微生物共利用木糖和葡萄糖生产化学品研究进展
2023-03-01王川东张君奇刘丁源马媛媛李锋宋浩
王川东,张君奇,刘丁源,马媛媛,李锋,宋浩
(1 天津大学化工学院,天津 300072;2 天津大学合成生物学前沿科学中心和系统生物工程教育部重点实验室,天津 300072;3 天津大学(青岛)海洋工程研究院有限公司,山东 青岛 266237;4 天津大学海洋科学与技术学院,天津 300072)
木质纤维素生物质主要由纤维素、半纤维素和木质素构成,常见于农作物秸秆、杂草、木材废料及其他固体废弃物中,是储量丰富、廉价易得的可再生资源[1−2]。植物光合作用产生的生物质大部分为木质纤维素类,其资源化开发有“不与人争粮,不与粮争地”的优势,可用于工业生产醇类、微生物油脂和有机酸等化学品。与纤维素水解生成的葡萄糖相比,半纤维素水解产生的木糖更难被微生物利用,例如酿酒酵母(Saccharomyces cerevisiae)能很容易地利用纤维素水解生成的葡萄糖发酵产乙醇,但难以利用木糖[3−4],这极大地限制了木质纤维素资源的利用。因此,构建木糖−葡萄糖共利用工程菌株是解决该问题的有效方法之一。相比原核生物,真核生物戊糖代谢途径涉及复杂的氧化还原反应,需要更多的酶及辅因子参与,使得在酵母中构建高效戊糖代谢途径难度较大[5]。因此,木糖及葡萄糖共利用宿主菌的开发有待进一步进行。目前,除酵母外,木糖−葡萄糖共利用菌株的开发已经在酪丁酸梭菌(Clostridium tyrobutyricum)、少根根霉(Rhizopus arrhizus)、大肠杆菌(Escherichia coli)等多种微生物中被报道[6−8]。本文主要综述了近年来木糖及葡萄糖共利用工程菌株的构建策略及在醇类、微生物油脂、γ−聚谷氨酸和有机酸合成领域的进展,总结了限制木糖−葡萄糖共利用效率的主要瓶颈及其解决方法,为提高木质纤维素生物质资源化利用和可再生能源化学品的产业化合成提供参考。
1 天然微生物木糖及葡萄糖共利用现状
自研究发现细菌、酵母菌能发酵木糖生产乙醇以来,现已知有两百多种微生物可以代谢木糖,包括酵母菌、新月柄杆菌(Caulobacter crescentus)、假单孢杆菌(Pseudomonas)、枯草芽孢杆菌(Bacillus subtilis)及大肠杆菌等。其中既能利用木糖又能利用葡萄糖的酵母菌是研究重点,主要包括树干毕赤酵母(Schefersomyces stipites)、嗜鞣管囊酵母(Pachysolen tannophilus)、马尔萨尼克鲁维酵母(Kluyveromyces marxianus)、休哈塔假丝酵母(Candida shehatae)、产朊假丝酵母(Candida utilis)及丝状假丝酵母(Candida lusitaniae)等[9−12]。
天然微生物的底物偏好性会影响木糖−葡萄糖共利用发酵性能。Saravanan等[13]对比了汉逊达布氏酵母 (Dabaryomyces hansenii var hansenii)、吉利蒙假丝酵母(Candida guillermondii)和嗜鞣管囊酵母(P.tannophilus)以玉米芯、稻草和小麦秸秆为原料生产木糖醇的动力学过程。利用Logistic 细胞生长模型、底物利用动力学模型及Luedeking−Piret产物生成模型分析,发现P.tannophilus发酵玉米芯水解液时,木糖醇产量更高,说明木糖醇产量受底物利用效率影响。不同来源的木质纤维素水解液中糖含量的差异是影响底物利用效率的关键,汤斌等[14]发现C.shehataeTZ8−13 利用木糖及葡萄糖共发酵产乙醇时,葡萄糖被优先利用,且木糖和葡萄糖质量比为1∶1时乙醇产量较高。Agbogbo 等[15]发现树干毕赤酵母(S.stipites)的葡萄糖利用也优先于木糖,并且葡萄糖大部分供给细胞生长,木糖则主要用于产物合成。该研究表明,水解液中混糖比例的不同会影响底物利用效率及乙醇产量,因此水解液的高效利用需要底物适应能力更强的菌株。
研究表明,S.stipites不仅可以代谢木糖和葡萄糖,还能代谢甘露糖、半乳糖、纤维二糖、甘露聚糖及木聚糖低聚物等[16−19],工业应用价值较高。但是,S.stipites对乙醇、弱酸及糠醛等抑制剂耐受度低,限制了其发酵性能,采取适当的原料预处理和脱毒方法有助于促进水解液糖化和去除抑制剂[20−21]。例如,利用稀硫酸处理糖化后的玉米秸秆水解液,补加氢氧化铵中和水解液中的酸后,使得乙醇转化率从0.32g/g 提高至0.44g/g(乙醇/木糖和葡萄糖)[22]。但是,能高效共利用木糖及葡萄糖发酵制备化学品的天然菌株较少,且普遍存在底物偏好性强、碳分解代谢物阻遏(carbon catabolite repression, CCR)、产量低等问题[14−15,18],因此,构建底物谱更广、混合糖利用率更高的木糖−葡萄糖共利用工程菌株是产业化应用的必然要求。
2 木糖及葡萄糖代谢途径简介
2.1 木糖代谢途径
研究发现,具备木糖代谢能力的微生物体内一般有4种木糖代谢途径,包括木糖氧化还原酶途径(XR 途径)、异构酶途径(XI 途径)、Weimberg 途径(WBG途径)和Dahms途径[23−26],见图1。此外,近期在E. coli中建立了木糖−1−磷酸酯(X−1−P)及核糖−1−磷酸酯(R−1−P)途径,实现了二碳化合物的合成。其中,原核和真核生物中木糖利用能力较强的菌株E.coli[27]和S.stipites[18]分别借助XI 和XR途径代谢木糖。在XI途径中,木糖首先被XylA基因编码的木糖异构酶(xylose isomerase, XI)转化为木酮糖,随后被XylB编码的木酮糖激酶(xylulokinase, XK)磷酸化,产生5−磷酸木酮糖进入磷酸戊糖途径(pentose phosphate pathway, PPP)[23]。
类似于XI 途径,在XR 途径中,木糖先被NAD(P)H 依赖的木糖还原酶(xylose reductase, XR,Xyl1基因编码)转化为木糖醇,进一步在木糖醇脱氢酶(xylose dehydrogenase, XDH,Xyl2 基因编码)作用下转化为木酮糖,后被XK(Xks1 基因编码)催化生成5−磷酸木酮糖,进入PPP,见图1。XR及XI 途径是构建木糖利用工程菌株的常用途径。例如,Sekar 等[28]利用定向进化激活了电活性微生物希瓦氏菌(Shewanella oneidensis)的木糖代谢能力。Li 等[29]采用合成生物学策略,在希瓦氏菌(S.oneidensis)中异源过表达XR 及XI 途径相关基因,获得了能够利用木糖为碳源生长产电的工程希瓦氏菌株,克服了S.oneidensis不能利用五碳糖的限制。

图1 木糖及葡萄糖共利用代谢途径与发酵产品
相比于前两种代谢途径,来自于新月柄杆菌(C.crescentus)的WBG途径各反应步骤有较大的热力学标准吉布斯自由能负向差值(表1),使得各步反应更容易自发进行;且终产物为α−酮戊二酸(AKG),可以绕过PPP 直接进入三羧酸循环(TCA),代谢流程相对较短;此外,该途径不产生二氧化碳,没有碳损失[30−32]。WBG 途径中D−木糖经五步酶促反应被氧化成AKG(图1),D−木糖首先在木糖脱氢酶(CcXylB基因编码)的作用下被氧化成D−木糖−γ−内酯,然后在基因XylC编码的D−木糖−γ−内酯酶(xylonolactonase, XLA)催化下转化为中间体D−木糖酸盐,进而在D−木糖酸脱水酶(D−xylonate dehydratase, XAD,XylD基因编码)和2−酮−3−脱氧−D−木糖酸脱水酶(KDX dehydratase,KDXD,XylX基因编码)参与下进行两次脱水反应,产生2−酮−3−脱氧−D−木糖酸(2−keto−3−deoxy−D−xylonate, KDX),之后生成α−酮戊二酸半醛(α−ketoglutarate semialdehyde, KGSA)。最后,被α−酮戊二酸半醛脱氢酶(KGSA dehydrogenase, KGSADH,CcXylA基因编码)以NAD(P)+依赖的方式氧化成AKG,进入TCA[25,30−32]。因此,碳流进入细胞中枢代谢的节点不同于XI 和XR 途径。目前,在枯草芽孢杆菌(B.subtilis)、E.coli、恶臭假单孢杆菌(Pseudomonas putida)和谷氨酸棒杆菌(Corynebacterium glutamicum)等宿主菌中构建该途径,获得了D−1, 2, 4−丁三醇(BT)、γ−聚谷氨酸(γ−PGA)、绿脓菌素、鼠李糖脂、乙醇酸盐、木糖酸盐、AKG、L−鸟氨酸等产品[31−37]。

表1 Weimberg途径标准吉布斯自由能变化[30]
发现于Pseudomonas的Dahms途径与WBG途径前半部分相同,不同的是反应进行到KDX 之后转化为乙醇醛和丙酮酸,见图1。
X−1−P 和R−1−P 途径能在一定程度上绕过复杂的内源PPP,直接裂解木糖[38−39],具有合成二碳化合物的天然优势[40−41]。两条途径中木糖均先被异构化为木酮糖,X−1−P途径中木酮糖被磷酸化为木酮糖−1−磷酸,之后经醛解生成乙醇醛和二羟丙酮磷酸(dihydroxyacetone phosphate, DHAP)。不同的是R−1−P 途径中木酮糖差向异构化形成核糖,再进行磷酸化和醛解转化为乙醇醛和DHAP。
2.2 葡萄糖代谢途径
葡萄糖作为大多数生物的主要碳源,在微生物体内的代谢途径主要有EMP 途径(Embden−Meyerhof−Parnas pathway, EMP, 又称糖酵解)、HMP途径(hexose monophophate pathway, HMP),ED 途径(entner−doudoroff pathway, ED) 及PK 途 径(phospholytic ketase pathway, PK)[42],见图1。其中EMP 途径是微生物中普遍存在的代谢途径。在无氧条件下葡萄糖被分解为丙酮酸,在此期间每分解1分子葡萄糖产生2分子丙酮酸及2分子ATP。
在微生物HMP 途径中,葡萄糖经过几步氧化反应产生核酮糖−5−磷酸和CO2,核酮糖−5−磷酸经同分异构化或差向异构化产生核糖−5−磷酸或木酮糖−5−磷酸,在无氧情况下发生碳架重排,产生己糖磷酸和丙糖磷酸。该途径是许多微生物木糖代谢的必经之路。
ED 途径是一种EMP 途径的替代途径,在研究Pseudomonas时发现,存在于一些缺乏完整EMP 途径的微生物中。ED 途径为微生物特有,海洋菌中较多[43],在革兰氏阴性菌中分布较广,如嗜糖假单孢菌(Ps.saccharophila)、铜绿假单孢菌(Ps.aeruginosa)、林氏假单孢菌(Ps.lindneri)、荧光假单孢菌(Ps.fluorescens)、真养产碱菌(Alcaligenes eutrophus)、运动发酵单孢菌(Zymomonas mobilis)等。该途径只需四步反应即可快速代谢葡萄糖,获得由EMP 途径经十步反应才能形成的丙酮酸,但产能水平较低,1 分子葡萄糖分解为2 分子丙酮酸时,只净得1分子ATP和1分子NADH[42−43]。在葡萄糖促进因子(glf基因编码)协助下葡萄糖被转运至胞内,在葡萄糖激酶(glk基因编码)催化下被氧化成葡萄糖−6−磷酸(G−6P),同时消耗1 分子ATP,然后在葡萄糖−6−磷酸脱氢酶(Zwf基因编码)作用下脱氢转变为葡萄糖−1, 6−二磷酸(1G−6P),1G−6P 被磷酸葡萄糖酸脱水酶(edd基因编码)催化脱水生成2−酮−3−脱氧−6 磷酸葡萄糖酸(KDPG),进而在KDPG醛缩酶(eda基因编码)作用下转化为甘油醛三磷酸和丙酮酸,在有氧条件进入TCA 或进行无氧乙醇发酵,见图1。ED 途径代谢流程较少,在微生物宿主中构建更容易,是木糖−葡萄糖共利用工程菌株发酵生产化学品的重要途径。
磷酸解酮酶途径是明串珠菌在异型乳酸发酵中分解已糖和戊糖的途径。该途径的特征酶是磷酸解酮酶,根据解酮酶的不同,具有磷酸戊糖解酮酶的称为PK 途径,具有磷酸己糖解酮酶的称为HK途径。
2.3 木糖及葡萄糖共利用代谢途径
构建木糖及葡萄糖共利用工程菌株是提高木质纤维素原料资源化利用效率的重要方法,可用于醇、油脂和有机酸等多种化学品的生物合成,见图1。EMP 或ED 途径是基因工程菌株构建葡萄糖代谢模块的常用途径,而木糖XR、XI及WBG途径则常被用来与葡萄糖代谢途径组合,构建木糖−葡萄糖共利用工程菌株。野生型酵母可以利用葡萄糖,且具有抗逆性强、培养周期短、遗传操作简单等优点,可作为底盘菌,用于开发木糖−葡萄糖共利用工程菌株。但是,复杂的氧化还原反应和酶催化系统增加了在酵母中构建异源戊糖或己糖代谢途径的难度。因此,研究者将木糖−葡萄糖共利用研究扩展至原核及其他真核底盘菌,如E.coli、C.tyrobutyricum、C.glutamicum、B.subtilis、Pseudomonas及少根根霉(Rhizopus arrhizus)等。目前在乙醇、BT、木糖醇及丁醇等醇类合成,乳酸、富马酸[44]、酮酸[45]、三羟基丙酸(3−HP)[46]、丁酸、柠檬酸等有机酸及γ−PGA、微生物油脂生产中成功应用,见表2。
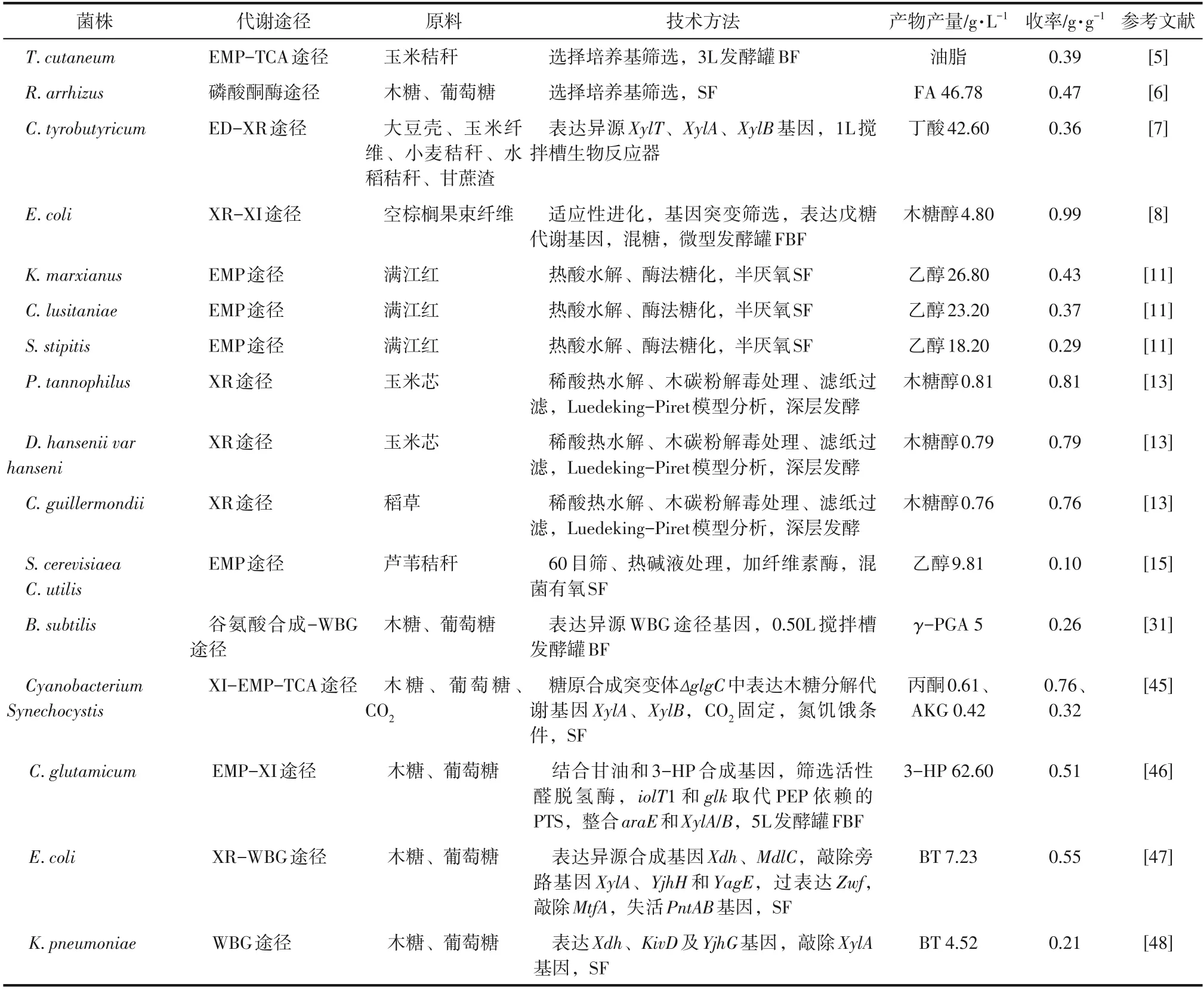
表2 微生物木糖及葡萄糖共利用生产化学品
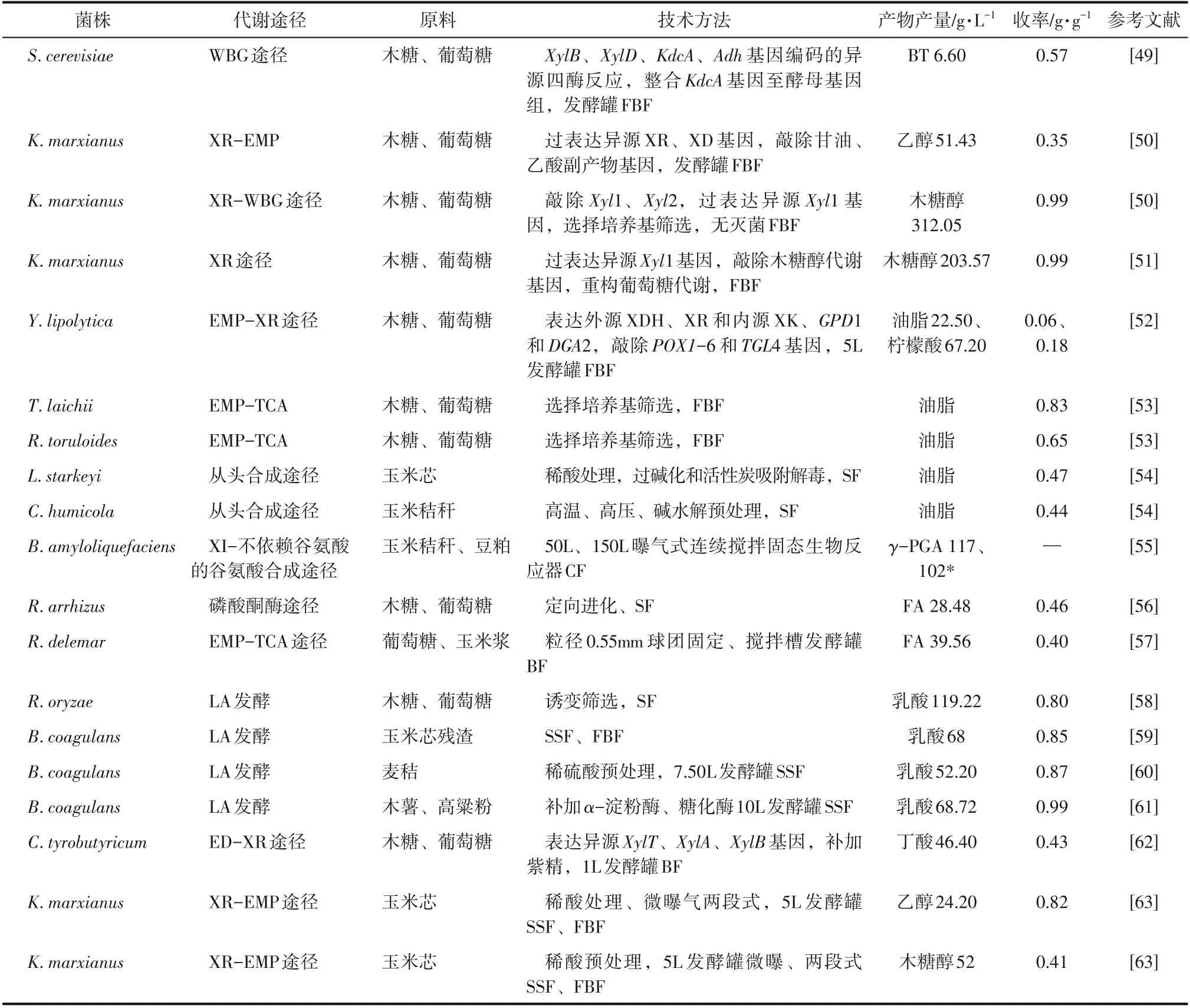
续表2
3 木糖−葡萄糖共利用发酵产品合成
3.1 醇类
3.1.1 乙醇
第二代生物乙醇的制备是木质纤维素最具前景的应用领域。酵母菌因细胞壁厚、营养要求低、体积大,在乙醇发酵中更具有优势[17]。S.cerevisiae和S.stipites因能利用多种碳水化合物发酵产生大量乙醇,且便于遗传操作、乙醇耐受性强,使其成为极具潜力的乙醇发酵菌株。例如,S.stipites能够利用多种木质纤维素生产乙醇[64],S.cerevisiae可以代谢木质纤维素水解液中的多种糖[65]。但S.cerevisiae缺乏木糖利用能力,因此,将木糖代谢途径引入S.cerevisiae是构建木糖−葡萄糖共利用工程菌的可行方法。Wahlbom 等[66]将来自S.stipitis的Xyl1 基因和里氏木霉(Trichoderma reesei)的Xyl2 基因转入S.cerevisiae中,经甲基磺酸乙酯诱变筛选后,得到的重组菌可以利用木糖生产少量乙醇,但转化率仅为0.25g/g(乙醇/木糖),且副产物木糖醇较多。Shi 等[67]将C.shehatae的Xyl1 和Xyl2 基 因 克 隆 到S.cerevisiae中,也存在木糖代谢能力差、副产物较多的问题。为了进一步疏通木糖代谢流、减少副产物,杜仁鹏等[10]选择发酵性能优良的S.cerevisiaeW5和具有木糖代谢能力的C.shehatae为原始菌株,利用原生质融合和基因工程育种方法,克隆Xyl1基因,提高了葡萄糖及木糖共利用产乙醇的效率。
除了高效的糖代谢途径外,宿主菌的抗逆性与底物适应性也是影响糖代谢效率的关键。K.marxianus因具有耐高温和可代谢多种底物的优点,有较高的应用价值[68]。洪炯课题组[69−71]首次鉴定了K.marxianus木糖代谢途径的3 个关键酶——木糖脱氢酶、XK 及XDH,推动了K.marxianus在木糖发酵领域的应用。韩锡铜等[72]对比了K.marxianus和S.cerevisiae6525利用不同碳源的情况,筛选出高效的戊糖发酵菌株K.marxianusDL1,相比S.cerevisiae6525,K.marxianusDL1 利用己糖(葡萄糖、甘露糖、半乳糖)和戊糖(木糖、阿拉伯糖)生长更快、乙醇产率更高,在20g/L 糖条件下,K.marxianusDL1 的细胞量及乙醇产量分别是S.cerevisiae6525 的 近2 倍 和1.70 倍。Chupaza 等[11]对 比 了K.marxianus、C.lusitaniae、S.stipitis及S.cerevisiae利用丝状满江红产乙醇的过程。除S.cerevisiae外,其他三种酵母菌都能利用葡萄糖和木糖混合物,且K.marxianus产量及转化率最高,分别为26.80g/L 和0.43g/g(乙醇产量/初始单糖)。其次是C.lusitaniae产量为23.20g/L,转化率为0.37g/g。S.stipitis因抗逆性较差,产量及转化率相对较低,分别为18.20g/L 和0.29g/g。S.cerevisiae只能利用混合物中的葡萄糖,而不能利用木糖,导致乙醇产量最低,仅为13.7g/L,转化率为0.22g/g。这表明菌株的抗逆性和底物偏好影响着底物利用效率和乙醇产量。
3.1.2 D−1, 2, 4−丁三醇
D−1, 2, 4−丁三醇(BT)是一种具有三个亲水羟基的四碳多元醇,是重要的非天然精细化学品,在医药、军工、化妆品等领域应用广泛[73−76]。相比传统的化学合成法,生物法反应条件温和、原料廉价易得、污染小,更符合绿色循环发展的要求。以木糖为底物的四步生化反应是目前最高效的BT 生物合成路线(图2),研究较多的宿主菌为E.coli、K.pneumoniae、S.cerevisiae、Ps.fragi等。Cao 等[33]在E.coli中共表达来自C.crescentus的CcXylB、XylC基因,E.coli的木糖脱水酶基因YjhG、醛还原酶基因AdhP以及P.putida的2−酮酸脱羧酶基因MdlC,同时敲除XylA和XylB基因以减少木糖分解代谢分流,构建的工程菌株以20g/L 木糖为原料,补料分批发酵BT 产量为3.92g/L,摩尔转化率为27.7%(BT/木糖)。
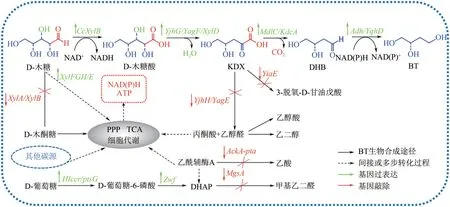
图2 木糖及葡萄糖共利用合成D−1, 2, 4−丁三醇(BT)代谢途径及其代谢旁路[74−77]
生物法合成BT 存在合成途径活性较低、副产物多及忽略宿主生长对BT 合成的影响等问题。王金保[73]从合成途径、辅因子和底物供给三个层面优化,在E.coliMG1655 中表达异源CcXylB和MdlC,敲除XylA和2−酮酸醛缩酶基因YjhH和YagE,适量过表达Zwf,敲除MtfA,失活转氢酶基因PntAB使辅因子供应平衡,成功构建木糖及葡萄糖共利用工程菌BT−02,BT 产量达7.23g/L,摩尔转化率为55%(BT/木糖),首次确定了PPP 是为BT 合成供给NADPH的主要途径。
研究发现,敲除XylA、XylB、YagE及YjhH基因可以削弱木糖分支代谢和副产物途径,有利于碳流向BT合成途径[74]。孙雷等[75]筛选获得了来自乳酸乳球菌(Lactococcus lactis)的2−酮酸脱羧酶(KivD基因编码),大幅提高了3−脱氧−D−甘油−戊酮糖酸的脱羧反应速率。优化后BT 合成效率虽有提高,但游离质粒诱导表达成本较高,且CCR 使细胞无法同时利用木糖和葡萄糖高效积累目的产物。改造葡萄糖磷酸转移酶系统(phosphotransferase system,PTS)可有效缓解CCR 现象[76]。针对上述限制因素,诸葛斌课题组[47,77]以缺失YagE、YjhH的菌株为基础,通过Red系统将外源基因KivD、CcXylB整合至E.coli基因组的XylA、XylB、ptsHI、ptsG、crr位点,利用廉价的乳糖替代IPTG 诱导表达,构建的工程菌E.coliW031 可利用木糖−葡萄糖混合糖(30g/L 木糖、10g/L 葡萄糖)发酵合成BT,产量为3.90g/L,转化率为30%,减少了副产物分流和质粒表达成本,为后续规模化应用打下了基础。
CCR 限制了以E.coli为宿主开发木糖−葡萄糖共利用工程菌合成BT 的研究。K.pneumoniae具有木糖及葡萄糖混合糖利用效果好、CCR 较弱、生长速度快等优点。因此,以K.pneumoniae为宿主构建合成BT 的工程菌,可提高混合糖利用能力。李玉石等[78]将来源于C.crescentus的CcXylB和L.lactis的KivD基因及E.coliW3110 的YjhG基因克隆至K.pneumoniaeZG25,并敲除了XylA减少碳分流,构建的工程菌发现增加葡萄糖能够提高生物量、减少木糖酸积累,以30g/L 木糖和10g/L 葡萄糖为底物共发酵时,BT 产量达4.52g/L,摩尔转化率为21%(BT/木糖)。
此外,Yukawa等[79]利用C.crescentus的CcXylB、XylD及L.lactis的KdcA和Adh基因编码的异源四酶催化反应,由木糖合成BT,将KdcA基因的多个拷贝整合到酵母基因组,增强工程菌性状稳定性。但是,异源BT合成途径的引入会导致NADH/NADPH失衡或缺乏,因此过表达S.cerevisiae的Pos5 基因(编码NADH 激酶)缓解了NADH/NADPH 不平衡问题,用低浓度的葡萄糖和木糖支持NADH 供应,工程菌以木糖为原料,BT 产量为6.60g/L,摩尔转化率为57%(BT/木糖)。可见,在构建重组高效BT 合成路径过程中,应重视宿主菌的自身生长和辅酶平衡问题。
3.1.3 木糖醇
木糖醇是一种五碳糖醇,常作为甜味剂,应用于食品、医药及化工等领域。生物转化法能够克服传统方法的污染重、资源消耗大的缺点,且反应条件更易控制[48,80]。
CCR作用导致野生型E.coli不能同时利用木糖和葡萄糖[49],限制了木糖醇产量,但E.coli能利用廉价培养基快速生长、遗传操作简单,依然具有研究价值[81]。Kim 等[8]将E.coliMG1655 经适应性进化获得了缺失阿拉伯糖转录调节因子(araC)的突变菌株GX50,其中木糖转录激活因子基因XylR、木糖ABC 转运体基因XylFGH及木糖XI 途径基因XylA、XylB组成型表达。此外,还获得了四个额外的突变,包括XylA和乳清酸磷酸核糖基转移酶pyrE基因间突变、编码阿拉伯糖和质子转运体的araE和十一碳烯基−二磷酸酶活性的ybjG基因的错义突变。最终,菌株GX50能够利用空棕榈果束纤维水解物中的木糖和葡萄糖生产木糖醇,转化率为0.99g/g(木糖醇/木糖)。
相比而言,利用酵母发酵生产木糖醇产量更高,更适合规模化生产。Saravanan 等[13]以玉米芯、稻草和小麦秸秆为原料,对比了D.hansenii、P.tannophilus和C.guillermondii生产木糖醇,发现P.tannophilus利用玉米芯水解液发酵产量较高。为了提高木糖代谢效率和木糖醇产量,张佳[50]过表达来自脉孢霉和S.stipites的Xyl1基因,以及糖转运相关基因KmFPS、CiGXF1、CiGXS1,同时敲除了K. marxianus自身的Xyl1 和Xyl2 基因,木糖和葡萄糖共利用发酵获得了312.05g/L 木糖醇。为了进一步 克 服CCR,Zhang 等[51]过 表 达S.cerevisiae的GAL2N376F和粗链孢霉的Xyl1 基因,进一步敲除GPD1、KU70、PGI1和Xyl2基因,重构了葡萄糖代谢途径,阻断木糖醇下游分解代谢路径,使得构建的工程菌株K.marxianusYZB194 可同时利用70g/L葡萄糖和140g/L 木糖,产生139.96g/L 木糖醇。当培养温度调至42℃进行补料分批发酵时,产量进一步提高至203.57g/L,转化率为0.99g/g(木糖醇/木糖)。该研究表明,采用适当的发酵方式,同时增强工程菌合成基因表达水平、阻断产物分解代谢路径,有利于提高木糖醇的产量。
3.2 微生物油脂
微生物油脂的主要成分为C16和C18系脂肪酸,是构成和维持生命活动的基本物质,在食品、能源领域应用广泛,也被用来制备生物柴油[52,82]。常由酵母、霉菌、细菌和藻类等利用碳水化合物或碳氢化合物合成,产油微生物生成并储存的油脂一般占其生物总量的20%以上[83−85],微生物生产油脂的过程分为菌体增长和油脂积累两个阶段[54,86]。微生物生产油脂相比传统油脂制备方法,具有细胞增殖速度快、生产周期短、原料丰富的优势。此外,使用融合、诱变、定向进化及合成生物学等技术优选和调控,还可生产特定的功能性油脂[53,82]。在多种产油微生物中,产油酵母产油能力较强(20%~70%)且易于基因改造,比微藻可用碳源更丰富,比霉菌需氧量更少、重金属离子耐受性更强,比细菌菌体更大、油脂易提取[53,87]。因此,是极具潜力的产油微生物。
产油酵母对木质纤维素水解产物中不同糖分的利用偏好不同,因此,促进各种糖类同时、快速、充分利用对木质纤维素向油脂的转化至关重要[84]。张艳芬[53]采用两段发酵模式研究了赖巴克丝孢酵母(Trichosporon laichii)、圆红冬孢酵母菌(Rhodosporidium toruloides)和斯达氏油脂酵母菌(Lipomyces starkeyi)在不同初始碳源浓度下的产油特性,发现在木糖、葡萄糖及混合糖中,经筛选获得的菌株T. laichiiIEM−14 可利用葡萄糖、木糖及混合糖生产油脂,质量分数最高可达82.74%,比R. toruloides和L.starkeyi具有更好的产油能力,同时发现葡萄糖有助于菌株生长,木糖则更利于油脂积累。R.toruloides以葡萄糖和木糖为基质产油性能相似,均在初始糖浓度为0.40mol/L 时油脂得率最大,分别为21.62%和11.44%。与单糖相比,混合糖更适合R.toruloides生产油脂,在初始糖浓度为2mol/L 时油脂得率达到25.06%。L.starkeyi利用木糖产油效率最高,油脂得率为19.67%,而混合糖则更利于L.starkeyi菌体生长。
Ledesma−Amaro 等[52]利用“开源节流”的策略,将木糖代谢与脂质合成基因组合调控,在解脂椰氏酵母菌(Yarrowia lipolytica)中过表达外源XDH、XR 和内源XK 促进木糖代谢,过表达GPD1基因(编码甘油三磷酸脱氢酶,参与甘油三酯前体甘油−3−磷酸的形成)和DGA2 基因(编码酰基转移酶,促进脂质合成),敲除POX1-6基因(编码脂酰辅酶A 氧化酶,参与甘油三酯合成的最后一步)阻止过氧化物酶体中的β−氧化,敲除TGL4基因(编码三酰基甘油脂酶)阻断脂肪酸从脂质体中释放,避免脂质降解或利用,进一步提高了木糖利用效率与油脂产量。最终得到的工程菌株能够利用木糖唯一碳源积累高达细胞干重42%的油脂,是野生型菌株的3.40 倍。该菌株在5L 生物反应器中共利用375g/L 木糖和葡萄糖混合碳源,补料分批发酵生产22.50g/L 油脂,产率为0.06g/g(油脂/糖)。同时,产生67.20g/L 的柠檬酸,产率为0.18g/g(柠檬酸/糖)。此外,该研究还发现木糖与葡萄糖共利用更利于柠檬酸生产,而木糖与甘油共利用有助于提升油脂产量。
孔祥莉等[88]发现L.starkeyi在一定的混合糖比例下,具有同时转化木糖和葡萄糖积累油脂的能力,以木糖−葡萄糖混合液(质量比1∶2)为碳源时,油脂得率超过50%。此外,Hu等[5]研究皮丝孢酵母AS 2.571(T.cutaneumAS 2.571)时,发现其可以同时代谢葡萄糖和木糖高效生产油脂,在3L搅拌槽生物反应器中,以木糖和葡萄糖混合糖(1∶2)发酵时,油脂含量高达59%,转化率为0.17g/g(油脂/糖),且没有二次生长现象和滞后期,在不同混糖比例的摇瓶培养中可以同时完全利用这两种糖。进一步研究发现,葡萄糖为唯一碳源时,细胞油脂含量、总油脂产量和转化率分别为52.40%、12g/L和0.20g/g;然而,木糖为唯一碳源时,则分别降低至46.50%、9.90g/L和0.16g/g;葡萄糖−木糖混合糖时,细胞油脂产量略低于葡萄糖,但高于木糖。并且增加底物中木糖的占比,会略微降低油脂的生成,相比而言,使用葡萄糖生产油脂明显比单独利用木糖或葡萄糖−木糖混合物作为碳源的转化率更高。此外,T.cutaneumAS2.571 以玉米秸秆水解液为原料产油脂,质量分数达39.2%,也展现了其有较强的工业应用价值,该研究为混合糖碳源发酵产油脂的碳源配比及更高效的木糖−葡萄糖共利用提供了参考。
为了克服木质纤维素水解产生的抑制物的影响,Huang 等[54]将经稀硫酸水解后的玉米芯水解物经碱脱毒和活性炭吸附,发现L.starkeyi可以同时利用水解液中的木糖和葡萄糖生产8.10g/L的油脂。Sitepu 等[89]则利用合成水解液解决抑制问题,研究了酵母利用合成水解液(SynH)APEXTM(利用高温、高压和氨工艺预处理的技术)的发酵性能,发现APEXTM可促进部分纤维素脱结晶和半纤维素解聚,降低木质素抗逆性,增强菌株利用玉米秸秆水解液(ACSH)的产油能力。大多数酵母菌株能够在SynH 中积累油脂,只有少数能够在ACSH 培养基中生长和生产油脂,土生隐球菌(Cryptococcus humicola) 在ACSH 中生长时,能够产生高达15.50g/L 的油脂,油脂含量达到细胞干重的40%,是目前报道的最高的利用真实水解液产油脂的酵母之一。此外,还发现在SynH 培养基中以木糖作为唯一碳源预培养,可使酵母菌在随后的水解培养基中更高效地利用葡萄糖和木糖。
3.3 γ-聚谷氨酸
γ−聚谷氨酸(γ−PGA)是微生物发酵产生的水溶性多聚氨基酸,由D−谷氨酸和L−谷氨酸单体以γ-位上的酰胺键聚合而成,具有良好的吸附性和生物降解性,在生物修复及医药领域应用较多[31]。谷氨酸的两个羧基导致γ−PGA 的化学合成非常复杂,因此其制备主要依靠细菌发酵。用于生产γ−PGA 的细菌主要为芽孢杆菌,如B.subtilis、地衣芽孢杆菌(Bacillus licheniformis)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens)和纳豆芽孢杆菌(Bacillus natto)、暹罗芽孢杆菌(Bacillus siamensis)、炭疽芽孢杆菌(Bacillus anthracis)等[90]。
发酵底物成本过高是制约γ−PGA 规模化生产的重要因素。研究表明,采用合成生物学策略,引入低能耗代谢途径或合成基因[91−93]是增强底物利用效率的常用方法。例如,Feng 等[91]在B.amyloliquefaciensNK−1 中引入节能型NADPH 依赖型谷氨酸脱氢酶(NADPH−GDH)途径后,菌株NK−1 以蔗糖为底物生产γ−PGA 的产量增加了9.10%。廉价的木质纤维是更为经济的γ−PGA生产原料[94],Halmschlag 等[31]改造B.subtilis的木糖XI 途径,引入更为高效的异源木糖WBG 途径,选择木糖诱导和葡萄糖抑制的PX43 启动子元件组合,使得细胞生长和γ−PGA 的产生解耦,有利于在生长阶段从葡萄糖获得更高的生物催化剂供应,并在葡萄糖耗尽后诱导γ−PGA 合成酶的表达,构建了能高效利用木糖的工程菌。在37℃、0.50L 搅拌槽生物反应器中,分批发酵γ−PGA 产量较野生型菌株提高了6倍。借助Cobra Toolbox 和Matlab 对菌株的代谢通量分析优化后,以D−木糖和D−葡萄糖混合物(木糖占80%)为底物,γ−PGA 产量为5g/L,C摩尔转化率为0.26(γ−PGA中碳/糖中碳)。
为了验证工程菌株的木质纤维素底物的利用能力,Fang 等[55]利用B.amyloliquefaciensJX−6 以玉米秸秆和豆粕为固体基质生产γ−PGA,将产量从10kg 提高到37.50kg,在50L 和150L 曝气式连续搅拌固态生物反应器中γ−PGA 产量最高,分别为116.88g/kg 和102.48g/kg。但是,随着发酵体系扩大,γ−PGA 产量下降,可能是反应器容量增大导致发酵参数和细菌群落的变化,影响了底物利用和产量。为了进一步简化生产工艺流程,该课题组深入研究工程菌株的环境适应能力,发现B.amyloliquefaciensJX−6 对底物和发酵条件具有良好的适应性,在非灭菌条件也可实现γ−PGA 的高效生产。
3.4 有机酸
3.4.1 富马酸
富马酸(FA)是由丁烯衍生的羧酸,作为重要的聚合原料,应用于食品、医疗、饲料及化学合成中间体领域[6,44,56]。生物转化法因原料储量丰富、可再生的优点,成为工业生产FA 的更好选择[57,95−96]。生物发酵制备FA常用的原料是葡萄糖和淀粉类,以木质纤维素原料为底物生产FA 可以实现废物利用,进一步降低原料成本[6,44,96]。但是,以木糖为底物生产FA相比葡萄糖有待进一步提高。Kautola 等[97]利用R.arrhizus以木糖为碳源发酵仅得到16.40g/L的FA。Wen等[56]采用定向进化获得一株优良的菌株R.arrhizusRH7−13,不同于之前的菌株通过呼吸作用利用木糖转化为二氧化碳提高生物量和能量供应的方式,该菌株突变后,更倾向于利用木糖促进细胞生长的方式来增强FA 生产,可将发酵木糖制备FA 的产量提高至28.48g/L,木糖消耗量达到83%,转化率达到46%(FA/木糖)。为了进一步提高FA 产量,该课题组[44]利用选择培养基从R.arrhizusRH7−13 筛选得到R.arrhizusRH7−13−9#,能更高效地利用木糖生产45.31g/L FA,转化率为73%。
木糖−葡萄糖共利用发酵比单糖发酵更利于纤维素原料的充分利用。但是,当前的研究多局限于葡萄糖或木糖单独发酵生产FA,而对木糖−葡萄糖混糖共利用制备FA 的研究相对较少。为了减少发酵批次,实现木质纤维素水解物中木糖及葡萄糖混合物的同步利用,该课题组[6]以高浓度木糖分离筛选获得的新菌株R.arrhizusRH7−13−9#,并研究了其混糖协同发酵过程,利用80g/L 葡萄糖发酵得到37.52g/L FA。进一步优化后,发现混合糖利用过程中葡萄糖仍然是首选糖,但葡萄糖的添加不仅加速了FA 的生成,还增加了副产物乙醇的产量;而木糖的加入不利于副产物乙醇的生产。因此,确定了最佳木糖/葡萄糖质量比为75/25,碳氮质量比为800/1,FA 产量可达46.78g/L。探究了混糖比例与FA 产量的关系,为高效利用纤维素原料规模化生产FA提供了参考。
3.4.2 乳酸
乳酸是广泛应用于食品、医药、纺织印染等领域的多用途有机酸[58−59,98]。米根霉(Rhizopus oryzae)营养要求低、产物纯度高,使其成为最有前途的乳酸生产菌种之一。潘丽军等[58]经N+注入诱变筛选得到菌株R.oryzaeN50−7。随后采用响应面法优化培养条件,确定最佳了混合糖比例为2∶1(木糖∶葡萄糖),摇瓶发酵72h,L−乳酸产量为119.22g/L,转化率为79.48%。该实验表明利用响应面法优化培养混合糖能够协调木糖−葡萄糖代谢平衡,解除胞内代谢扰动,从而促进产物的转化。
木质纤维素水解液直接用于发酵生产L−乳酸,能使成本更低,利于产业化。姜珊[59]以玉米芯残渣为 底 物, 利 用 凝 结 芽 孢 杆 菌H−1 (Bacillus coagulansH−1)补料分批发酵,L−乳酸产量达68g/L,转化率为0.85g/g(L−乳酸/玉米芯),实现了木质纤维素向可用化学品乳酸的转化。成分复杂的水解液直接用于生产发酵时,其中的弱酸、酚类等多种物质会抑制菌株生长和产物得率。针对该问题,Zhang 等[60]分离得到一株嗜热菌株B.coagulansIPE22,能够发酵戊糖、己糖和纤维二糖,且对水解液中多种抑制物具有耐受性,结合这一特点,建立了以100g小麦秸秆为底物的高温发酵集成工艺:小麦秸秆经稀酸预处理后,无需固液分离和脱毒步骤,直接同步糖化共发酵生产46.12g乳酸,减少了设备投入,降低了能耗和生产成本。此外,该课题组[61]首次报道了利用B.coagulans发酵非食用淀粉一步法生产乳酸,利用B.coagulansIPE22 以木薯和高粱粉等非食用淀粉为原料直接分批发酵生产乳酸时淀粉酶活性过低,补加中温α−淀粉酶和糖化酶后,不灭菌,同步液化与糖化,发酵乳酸产量为68.72g/L,转化率0.99g/g。
3.4.3 其他
除上述应用外,Chen等[46]在C.glutamicum中将葡萄糖代谢途径与甘油合成途径相结合,利用肌醇渗透酶(iolT1)和葡萄糖激酶(glk)取代PEP 依赖的PTS减少副产物生成,实现木糖和葡萄糖共利用高效生产3−HP,产量达38.60g/L,转化率为0.46g/g(3−HP/葡萄糖)。在此基础上,进一步整合戊糖转运蛋白基因araE及木糖分解代谢基因XylA、XylB,实现了更高效的木糖−葡萄糖共利用。补料分批发酵,3−HP 产量提升至62.60g/L,转化率为0.51g/g,表明模块化构建功能性代谢路径有利于平衡工程菌株生长和产物合成过程。
Fu 等[62]在Clostridium tyrobutyricum(C.tyrobu⁃tyricum)中过表达来自丙酮丁醇梭菌(Clostridiumacetobutylicum)的XylT、XylA和XylB基因,获得的工程菌株Ct−pTBA 可以实现木糖−葡萄糖共利用,丁酸产量由19.40g/L 提升至37.80g/L,木糖利用率由13.80%提高至94.30%,丁酸转化率达到0.32g/g(丁酸/木糖−葡萄糖混合物)。为了便于实际生产,该课题组研究了工程菌株Ct−pTBA 共利用大豆壳、玉米纤维、小麦秸秆、水稻秸秆和甘蔗渣水解物中的木糖和葡萄糖的过程,发现Ct−pTBA较野生型菌株表现出更高的木糖利用率,且没有显著的CCR,在pH为6.0的生物反应器中,水解物中的木糖和葡萄糖利用率接近100%,丁酸产量为42.60g/L,转化率0.36g/g(丁酸/水解物),证明了Ct−pTBA 具有利用木质纤维素水解物生产丁酸的潜力。
上述木糖−葡萄糖共利用产品的应用实例展现了以木质纤维素生物质为底物生产高值化学品的巨大潜力,而木糖−葡萄糖共利用工程菌能否构建成功、混糖利用效率及产物产量除了代谢途径的糖转运蛋白效率、合成基因表达水平等关键因素外,还受底物预处理方法及分支代谢途径制约[20−22,99−103]。
4 木糖及葡萄糖共利用发酵优化策略
4.1 木糖及葡萄糖共利用基因水平调控
4.1.1 木糖及葡萄糖转运蛋白基因优化
木糖、葡萄糖被转运到胞内是被微生物代谢利用的前提,糖转运蛋白在微生物摄取糖的过程中扮演着重要角色,决定了代谢通量和糖利用效率。微生物单糖转运蛋白可分为三磷酸腺苷结合盒转运蛋白(ATP−binding cassette, ABC)、易化超家族转运蛋白(major facilitator superfamily, MFS)和磷酸烯醇式丙酮酸−糖磷酸转移酶转运蛋白(carbohydrate phosphotransferase system, PTS)三大类[104],见图3。其中MFS 型转运蛋白分为借助糖浓度梯度、不消耗能量的单向异化扩散型(facilitator/uniporter)被动运输和偶联质子或钠离子浓度消耗ATP 能量、不依赖糖浓度梯度的质子或钠离子同向协同型(H+−linked symporter,Na+−linked symporter) 主 动运输[104−105]。相关研究证明,S.cerevisiae内源非特异性戊糖转运蛋白属于不消耗能量的单向异化扩散型[9,105]。ABC 转运蛋白因其较高的底物亲和力和清晰的结构研究而成为细菌戊糖转运基因改造的优先选择。
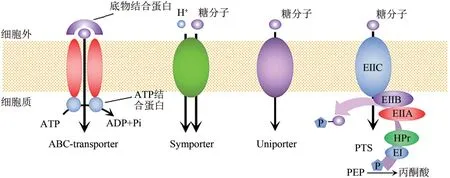
图3 转运蛋白分类示意图
木糖−葡萄糖共利用菌株构建面临的首要问题是葡萄糖的存在对木糖的分解代谢物阻遏作用(CCR),CCR突出表现在一些非特异性的木糖转运蛋白层面[106]。如S.cerevisiae的内源性己糖转运蛋白Hxt7p 和Gal2p[105,107−108],虽可以非特异性地转运木糖,但对木糖的亲和力远低于葡萄糖,且易受到葡萄糖的强烈抑制。在木糖−葡萄糖共利用过程中,利用专一性强的木糖转运蛋白增强转运蛋白对木糖的亲和力,使木糖及葡萄糖转运蛋白“各司其职”是缓解CCR 的有效策略[7,9,108−110]。例如,Shin 等[111]在缺乏Hxt1−7和GAL2基因的菌株中表达S.cerevisiae的隐性糖转运蛋白Hxt11,实现了有氧和无氧条件下木糖及葡萄糖共利用;Hxt11的诱变产生的转运蛋白与D−葡萄糖相比对D−木糖有更好的亲和力,并保持高转运率。而此前没有发现Hxt11 与D−木糖转运相关,表明内源性转运蛋白筛选优化是解决CCR的可行方法。
4.1.2 敲除抑制基因及分支代谢基因
敲除抑制基因或阻断分支代谢途径是提高产物产量和目标代谢途径碳通量的重要策略。敲除关键抑制基因可以有效调控木糖−葡萄糖共利用。蔡艳青等[112]敲除了S.cerevisiae中CCR 系统的两个关键调控因子Mig1 和Snf1,激活了氮分解代谢阻遏基因表达,增强了S.cerevisiae的混糖利用能力,使木糖消耗速率提高到初始菌株的1.23倍。Gao等[113]敲除了葡萄糖代谢途径中间产物的分支代谢基因fadR、lysC、aspB、pckA以及下游的谷氨酸分解代谢基因proAB、rocG和gudB,获得的菌株NK−A6的γ−PGA产量为4.84g/L,比初始菌株提高了31.50%。此外,启动子的类型会影响基因表达水平,且限速步骤的酶催化反应大多需要能量及辅因子参与[83]。因此,启动子的选择、胞内还原力水平调控对于增强代谢流、提高产量至关重要。该课题组进一步在异柠檬酸脱氢酶基因icd上游插入强启动子PC2up,调控NADPH 相关基因pgi和gndA表达,增加胞内还原力水平,最终γ−PGA产量提高至7.53g/L。
4.1.3 过表达产物合成基因
合成基因编码的关键酶的活性决定了反应速率,是整个代谢网络的限速步骤。过表达代谢途径合成基因是提高代谢通量、克服CCR、提高混糖共利用效率的有效方法。如姜珊[59]通过表达异源葡聚糖内切酶基因Bscel5,使B.licheniformisBN11 可以利用无定形纤维素生产D−乳酸。Fu 等[7]表达异源木糖分解代谢基因XylT、XylA和XylB,消除了C.tyrobutylicum中的CCR。构建的工程菌株可以共利用木质纤维素水解物中的木糖和葡萄糖,利用率接近100%。此外,张佳[50]过表达粗糙脉孢霉(Neurospora crassa)的Xyl1 基因、S.stipitis的Xyl2基因和白色假丝酵母(Candida albicans)的Xyl2基因,改善了辅酶不平衡问题,提高了利用木糖生产乙醇的能力。同时,敲除3−磷酸甘油脱氢酶基因GPD1降低甘油和乙酸等副产物积累,42℃条件下,实现103.97g/L 木糖和40.96g/L 葡萄糖共利用乙醇产量达到51.43g/L。
4.2 木糖及葡萄糖共利用发酵技术优化
4.2.1 细胞固定化
根据培养基的形态不同,发酵方式可分为固态发酵和液态发酵。细胞固定化发酵是指用物理或化学的方法将细胞固定于一定的结构域内,将整个固定化体系放置于液态培养基中发酵,可以结合两种方式产物密度高、可控性强的优势,具有抗逆性强、稳定性好、可重复利用等优点[114−116]。根据固定化原理和细胞表面特性不同,常用的固定化方法有包埋法、吸附法、共价法和交联法等。常见的天然有机高分子载体材料有海藻酸钠、果胶和壳聚糖等,因无毒性、传质性能优和生物相容性好,已被广泛应用于生物发酵系统[116]。
此外,根据包埋物的不同,固定化又分为单种细胞固定化、细胞−酶固定化、细胞−细胞固定化、酶固定化等形式。Xu 等[117]将海藻酸钠用于固定热纤维梭菌(Clostridium thermocellum)和热解糖梭菌(Clostridium thermolacticum)的共培养系统,与游离细胞相比,固定化细胞发酵木质纤维素底物的乙醇产量提高了60%以上。纤维素原料预处理过程中产生的有毒物质及极端pH 条件会抑制细胞生长和代谢,降低乙醇产量。海藻酸钠可包埋细胞形成微球,将菌体包埋于微球核心中,以空间隔离的方式使细胞免受发酵体系中有毒物质的伤害,同时可提高细胞对pH 变化的适应能力。此外,Evrim günes 等[118]将葡萄糖苷酶和Z.Mobilis共固定化,利用淀粉生产乙醇,发现98%的底物转化为乙醇。多种细胞共固定化利用微生物种间协调共生的特点,细胞与酶共固定化利用了微生物与酶的协同作用。
4.2.2 混菌发酵技术
混菌发酵是指利用两种或两种以上的微生物混合共培养的发酵技术,属于微生物生态工程范畴。混菌培养包括直接共培养、间接共培养和共固定化混菌培养等[119]。混菌发酵能够利用共培养微生物种间的协同代谢、诱导作用、基因转移及种间群体感应等[119−125],使发酵系统持续稳定地运行。
混菌发酵能够结合不同菌株的优势,利用种间相互作用克服一些微生物不能利用木糖或葡萄糖的限制。江枫等[121]采用包埋法固定化C.shehatae和S.cerevisiae混菌发酵(菌体比例4∶1)生产乙醇,可以快速利用葡萄糖,解除葡萄糖对木糖代谢的抑制,缩短发酵时间,产量达14.89g/L,较单菌发酵提高了12.12%。Ferreira 等[122]利用Z.mobilis和P.tannophilus混菌发酵,将Z.mobilis先于P.tannophilus接种到经稀硫酸处理后的香蕉皮水解产物中,有利于加速葡萄糖利用,解除CCR。针对S.oneidensis不能利用木糖及葡萄糖作为碳源的局限,Li 等[123−124]构 建 了K.pneumoniae和S.oneidensisMR−1的混菌产电系统,借助K.pneumoniae以葡萄糖、木糖或甘油作为底物高产乳酸,生成的乳酸作为产电菌S.oneidensisMR−1生长的碳源,形成了种间协同作用;Lin 等[125]将S.cerevisiae和S.oneidensisMR−1 混合共培养,提高了S.cerevisiae以葡萄糖为底物生产乳酸的能力,产生的乳酸作为碳源供给S.oneidensisMR−1产电。此外,混合培养还增强了S.oneidensisMR−1细胞膜通透性和导电性,使得产电功率得到较大提高。Valdez−Vazquez 等[119]在直接共培养中,将木质纤维素发酵转化为氢气,水解菌(Clostridium cellulovorans) 和发酵菌(Clostridium acetobutylicum)的接种比例为5∶3 时,建立了协同关系,利用小麦秸秆产氢气量达128mL/L,比单培养提高了2~3倍。然而,这种协同关系并非广泛适用于多种发酵底物,如:在天然麦草微生物群落的生物强化过程中,两种菌之间的协同关系消失,对甘蔗渣的发酵产氢率也没有提高。
针对混菌发酵导致副产物较多的问题,Zhang等[126]利用CCR 较弱的短乳杆菌(Lactobacillus brevis)同时利用葡萄糖和木糖生产乳酸,但乳酸转化率仅为0.52g/g,还伴随着副产物乙酸和乙醇的产生。但将Lactobacillus brevis(L.brevis)与Lactobacillus plantarum(L.plantarum) 共 培 养 后减少了副产物乙醇,显著提高了乳酸产量,L.plantarum在葡萄糖消耗上优于L.brevis,L.brevis主要将木糖转化为乳酸,由于发酵系统中产生的NADH 较少,导致副产物乙醇减少。该体系L.brevis和L.plantarum连续共发酵杨树水解液生产乳酸的转化率高达0.80g/g,碱处理玉米秸秆的乳酸转化率提高到0.78g/g,可见合理组合利用微生物之间的代谢关系,不仅可以避免多种微生物共同代谢产生较多的副产物,还能进一步提高产量。
将功能性代谢途径整合到不同的宿主菌中,利用混菌技术共利用木糖和葡萄糖可以避开复杂的微生物代谢调控过程。Saini等[127]在梭状芽孢菌辅酶A依赖的正丁醇合成途径中,敲除adhE、frdA、ldhA、poxB和pta基因以保存NADH和减少碳浪费,将CCR 不敏感的菌株转化为葡萄糖和木糖利用菌株,其中不含XylA基因的菌株BuT−8−Glu 选择性代谢葡萄糖,而缺乏glk基因的菌株BuT−8−Xyl 选择性利用木糖,将正丁醇合成途径分配到两个菌株中,建立了由两种菌组成的共培养体系,两种糖(木糖/葡萄糖质量比2∶1)共利用发酵获得5.20g/L正丁醇。Wen等[128]敲除Clostridium celllovorans(C.celllovorans)的醋酸激酶基因Clocel-1892 和乳酸脱氢酶基因Clocel-1533,过表达丁酸激酶基因Clocel-3674,将碳流量拉向丁酸生成。与此同时,为了提高乙醇产量,使用CRISPR 干扰下调了氢化酶基因Clocel-2243 的表达, 并在Clostridium beijerinckii(C.beijerinckii)中导入了有机酸再同化基因(ctfAB,cbei)和戊糖利用基因XylR和XylT。最终,改造后的C.celllovorans和C.beijerinckii组成的双梭状芽孢菌系统可分解83.20g/L经碱处理的玉米芯生产11.50g/L 丁醇,同时产生4.25g/L 丙酮和6.37g/L 乙醇,为发酵木质纤维素提供了有效的平台。
此外,混菌技术有助于减弱水解物的毒性。Theiri 等[129]以预处理后的半纤维为底物将Ureibacillus thermosphaericus和Cupriavidus taiwanensis共培养解毒,再与Paenibacillus campinasensis共培养水解,发现絮凝后酚类化合物被还原。但这种解毒作用相对有限,还需采取针对性的措施才能提高脱毒效果。
4.2.3 发酵液副产物脱毒处理技术
底物预处理过程中一般会产生一些副产物,导致发酵液成分、pH 等关键条件改变,降低产量或增加产物分离纯化难度,如酚类、糠醛、呋喃类衍生物、弱酸、碱或固形物等。研究表明,在甘蔗水解过程中会释放乙酸和糠醛,木聚糖主链上的乙酰侧链水解时,容易释放乙酸[130]。这些副产物对微生物发酵具有较强的抑制作用,乙酸会使发酵液酸性增强而影响微生物细胞的生理状态,糠醛及高浓度的呋喃类衍生物会抑制菌株生长、干扰大分子合成和糖酵解酶等[16],从而导致微生物细胞活力下降,使产量降低。
产物合成过程产生的高浓度副产物也是生物发酵规模化应用的一个重要制约。如发酵过程产生的弱酸会使发酵液的pH 降低,而混糖发酵时,木糖代谢比葡萄糖代谢对培养液酸度更敏感,低pH 更容易抑制木糖发酵[103]。Wu 等[103]在研究C.acetobutylicum的木糖−葡萄糖共利用过程时,发现当初始总糖浓度为120g/L 时,两种糖的利用率均随其浓度或比例的增加而增加,在pH 为6.0 时,总糖利用率基本不变,降低pH至5.0显著降低了糖的利用率和丁酸产量,对木糖代谢比葡萄糖的影响更明显。添加紫精促进乙酸的再吸收,丁酸产量则提高至46.40g/L、转化率为0.43g/g。张影等[131]发现混糖发酵时,葡萄糖的代谢产物乙酸、乙醇与甲酸,三者共存时对木糖发酵表现出强烈的协同抑制,而单独存在时抑制作用大大降低。因此,采取适当的底物预处理及水解液脱毒方法[22]、基因工程改造或发酵工艺改进措施等对解除产物和代谢中间体的抑制作用是必要的。
4.2.4 补料方式
根据料液添加时间和方式的不同,发酵方式通常分为分批发酵(BF)、连续发酵(CF)、补料分批发酵(FBF)等。FBF 因底物利用率及转化率高、便于自动化控制、能解除产物反馈抑制和分解代谢物阻遏作用等优势而得到广泛应用[63,132−133]。
以木质纤维素水解液为底物混糖发酵面临的最大的挑战是CCR,FBF 是解除CCR 的有效方式。Jin 等[132]在水稻秸秆补料分批半同步糖化发酵中,将酶补料(E 补料)、底物补料(SR 补料)、酶和底物组合补料(E−SR 补料)三种方式与BF 比较。结果表明:SR补料和E−SR补料乙醇产量较高,分别获得116.80g/L 和118.90g/L 的乙醇,该方法有效提高了混糖利用率。Abdel−Rahman 等[63]利用木质纤维素来源的糖借助FBF工艺得到129g/L乳酸,发现木糖浓度大于10g/L 时,可实现均质乳酸发酵、减少副产物生成,更关键的是葡萄糖浓度低于25g/L可避免CCR。经基因工程改造、系统重构代谢路径获得的工程菌采用FBF的方式生产时,能发挥二者优势,最大程度地削弱CCR[88]。此外,结合两段式发酵策略利用稀酸预处理后的玉米芯浆液生产木糖醇和乙醇的过程同样消除了CCR,第一阶段木糖在微曝气条件下生产木糖醇,第二阶段同步糖化发酵葡萄糖生产乙醇[134]。
5 结语
目前,以木质纤维素水解液中的木糖及葡萄糖共利用发酵生产醇类和有机酸等化学品已经取得许多突破性进展。但是,现阶段的研究存在一些亟待改进的问题,例如:①菌株诱变或定向进化筛选效率较低;②能够同时高效利用木糖和葡萄糖共发酵的菌株较少,分支代谢及副产物较多,基因改造及代谢调控缺乏系统性和全局性,未能完全解除CCR,代谢通量有待提高;③底物预处理方法复杂、易产生毒性物质且脱毒困难;④产物分离纯化成本较高、产量低;⑤发酵工艺适用范围窄、反应条件控制成本较高。
针对上述问题的优化方向主要有:①开发高效的、选择性好的高通量、定向菌株筛选方法;②借助合成生物学策略,利用基因工程、代谢工程、组学分析及系统工程等对菌株进行理性改造与设计,重构代谢途径,获得发酵性能优异的工程菌株;③使用物理、化学或生物等多种方法组合联用的方式设计改进底物和代谢产物的预处理及脱毒方法;④利用材料科学与生物学交叉学科优势开发性能优异的细胞固定化载体材料和固定化方法,借助菌群分析、代谢组学等科学的设计混菌生态系统;⑤针对不同宿主菌或产物分离需要个性化设计发酵工艺条件的同时兼顾工艺功能区模块化、组合化,降低工艺设计成本。
相信通过系统化、全局化、科学化的设计与针对性的工程化探索,未来储量丰富的木质纤维生物质会成为更加实用的可再生资源,而不是当作废物造成环境污染和资源浪费,以其为底物必然能够给社会经济发展创造更多有价值的产品,催生更多的绿色环保产业,产生深远的经济社会效益。
