烟草中木质素调控研究进展*
2023-03-16张建铎张承明陈建华孔光辉吴玉萍李雪梅刘恩芬杨光宇
王 晋,张建铎,许 永,张承明,李 晶,刘 欣,陈建华,孔光辉,吴玉萍,李雪梅,刘恩芬,杨光宇**
(1.云南中烟工业有限责任公司技术中心,云南 昆明 650106;2.云南省烟草农业科学研究院,云南 昆明 650031;3.云南中烟再造烟叶有限责任公司,云南 昆明 650106)
木质素是植物体内的酚类大分子聚合物,是次生细胞壁的主要成分。在自然界界中,木质素含量仅次于纤维素,可用于化工原料和生物质燃料,同时,对于造纸制浆、畜牧饲草也有着重要影响[1-5]。
木质素在烟草中广泛存在,具有重要的生物功能,如:增强植株的物理机械性能,改善茎秆强度[6];有利于水分和营养物质的输送,改善植株的水分保持能力[7-10];防御病原体、线虫等生物侵害,以及干旱、低温等非生物胁迫;等等[11-14]。卷烟燃烧过程中,木质素容易产生木质气、杂气等,引起喉刺涩口等,而木质素燃烧热裂解产生的苯酚、多酚等小分子对健康也存在一定影响。因此木质素对卷烟烟气的感官品质和卷烟的安全性存在重要影响[50-52]。
1 烟草木质素结构及表征
木质素,作为一种复杂的酚类聚合物,按组成单元可分为:愈创木基木质素(Guajacyl Lignin,G-木质素)、紫丁香基木质素(Syringyl lignin,S-木质素)和对苯羟基木质素(Hydroxy-phenyl lignin,S-木质素),他们分别由松柏醇(Coniferyl alcohol)、芥子醇(Sinapyl alcohol)和香豆醇(Coumary alcohol)聚合而成,如图1所示[2,15-16]。烟草中木质素主要是G-木质素和S-木质素。G-木质素结构单元上只含有一个甲氧基,其游离的C-5位置可与其他单体反应形成稳定的C—C交联键,相对来说难分离难降解;S-木质素结构单元上含有两个甲氧基,无游离的C-5,不能形成C—C键,结构较疏松,相对易于分离降解[16]。S/G的比值是评价木质素的一个重要指标,在造纸制浆、再造烟叶和生物质燃料方面有着重要作用[1,6,17]。

图1 木质素单体结构
木质素的表征包括木质素含量的表征和木质素组成的表征。对于木质素总量表征的方法有:间苯三酚法、Kalson法和乙酰溴法等[50-53]。间苯三酚法主要用于活体植株木质素的半定量分析,将植株切片后,利用苯三酚对细胞壁将木质素染成红色,在显微镜下观察颜色分布、深浅来对木质素含量进行分析。Kalson法,利用浓硫酸对样品进行降解,得到的难溶物固体即为木质素,通过称重就可以得到木质素的含量。乙酰溴法,对样品用乙酰溴处理,然后测定 280 nm 下UV的吸收强度。
木质素组成的表征:①Mäule染色,样品利用高锰酸钾和盐酸处理,S-木质素上的紫丁香环变成二甲氧基邻苯二酚后,再以氨水生成甲氧基邻醌,显紫色;而G-木质素不发生显色。因此可利用颜色的深浅进行木质素组成的表征[18]。②中红外法,利用G-木质素和S-木质素的不同红外吸收特征来分析S/G,如 1320 cm-1为愈创木基环结构的红外特征吸收[19]。③拉曼光谱法[54],1270 cm-1归属于愈创木基环醚键伸缩振动,1331 cm-1为紫丁香基环酚羟基弯曲振动,利用其拉曼光谱吸收强度表征S/G。此外,还有核磁法、热解/裂解-气质联用法等。
2 木质素生物合成及代谢路径
木质素生物合成可通过单体合成和单体聚合实现。木质素单体的合成需要依次经过草莽酸途径(Shikimic Acid Pathway)、苯丙烷途径(Phenylpropanoid Pathway)和特异性途径。草莽酸途径得到的苯丙氨酸经过脱氨反应、羟基化、甲基化反应和氧化还原反应等一系列反应,最终生成3种木质素单体(如图2)[1-2,6,14]。单体经过催化聚合得到相应的木质素聚合物,目前,该聚合过程机理尚不清楚。
3 木质素合成酶及其基因调控
3.1 苯丙氨酸氨解酶(phenylalanine ammonialyase PAL)
苯丙氨酸在PAL催化下脱氨形成反式肉桂酸,该反应为木质素单体合成在苯丙烷途径中的第一步反应。在转基因烟草中,PAL含量或活性增强,木质素含量增加;反之,木质素含量降低,但是要使木质素水平显著降低,则需要将PAL活性降低至野生型的10%以下,而过度抑制PAL的活性将导致植株矮化,抗逆性、环境适应性降低[55]。同时,PAL抑制后,叶片形状发生改变,生长迟缓,花的形态颜色发生变化,花粉活性降低[20]。PAL活性降低,S/G比值增加,甲基含量增加[21,56]。
PAL不仅是烟草木质素合成关键酶,同时也对类黄酮代谢物等合成存在影响。PAL是合成绿原酸的关键控制点,但是对于芸香苷影响不大,PAL活性和绿原酸含量之间近似线性正相关[56]。PAL活性降低,可溶性苯丙烷类产物降低[20]。此外,生长发育和环境等将调控影响PAL2启动子的时空表达,调节多种苯丙烷类天然产物的合成[22]。
3.2 肉桂酸-4-羟基化酶(cinnamate-4-hydroxylase,C4H)
C4H是木质素单体合成的第二个限速酶,属于植物细胞色素P450(CYP450)单加氧酶中的一种。C4H催化底物反式肉桂酸羟基化为对-香豆素。与PAL一样,C4H不仅参与木质素的合成,也参与类黄酮等苯丙烷代谢物的合成。在转基因烟草中,C4H活性降低,木质素含量降低,并伴随着S/G比率降低[23,24]。
3.3 4-香豆酸辅酶A连接酶(4-Coumarate:CoA ligase,4CL)
4CL是苯丙烷代谢途径的第三个限速酶,是木质素的正向调控酶,调节木质素的生物合成[24-27]。
4CL可将多种底物如阿魏酸、咖啡酸和对香豆素等羟基肉桂酸类代谢物,催化成对应的羟基肉桂酸CoA酯。在4CL有义和反义调控转基因烟草中,4CL下调,木质素细胞壁中香豆酸、阿魏酸和芥子酸等3种羟基肉桂酸含量增加[25-27],木质部细胞壁由白色变为棕色;4CL活性增加,花中山柰酚含量增加了2倍[24]。
4CL还能调控木质素的组成。4CL下调,G木质素和S木质素都降低,但是G木质素降低更多,S/G值增加。
3.4 阿魏酸5-羟基化酶(Ferulate 5-hydroxylase,F5H)
F5H是木质素合成中除C4H外的另一个重要的羟基化酶,也属于细胞色素 P450 家族[18]。在木质素合成过程中,F5H可将阿魏酸、松柏醛、松柏醇等催化得到相应的5位羟基化化合物如5-羟基阿魏酸、5-羟基松柏醛和5-羟基松柏醇等,是S木质素合成的关键酶[57]。在转基因烟草中,F5H的基因沉默和过表达对植株木质含量和生长影响较小[57],但是S木质素会相应的降低和增加[18],因此,F5H基因是调控木质素组成的很好的调控位点。
3.5 O-甲基转移酶(O-methyltransferases,OMTs)
植物的O-甲基转移酶(OMTs)家族分为两种亚型[16],其中,咖啡酸-O-甲基转移酶(caffeic acid O-methyltransferase,COMT)属于I型OMTs,咖啡酰-辅酶 A 甲基转移酶(caffeoyl CoA 3-O-methyltransferase,CCoAOMT)属于II型[27,58-60]。Maury认为,烟草中存在两种COMT类型(I和II)和三种不同的CCoAOMT类型[14]。CCoAOMTs在茎中表达量最高,它们参与G木质素前体的合成。Pellegrini和Martz等[12-13]研究表明,COMT I基因专门参与木质素的生物合成,而COMT II受病源物感染强烈诱导。
COMT可催化酸、醛或醇如咖啡酸、5-羟基阿魏酸、5-羟基松柏醛和5-羟基松柏醇等代谢物苯环上C5位的酚羟基甲氧基化。CCoAOMT催化底物为咖啡酰CoA、5-羟基阿魏酸CoA等,将其苯环上C5位的酚羟基甲氧基化。甲氧基化可以调控木质素的化学组成及性能。抑制OMT活性能抑制丁香基单元的生物合成,提高S/G值[25-26]。
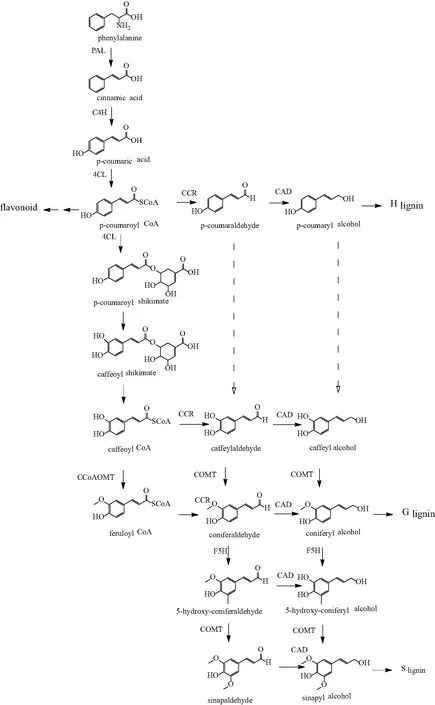
图2 木质素生物合成的苯丙烷代谢路径
3.6 肉桂酰辅酶A还原酶(cinnamoyl-CoA reductas CCR)和肉桂醇脱氢酶(cinamyl alcohol dehydrogenase,CAD)
木质素特异合成途径酶主要有两种:CCR和CAD。CCR为该合成途径的第一限速酶,可将阿魏酰辅酶A、对-香豆酰辅酶A和5-羟基-阿魏酰辅酶A等三种底物分别催化为松柏醛、对-香豆醛和5-羟基-松柏醛。CAD是木质素单体生物合成的最后一步的关键酶,可将对羟基肉桂醛、咖啡醛、松柏醛、5-羟基松柏醛和芥子醛等催化还原相应的醇[28-31]。CCR和CAD的调控都将影响烟草中木质素的含量和组成。Yahiaoui[19,37]等研究了CAD活性显著下调的(对照的8%~56%)的转基因烟草。其S/G显著降低,同时转基因烟草中木质部中检测到松柏醛(coniferyl aldehyde)。他们发现,未转化烟草茎中木质部细胞壁肉桂醛的原位聚合产生红棕色与反义的CAD下调植物木质部的红棕色类似。他们认为肉桂醛的原位聚合是CAD下调植株中木质部变红得的原因。
3.7 聚合
化学上,木质素单体通过脱氢聚合得到木质素,而该生物合成过程仍然处于探索的初级阶段。相关研究表明,负责催化该过程的酶主要是漆酶(Laccase,LAC)和过氧化物酶(Peroxidases,POX),而其调控机理的还有待进一步研究[1-4]。
Kavousi等[25]分别利用四季豆(French bean)和烟草的过氧化物酶FBP1和TP60反义转化烟草。研究表明,除CCR外,所有木质素合成酶转录水平皆下调,木质素降低20%,纤维素和半纤维素合成基因转录水平增加,但是维管多糖含量和组成变化不大。形态上,转基因烟草的木质部欠发达,其纤维和维管的细胞壁薄,且次生壁增厚有限,并带有异常的S2层,但是总体生长正常。转基因烟草,叶片厚度增加和叶尖分叉,叶绿体体积大和气孔少。叶片颜色变浅,叶绿素a,b和类胡萝卜素含量明显降低。
Blee等[26]通过烟草过氧化物酶TP60在烟草中使用反义转化下调,这使转化株的木质素降低高达野生型(对照)植物的40%~50%。通过硫酸解和硝基苯氧化分析,愈创木酰基和紫丁香基的含量显著地随着木质素含量的降低呈线性下降。叶片有波浪状而不是光滑的边缘,花冠减少。
4 木质素合成的转录组调控
转录因子即反式作用因子,是指能够与基因启动子区中顺式作用元件发生特异性结合的结合蛋白,通过蛋白的相互作用调控基因的转录。转录因子对烟草的木质素的合成起重要的调控作用。
NAC(NAM / ATAF / CUC)是最大的植物特异性转录因子(TF)家族之一,在木质素生物合成中起重要作用。Yao在烟草中过表达NAC15基因,结果表明,CAD、PAL、4CL、C4H等木质素相关基因表达水平增加,同时木质素,半纤维素和纤维素含量增加[27]。
MYB(v-myb avian myeloblastosis viral oncogene homolog)也是调控木质素合成代谢的重要的转录因子家族。R2R3-MYB转录因子的4亚群被认为是调节苯丙烷途径的抑制剂。Ma等[28]报道了一个编码小麦4亚群R2R3-MYB因子的cDNA,TaMYB4。TaMYB4基因在茎和根组织中高度表达。酵母细胞的体外结合分析表明,TaMYB4可以与大米的(Rice Cinnamyl Alcohol Dehydrogenase 2) OsCAD2启动子中的AC-II元件相互作用。TaMYB4在转基因烟草中过表达导致了参与木质素生物合成的CAD和CCR基因的转录降低。酶促试验表明,转基因烟草的CAD和CCR活性降低,明显降低了总木质素水平,但S/G比值增加。此外,转基因烟叶中总黄酮含量增加,提示TaMYB4的过表达可能导致了代谢通量从木质素途径转向黄酮类途径。
MYB转录因子在苯丙烷代谢的调节中发挥广泛的作用,同时也调控植物类黄酮代谢。Tamagnone等[29]研究发现,烟草中过表达了金鱼草的AmMYB308和AmMYB330基因后抑制了酚酸代谢和木质素合成,导致植物的维管组织中木质素至少减少17%。研究表明,AmMYB308作为一个非常弱的转录激活子,它的过表达将可能会竞争性地抑制识别相同目标序列的强激活子的活性。
AC 元件也称为 PAL-box 和 H-box,最早从香芹的PAL1 的启动子中分离鉴定来。大部分木质素合成基因如PAL、4CL、CCR 和 CAD的启动子中都含有AC元件。AC元件分为三类AC-1(ACCTACC)、 AC-II(ACCAACC)和AC-III(ACCTAAC),AC元件的活性将调控木质素合成基因的表达。其中,AC-1和 AC-II对木质素相关基因表达影响较大,它们单个的突变将导致木质素相关基因表达降低,它们同时突变则会导致相关木质素基因不表达。而AC-III元件的突变将轻微降低木质素相关基因表达。AC 元件是 MYB 基因调控木质素合成的重要方式。
Lin-Isl-Mec domain (LIM结构域蛋白质) 是由最早克分离得到的Lin-1、Isl-1和Mec-3 三个转录因子的首字母命名。Pal-box富含AC的元件,是苯丙烷途径生物合成中基因表达的重要顺式作用元件。Kawaoka等[30]得到了一个编码Pal-box结合蛋白的基因Ntlim1。Ntlim1的氨基酸序列与LIM蛋白家族中含有锌指基序的成员高度相似。Ntlim1具有特异性的DNA结合能力,由Pal-box序列驱动的β-葡萄糖醛酸酶报告基因(β-glucuronidase reporter,GFP)的瞬时激活转录。烟草细胞瞬时表达试验表明,GFP与Ntlim1的融合蛋白可以进入细胞核。反义Ntlim1转基因烟草显示PAL、4CL和CAD等关键苯丙类途径基因转录量低。反义Ntlim1转基因烟草的木质素含量降低了20%以上。
EgMYB2是转录因子R2R3 MYB家族的一个亚群,Goicoechea等[31]从桉树木质部中克隆得到了EgMYB2。研究表明,EgMYB2蛋白能够特异地结合两个木质素生物合成基因CCR和CAD启动子的顺式调节区域,调节它们的转录。EgMYB2过表达的转基因烟草植株表现出与野生型植株相比次生细胞壁厚度显著增加,木质素含量增加不大(5%),但S木质素增加,G木质素减少,S/G值明显增加。EgMYB2 是次生细胞壁形成和木质素生物合成的正向调节剂。
WRKY转录因子对于植物生长发育、植物抵抗生物胁迫和非生物胁迫具有重要作用,同时也对植物木质素代谢合成起一定的作用。Mzid等[11]研究表明,过表达葡萄(Vitis vinifera L.)转录因子VvWRKY2的转基因烟草植物对坏死性真菌病原体抗病性增强。VvWRKY2参与木质素生物合成途径的VvC4H基因启动子的激活。茎和叶柄中丁香基/愈创木基比值均降低。
AP2/ERF (APETALA 2/ethylene-responsive element binding factor)转录因子家族不仅能够调控植物生长发育,也能调控木质素的生物合成。Liu在烟草中过表达了胡杨PsnSHN2基因[32],它是拟南芥AP2/ERF型转录因子SHINE2的对应基因。烟草中PsnSHN2的过表达显著改变了与次生壁形成有关的转录因子和生物合成基因的表达,导致次生壁比野生型增加了55%,纤维素增加了 37%,半纤维素增加了 28%,木质素减少了 34%,根部断裂应力增加了22%。说明PsnSHN2 协同调节纤维素、半纤维素和木质素的生物合成。转基因烟草的qRT-PCR 分析结果表明,烟草木质素生物合成途径基因PAL1、C4H、4CL1、CCR、COMT、CCoAOMT、CAD等转录水平受到显著抑制。特异性激活木质素生物合成途径基因的 NAC 转录因子(SND1 和 VND6)和 MYB 转录因子(MYB58、MYB63 和 MYB85)的转录水平在PsnSHN2 转基因株系中被显著抑制。相反,专门激活了纤维素和半纤维素生物合成基因的中间开关基因 MYB8311 和几个 MYB 基因(MYB20、MYB43 和 MYB69)的表达水平,在 PsnSHN2 转基因品系中显著增加。PsnSHN2通过选择性上调/下调,控制次级壁形成的下游转录因子(NAC和MYB)来协调调控次级壁形成。
KNOX(Knotted1-like-homeobox)同源盒基因是转录因子,可用于调节发育和农学上重要的植物特征,以优化生物量质量。BREVIPEDICELLUS(BP)/ KNAT1,一种特定的KNOX基因,是拟南芥形态发育以及木质素生物合成的调节剂[67-68]。
BP功能突变将导致花序中木质素生物合成的增加,而该基因的过表达导致植物中木质素沉积的抑制[69]。这些结果表明,BP可通过抑制木质素生物合成酶的作用,从而抑制了木质素的沉积。因此,KNOX基因(如BP)参与控制木质素生物合成。
Townsley等[33]发现过表达Kn1和LeT6(一种来自番茄与拟南芥(Shoot Meristemless)STM直系同源的非BP型KNOX1基因)的转基因烟草叶子呈卷曲圆形,叶柄较短,木质素含量在LeT6中没有显著改变,但在Kn1植物中显著降低。相对于野生型转录水平表明,Kn1中过氧化物酶PRX(peroxidase)基因显著增加,而在Kn1和LeT6中观察到CAD显著减少,表明KNOX基因至少调节木质素生物合成途径中的两个关键步骤(CAD 和 PRX)。肉桂醛(木质素的单体前体)分析表明,LeT6 中的肉桂醛部分没有显著差异,而Kn1茎中的肉桂醛部分相对于野生型显著降低,这可能是由于木质素含量降低造成的。Kn1可能通过改变4CL上游的生物合成步骤的水平来抑制木质化,而CAD表达水平并没有因为这一途径流量的减少而受到限制。
5 其他
5.1 酶
糖基化发生在植物的许多生物过程中,并被认为在一系列植物化合物的生产中起重要作用。糖基转移酶(GTs)是负责植物化合物糖基化的酶。糖基转移酶在维持细胞稳态和调节植物生长,发育以及对压力环境的防御反应中起重要作用[70-71]。糖基转移酶在次级代谢产物的合成,激素的修饰,和细胞壁合成等过程中发挥重要作用。
Wang等[34]将杨树的糖基转移酶基因PtGT1(属于GT1家族)在烟草中过表达,发现在转基因烟草中木质素含量显著增加70%以上,同时还出现明显早花现象。但是GT1调控木质素的机理还不清楚。
5.2 激素
植物激素是调控植物生长发育和抗病抗逆的重要因子,而木质素则是植物生长细胞壁的组成部分和抗病抗逆的重要代谢化合物。因此激素也能调控木质素的代谢合成。其中,赤霉素(Gibberellin, GA)、水杨酸(Salicylates,SA)、茉莉酸(Jasmonates ,JA)、乙烯(Ethylene,ETH)和吲哚乙酸(indoleacetic acid,IAA)等植物激素对木质素合成调控研究较为广泛[1-6]。
Sitbon在烟草中共表达了来自根癌土壤杆菌(A. tumefaciens)的 IAA 生物合成基因35S-iaaM和iaaH,在转基因烟草中IAA含量增加,乙烯含量也显著增加[35]。转基因烟草茎中木质部的木质素含量高于野生型,木质素的组成也发生了变化,G-木质素增加,S/G比值降低。过氧化物酶(POD)活性增加明显,而CAD酶和β-葡萄糖苷酶活性变化不大。POD的mRNA水平增加明显,而POD的mRNA水平增加被认为与与木质素和相关多酚的水平升高有关。他们认为转化体中的高 IAA 水平,通过诱导乙烯合成,增加了 POD 活性,因此也增加了木质素沉积。
6 总结及展望
木质素作为一种重要的代谢化合物,不仅对烟草的生长发育、抗病抗逆性存在影响,也对烟草的感官质量存在重要影响。目前,对于烟草中木质素的调控研究,有着广泛的研究,其主要调控酶研究的较为清楚和透彻,但是其主酶调控基因以及主酶调控基因间相互作用影响研究还有待进一步挖掘完善;木质素单体到木质素的聚合过程机理及基因调控机制还不是很清楚,有待进一步研究;而转录因子以及转录调控网络有一定研究,但是基因和调控机制还需系统研究;其他酶和激素基因对烟草木质素的调控以及调控机理也有待拓展完善;现有技术主要使用的是转基因技术,而随着基因编辑技术的成熟应用,基因编辑技术在烟草木质素中的应用,将有广阔的研究空间。另外,烟草木质素在烟草农业适应性、抗病性能、烘烤性能、香吃味感官性能、物理性能、加工性能、工业适用性能等研究较少,可以进一步拓展相关研究。
