中国南瓜强雌性状的遗传分析
2023-03-01杨建国汪端华吴双花
李 倩 杨建国 汪端华 吴双花 王 鑫
(湖南省蔬菜研究所,湖南省蔬菜工程技术研究中心,湖南长沙 410125)
南瓜(Cucurbita moschataDuch.)为葫芦科南瓜属雌雄同株异花一年生草本植物,在自然群体中主要表现为强雌株、普通性型株、强雄株3 种性型。强雌株主要指主蔓上初现几节雄花,后续连续出现雌花,雌花节率在80%以上的植株;强雄株指雄花多雌花少,雌花节率为30%以下的植株;普通性型株指雌、雄花交替出现,雌花节率在30%以上、80%以下的植株。南瓜为单性花植物,性型表达相对稳定,极少出现两性花,其生长一般先现雄花,之后雌雄花交替出现,极少数特异资源到某一节位后只有雌花产生(Switzenberg et al.,2014)。瓜类作物一般都是雄花产生花粉,雌蕊通过蝴蝶、蜜蜂等昆虫或人工授粉受精后发育成果实,雌花节率在很大程度上决定了植株坐果性和大田产量,进而对经济效益产生影响。瓜类蔬菜育种上,利用强雌系生产杂交种F1时,可免去或减少去雄的工作,节省人力和时间的同时还可以提高杂交种纯度,并且强雌株一般为早熟材料。因此,掌握中国南瓜强雌性状遗传规律,有利于更好地进行中国南瓜强雌性种质创新和新品种选育。
本试验以中国南瓜强雌系和普通性型株系为亲本,通过杂交和回交,构建6 世代群体,春、秋两季连续统计25 节以内的雌花节率,利用多世代联合分析法进行强雌性状遗传分析,旨在解析中国南瓜强雌系遗传规律,为中国南瓜强雌材料创制和新品种选育提供理论基础。
1 材料与方法
1.1 试验材料
供试中国南瓜亲本材料由湖南省蔬菜研究所南瓜课题组提供,F81 为强雌系,M32 为普通性型株系,2 份材料均为遗传稳定的高代自交系。
1.2 试验方法
试验在湖南省农业科学院高桥基地进行。2019年春季,将强雌系F81 与普通性型株系M32 进行杂交,获得F1种子;2019 年秋季,Fl自交,并以F81 和M32 分别作为亲本进行回交,获得F2、B1、B2种子。6 世代材料于2020 年秋季和2021 年春季分别种植于大棚和露地,用于雌花节率调查、分析。
性型样本调查容量:P1和P2为15 株,F1为45 株,F2> 200 株,B1> 85 株,B2> 120 株。
性型调查方法:为保证调查结果的可靠性,整个性型调查分2 个阶段,第1 个阶段为植株长到15 节左右,第2 个阶段为植株长到25 节以上时,最后统计25 节内的雌花节率。
1.3 统计分析
6 世代的基本参数由Excel 软件进行统计。遗传分析运用曹锡文等(2013)研制的SEA 软件包,采用植物数量性状混合遗传多世代联合分析方法进行分析。各世代、各成分分布的参数利用极大对数似然法、IECM 算法,通过赤池信息量准则(AIC)判别候选模型,并进行5 项检验(U12、U22、U32、nW2、Dn),选择最优遗传模型,估算一阶和二阶参数。
2 结果与分析
2.1 中国南瓜6 世代群体强雌性状的差异分析
从表1 可以看出,中国南瓜6 世代群体春、秋两季的雌花节率各参数值差异明显,各世代材料两季中F2、B1、B2群体极差与变异系数均高于P1、P2、F1群体,说明3 个分离世代群体材料离散度高,变异幅度较大,存在较高的遗传多样性。
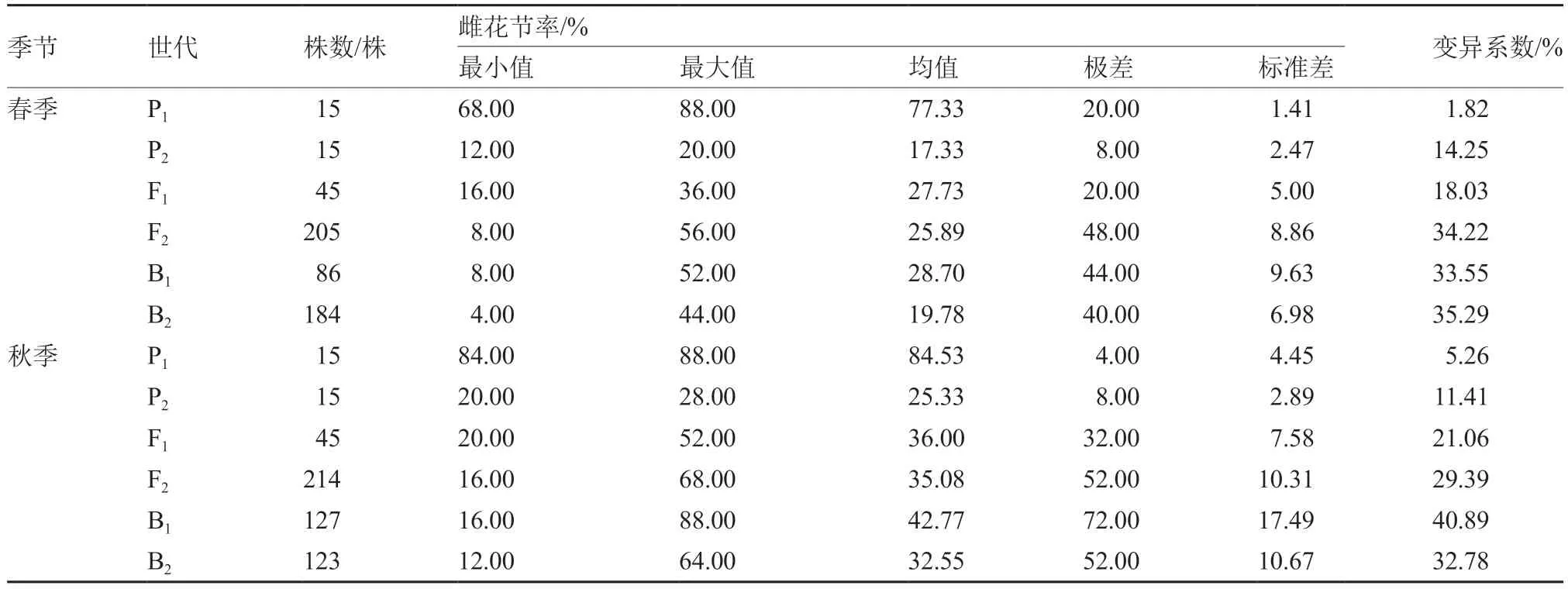
表1 中国南瓜6 世代群体雌花节率的基本参数
2.2 中国南瓜6 世代群体雌花节率的频次分布
由图1、2 可知,中国南瓜6 世代群体雌花节率的频次分布春、秋两季差异明显。春季,3 个分离世代偏普通性型亲本(P2)和中间类型;秋季,3 个分离世代离散度较高,B1出现了部分偏强雌亲本(P1)现象。另外,3 个分离世代群体频次分布均呈连续分布状态,并表现出偏正态分布的单峰。
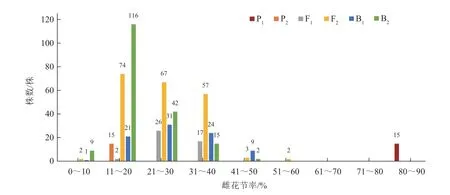
图1 中国南瓜6 世代群体春季雌花节率的频次分布
2.3 中国南瓜强雌性状遗传模型及适合性检验
中国南瓜强雌性状遗传模型多世代联合分析结果表明(表2),春、秋两季的AIC 值较小的模型有C-1(3 383.170/3 463.027)、C-2(3 396.193/3 464.271)和C-3(3 397.110/3 475.772),其中最小的为C-1 模型,即1 对主基因,同时存在多基因效应。
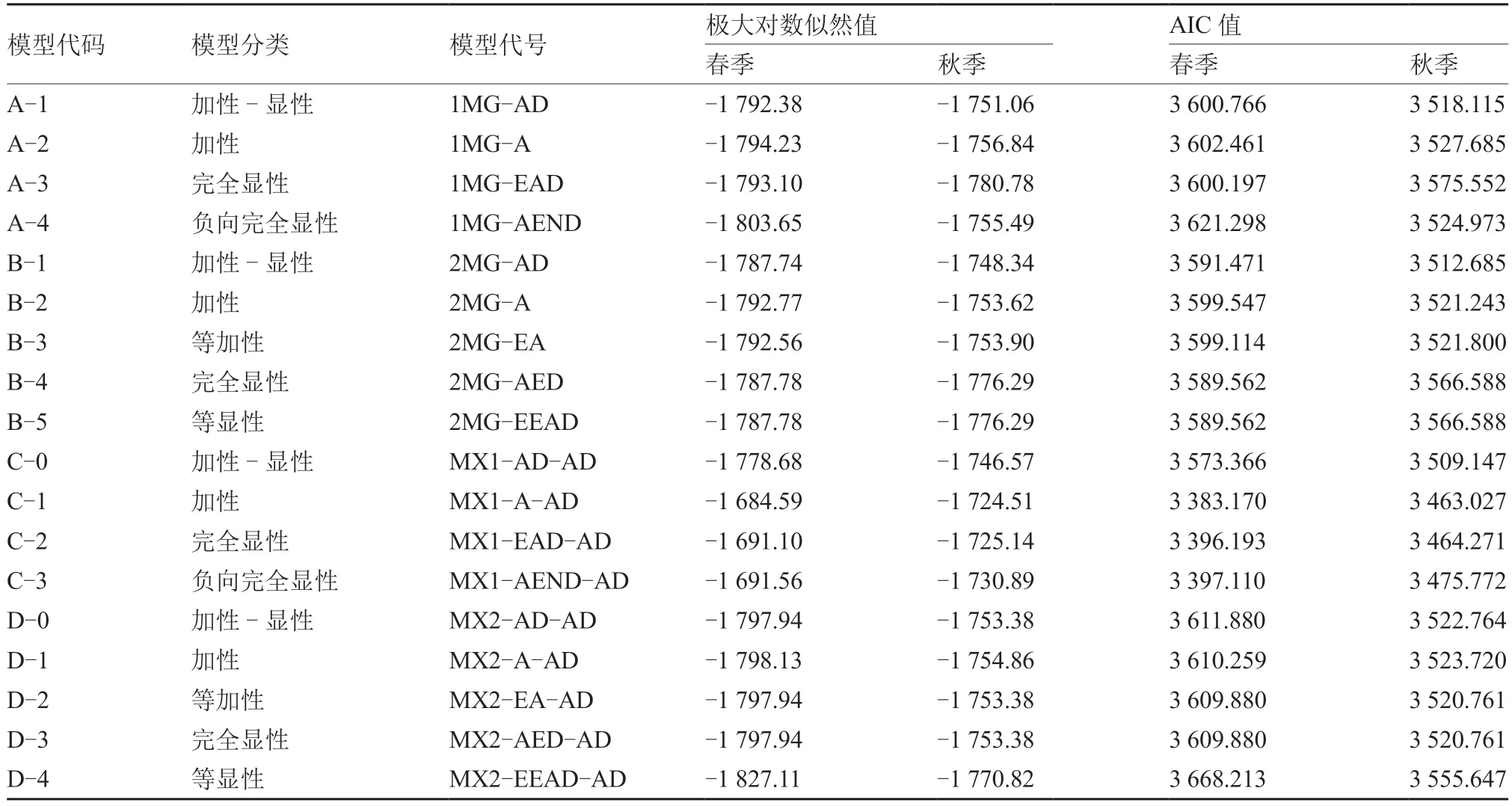
表2 中国南瓜强雌性状6 世代群体各种模型的极大对数似然值和AIC 值
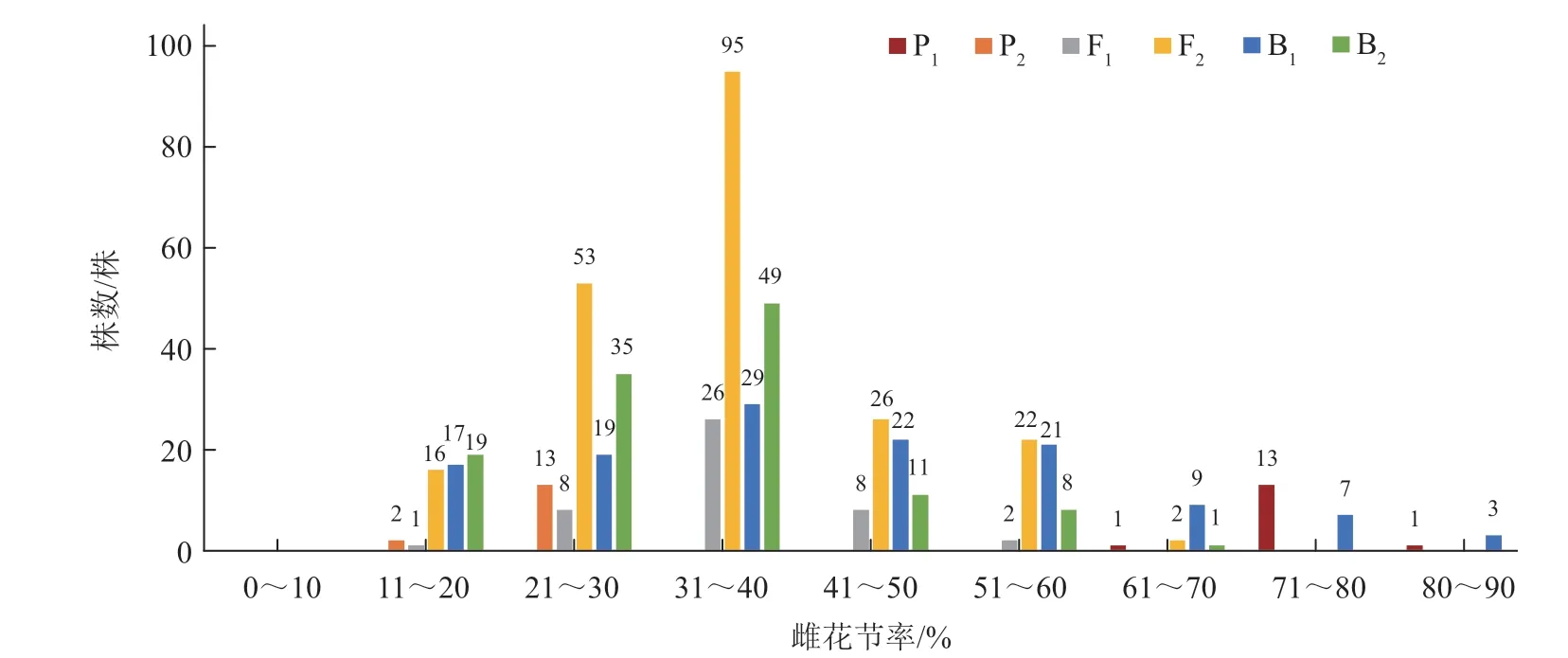
图2 中国南瓜6 世代群体秋季雌花节率的频次分布
进一步对C-1 模型进行适合性检验,结果显示(表3),春季6 世代群体在0.05 水平上有14 个统计量达到显著差异,在0.01 水平上有2 个统计量达到极显著差异;秋季6 世代群体在0.05 水平上有16 个统计量达到显著差异,在0.01 水平上有3 个统计量达到极显著差异。
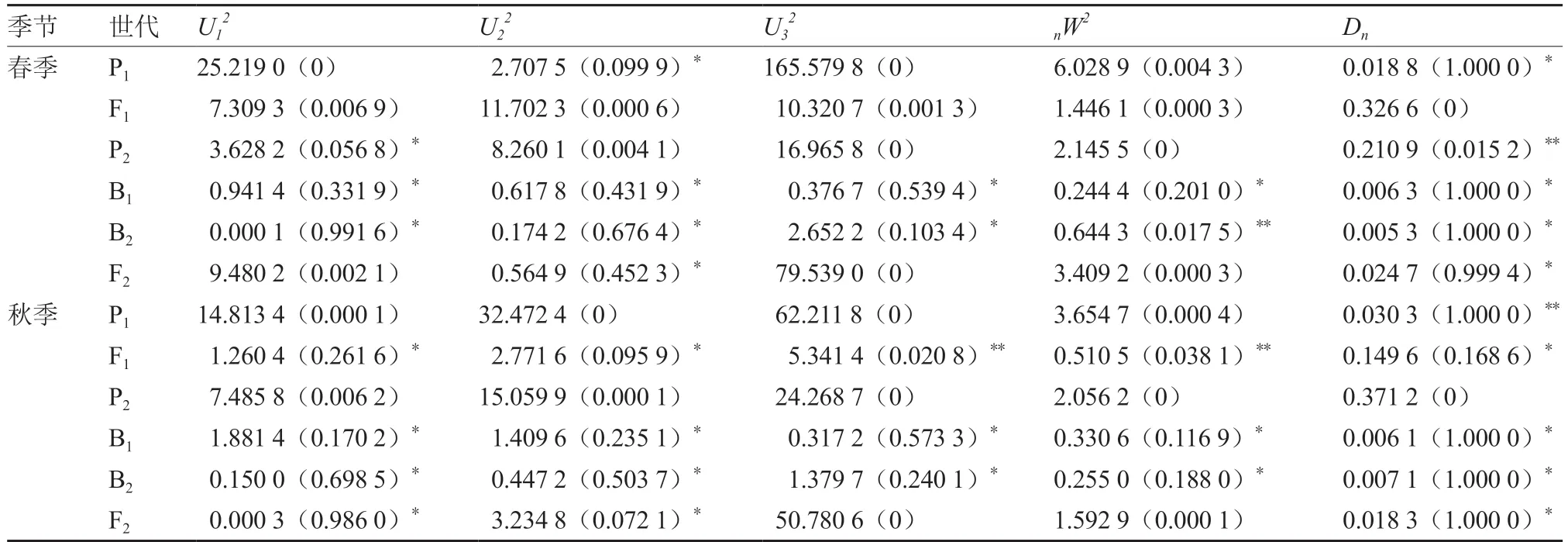
表3 C-1 模型的适合性检验
2.4 C-1模型的遗传参数估计
C-1模型的遗传参数估计结果显示(表4),中国南瓜强雌性状由1 对主基因+多基因控制,并且第1 主基因存在加性效应。春、秋两季的6 世代群体平均值分别为16.521 2、40.598 5,第1 主基因的加性效应值分别为4.644 2、4.136 7,多基因的加性效应值分别为4.320 0、10.246 2,均为正向作用;而多基因的显性效应值分别为12.521 2、-6.873 2。春季,多基因的显性效应值大于加性效应值,而秋季则相反。在分离世代B1、B2、F2中,春季主基因遗传率分别为4.38%、8.56%、0,在B2世代遗传效率较高;秋季3 个分离世代主基因遗传率仍在B2世代中最高(2.33%)。两季分离世代中多基因的方差和遗传率均为0,说明多基因对3 个分离世代的影响微效,可忽略不计。
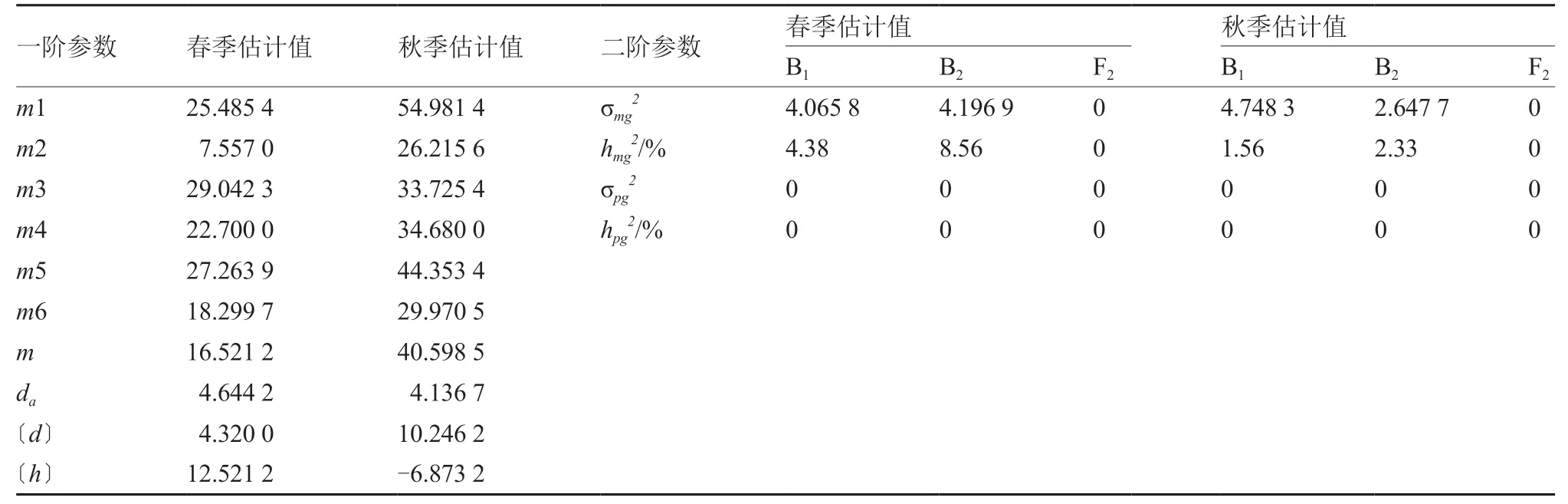
表4 C-1 模型一阶和二阶遗传参数
3 讨论与结论
瓜类作物的熟性和产量受雌花节率影响较大,雌性性状是瓜类最重要的育种目标性状之一(李海真 等,2021)。单文琦(2016)研究表明,印度南瓜雌性性状遗传规律符合主基因+多基因模型,控制印度南瓜雌性性状的主基因为单隐性核基因。赵志伟等(2012)研究发现,苦瓜强雌性状受1 对主基因控制,且受多基因影响。邹晓艳(2007)研究也证实,黄瓜全雌性和强雌性均由1 对主基因控制,伴随微效多基因作用,并且全雌性组合的主基因遗传率略高于强雌性组合。本试验结果表明,中国南瓜强雌性状遗传规律与印度南瓜、苦瓜、黄瓜等结果相似,但与陈清华等(2002)在节瓜纯雌系5 个世代花性研究发现的遗传规律(节瓜花性表达受2 对独立分离基因调控,纯雌系作用基因为2 对纯合基因,全雄系作用基因为1 对杂合、1 对纯合基因)不同。以上研究表明,葫芦科蔬菜在性型遗传规律上有相似之处,但也存在一定差异,反映了同科作物间的亲缘关系远近及遗传的多样性。
本试验采用多世代联合分析法对中国南瓜强雌性状的遗传规律进行分析,结果发现该性状符合C-1 模型,进一步对最适模型进行适合性检验和遗传参数估值,发现第1 主基因和多基因的加性效应值在春、秋两季均为正向作用,而多基因的显性效应值在秋季为负向作用,两季中主基因的遗传率均占主要作用,而多基因作用可忽略不计。表明,中国南瓜强雌性状以第1 主基因为主,且受环境影响,该结果与梁少华(2020)在西瓜性型上的研究结果相似。同样,Lai 等(2018)连续5 年对黄瓜核心种质进行雌性季节变化研究,结果表明71.3%的材料在早秋与春季相比,会显著降低雌性节率。厉建梅等(2011)在黄瓜雌性性状上的遗传分析也表明,黄瓜雌花节率除了由基因的加性效应控制外,还受到环境(季节)的影响。
综上,中国南瓜强雌性状的遗传符合1 对主基因+多基因模型,与大多数瓜类蔬菜性型遗传模型一致。但中国南瓜雌花节率受环境影响较大,下一步将结合不同环境条件,利用基因组学手段对强雌性状调控机理进行研究。
