and the Adjacent Region from Double-Difference Tomography
- Differences of Polygonal Faults with Irregularly Polygonal Geometries: A Case Study from the Changchang Sag of Qiongdongnan Basin, Northern South China Sea
- Characterization of Bacterial Communities in Aerosols over Northern Chinese Marginal Seas and the Northwestern Pacific Ocean in Autumn
- Assessment and Application of Beach Quality Based on Analytic Hierarchy Process in Yangkou Beach, Qingdao
- Role of Resuspended Sediments as Sources of Dissolved Inorganic Phosphorus Along Different Dimensions in the Subei Shoal, South Yellow Sea, China
- Pharmacokinetics of Enrofloxacin and Its Metabolite in Carp (Cyprinus carpio) After a Single Oral Administration in Medicated Feed