稻虾共作模式下喷施乙烯利对水稻抗倒伏能力及产量的影响
2023-02-24


摘要:以南粳5718为供试水稻材料,在水稻拔节1叶龄期叶面喷施浓度为200 mL/hm2(ET1处理)、400 mL/hm2(ET2处理)、600 mL/hm2(ET3处理)的乙烯利,同时以喷施等量清水为对照处理(CK),研究稻虾共作模式下乙烯利施用对水稻产量形成及抗倒伏特性的影响。结果表明,与CK处理相比,ET1处理的水稻产量几乎不变,水稻株高显著下降了6.60%,茎壁厚度增加了10.46%,基部节间抗折力、折断弯矩和弯曲应力分别增加了23.47%、23.47%和29.63%,倒伏指数显著降低了31.28%,抗倒伏能力增强。尽管在ET2、ET3处理下水稻茎秆的倒伏指数比CK处理降低了18.25%~20.15%,但其抗折力、弯曲力矩也有所降低,且水稻减产12.13%~15.74%。综合来看,ET1处理的水稻稳产、抗倒伏效果最好。在ET1处理下,水稻倒伏特性各指标间的相关性分析结果表明,抗折力主要与折断弯矩、弯曲应力呈正相关,与株高呈负相关。综合分析可知,在稻虾共作模式下,喷施低浓度乙烯利可在不影响粮食安全的情况下提高水稻抗倒伏能力。
关键词:水稻;抗倒伏;稻虾共作;乙烯利;植物生长调节剂
中图分类号:S511.04 文献标志码:A
文章编号:1002-1302(2023)23-0108-06
倒伏是水稻生育后期普遍发生的问题,倒伏极大地影响了水稻产量和稻米品质,同时也降低了水稻收获的机械化率[1-2]。影响水稻倒伏的因素众多,包括遗传因素、栽培因素、气象因素及生物因素[3]。倒伏现象极为复杂,根据水稻茎秆倒伏形态可分为弯曲型、挫折型和扭转型倒伏[4]。稻虾[克氏原螯虾(Procambarus clarkii),俗称小龙虾]共作是我国最主要的稻田综合种养模式之一[5]。在这种模式下,由于小龙虾的活动(如在根土界面打洞)及較长时间的淹水灌溉等,会造成水稻茎秆折断及根部倒伏,倒伏风险加大;而稻虾田较高的土壤氮含量也可能导致植株旺长[6-7],但秆壁却过于薄弱,茎秆组织不紧密,使得稻株过高,降低了抗倒伏能力。上述问题严重制约了稻田综合种养模式的可持续发展。因此,研究如何提高稻虾共作模式下水稻的抗倒伏能力,对于确保稻田综合种养可持续发展和国家粮食安全尤为重要。
施用外源植物生长调节剂会引发植物体内源激素的变化,从而控制基因调控型作物在逆境中的生长发育[8]。目前,植物生长调节剂在作物抗倒方面的应用广泛,较为常见的植物生长调节剂有多效唑、烯效唑、芸薹素和乙烯利等。其中,乙烯利是一种与乙烯有相同作用的人工化学合成的植物生长调节剂,可释放乙烯或在一定条件下诱导植物自身进行乙烯合成,具有促进果实成熟、雌雄诱导及植株矮化等作用[9]。将乙烯利作用于玉米、大麦的研究结果表明,叶面喷施适量浓度的乙烯利能够显著提高作物茎秆强度,增强作物的抗倒伏特性,并从力学特性角度揭示乙烯利的抗倒机制,减少作物倒伏,保持籽粒产量[10-11]。近年来,也有应用乙烯利缓解水稻盐胁迫及减少稻田甲烷排放的报道[12]。前人对于乙烯利调控水稻抗倒伏特性的研究主要集中于常规稻田,鲜有作用于稻田综合种养的研究。文廷刚等研究发现,在水稻始穗期喷施乙烯利,会使植株矮化,对基部节间的作用显著,同时提高了水稻茎秆的机械强度,降低了倒伏指数[13]。孙万纯等研究指出,分蘖期或扬花后7 d施用低浓度乙烯利,可有效缩短水稻基部节间且无不良影响[14]。解振兴等利用乙烯利处理再生稻的结果则表明,拔节期叶面喷施乙烯利能够促进节间伸长,对茎壁厚度无影响,倒3节间的抗折力显著增强[15]。针对稻虾共作的抗倒伏研究主要集中在水肥管理等栽培调控措施方面[16-17],而缺乏关于外源植物生长调节剂等化控措施的报道。探究拔节初期叶面喷施不同浓度乙烯利对稻虾共作模式下水稻抗倒伏特性的调控作用及其对产量构成的影响,可为降低稻虾共作模式下水稻倒伏发生风险及稳定国家粮食安全提供理论和实践支撑。
1 材料与方法
1.1 试验地点与供试材料
试验于2022年在江苏省盱眙县扬州大学稻田综合种养创新试验基地进行,试验期间的平均气温和风速分别为26.84 ℃和2.41 m/s,总降水量为174.07 mm(图1)。试验供试水稻品种为中熟中粳稻品种南粳5718,供试植物生长调节剂为40%乙烯利水剂。供试小龙虾购自江苏省盱眙现代农业产业园发展有限公司。
1.2 试验设计
于2022年5月28日播种,6月20日移栽,水稻种植密度为30.0 cm×12.4 cm。试验共设低、中、高3个乙烯利喷施浓度,即200 mL/hm2(ET1)、400 mL/hm2(ET2)、600 mL/hm2(ET3),各处理兑水量均为751 kg/hm2,以喷施等量清水的空白处理为对照(CK)。于拔节初期晴朗无风天气的08:00—10:00,采用小型电动喷雾器进行叶面喷施,喷施时各处理间用农膜遮蔽,以防止互相干扰。试验为随机区组设计,小区面积为9 m2,3次重复,共计12个试验小区。各小区施肥量一致,氮肥投入量为 210 kg/hm2,分别于移栽前1 d、分蘖期和穗期按 3 ∶4 ∶3 的比例施用尿素。各处理磷肥、钾肥分别基施150 kg/hm2 P2O5、150 kg/hm2 K2O。田间水分管理前期与常规稻田水分管理要求一致,搁田7 d后逐步提深水位至30~35 cm,小龙虾收获前缓慢降低各小区水位,便于小龙虾退回环沟内,至水稻收获前10 d将田间水全部排出。
1.3 样品采集与测定方法
1.3.1 水稻产量及其构成因素 在水稻成熟期,选择平均长势的20株水稻普查有效穗数,选取10株水稻,对每穗粒数、结实率和千粒质量进行考种分析。去除小区边行,各小区收割30穴,测量水分后按14.5%含水量折算成实际产量。
1.3.2 茎秆形态学指标 分别于拔节期(JO)、抽穗期(HD)、抽穗后30 d(HD 30)和成熟期(MA)对各处理株高进行测量。抽穗后30 d各处理分别选取3株水稻,每株择取3个有代表性的主茎,测定重心高、节间长度、穗颈高度、鲜质量、茎粗和壁厚。重心高为茎秆基部至平衡支点的距离;节间长度为单个节间距离;穗颈高度为茎基部至穗颈节的距离;鲜质量为单个主茎的质量;将主茎第2节间从中部剪开,用游标卡尺测量该节间的长轴外径(b1)、长轴内径(b2)、短轴外径(a1)和短轴内径(a2)。茎粗及壁厚的计算公式:茎粗=(b1+a1)/2;壁厚=[(b1-b2)+(a1-a2)]/4。
1.3.3 茎秆力学指标 利用日本AIKON推拉计RZ-5测定抽穗后30 d的主茎基部第2节间抗倒相关指标及倒伏指数。将第2节间、第3节间横放在8 cm的架上,用推拉计垂直向下轻压至节间折断,读数为第2节间抗折力(N)。测量折断部位至穗顶的距离及质量,并计算力学指标。相关公式参照文献[18]:
弯曲力矩=折断部位至穗顶的鲜质量×折断部位至穗顶距离;
折断弯矩=节间抗折力×L/4(L:两支点间的距离);
断面系数=π/32×(a31b1-a32b2)/a1;
弯曲应力=折断弯矩/断面系数;
倒伏指数=弯曲力矩/折断弯矩×100%。
1.3.4 茎秆化学成分指标 取抽穗后30 d植株的第2节间茎秆作为样品,将其烘至恒质量后,将茎鞘剥离,分别磨成粉样,测定非结构性碳水化合物(NSCs)含量。采用微量法提取可溶性糖、淀粉(试剂盒购自苏州科铭生物技术有限公司,货号为KT-1-Y、DF-1-Y),用酶标仪(infinitem 200pro,Tecan,Switzerland)测定后计算NSCs含量。
1.4 统计分析
用Excel 2016、SPSS 26对产量及抗倒相关指标进行多重比较检验,用R 4.2中的corrplot包对倒伏指数与抗倒伏相关指标进行相关性分析及热图绘制。
2 结果与分析
2.1 喷施乙烯利对水稻抗倒伏能力的影响
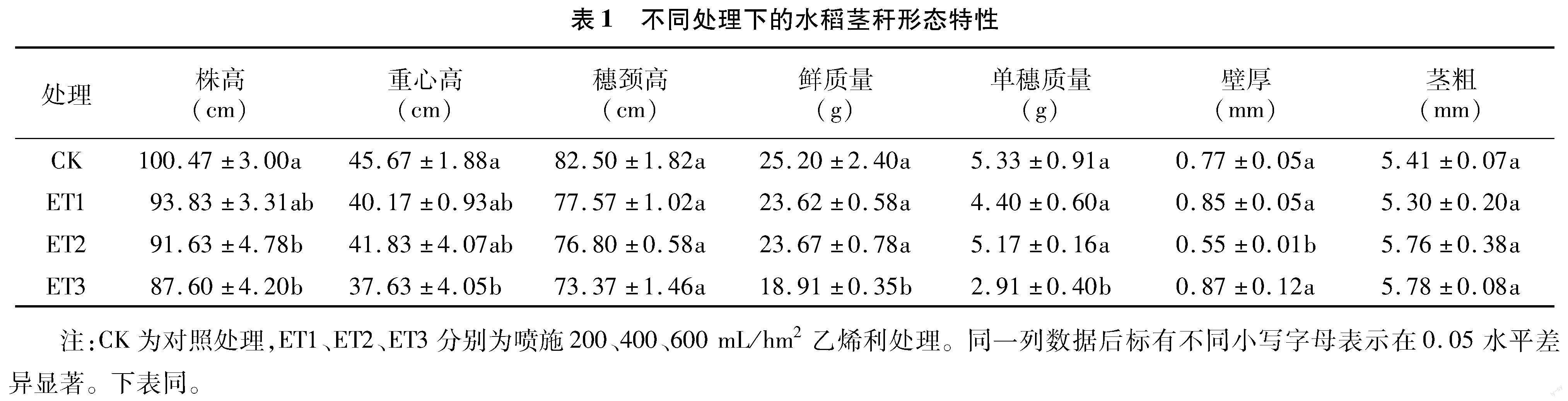
2.1.1 植株形态特征 由表1可知,喷施乙烯利降低了水稻株高,株高降低幅度随乙烯利施用浓度增加而增大。各处理关键生育期的株高表现为CK>ET1>ET2>ET3,但拔節期ET2处理的株高与CK相比差异不显著(图2-a)。喷施乙烯利在抽穗后30 d对植株重心高的影响均达显著水平(P<0.05),ET1、ET2、ET3处理分别比CK处理降低了12.04%、8.39%、17.59%。各喷施乙烯利处理的穗颈高比CK处理降低了5.98%~11.07%,但差异不显著。喷施乙烯利在降低株高和穗颈高的同时,也降低了植株鲜质量、单穗质量,各处理植株的鲜质量、单穗质量分别比CK处理降低了6.07%~24.94%、3.00%~45.40%,其中只有ET3处理的鲜质量、单穗质量差异达到了显著水平,这与ET3处理的株高、重心高和穗颈高大幅降低有关。各处理基部第2节间的长轴外径、短轴外径、长轴内径和短轴内径总体上表现为ET2>ET3>CK>ET1(图2-b)。但各处理对茎粗、壁厚的影响并不一致,与CK处理相比,ET1处理的茎粗与ET2处理的壁厚有所降低,而ET3处理的茎粗、壁厚均有增加,具体原因有待探究。此外,N1~N7节间长度总体表现为基部N1~N3节间伸长,上部N4~N7节间缩短(图2-c)。
2.1.2 茎秆力学特性及倒伏指数 由表2可以看出,喷施乙烯利后,折断部位至穗顶的鲜质量和折断部位至穗顶的距离均有不同程度的减小,减小程度均随喷施浓度的增加而增加。弯曲力矩受到这2个指标影响也呈现下降趋势,表现为ET3 2.1.3 茎秆化学成分 由表3可以看出,乙烯利对茎鞘可溶性糖、淀粉含量的影响显著,与CK处理相比,ET1、ET2、ET3处理茎秆中的可溶性糖含量分别增加了65.55%、91.14%、37.75%。叶鞘中的可溶性糖含量在CK与ET1、ET2处理间差异不显著,ET3处理则显著增加了47.34%。与CK处理相比,ET1、ET2、ET3处理茎秆、叶鞘的淀粉含量分别增加了38.53%~45.03%、 27.80%~94.88%, 差异达到显著水平(P<0.05)。 2.1.4 茎秆各倒伏指标的相关性 分析不同浓度乙烯利喷施处理对水稻产量构成及抗倒伏特性的影响发现,ET1处理的效果最佳,因此进一步探究该处理下抗倒相关指标间的相关性。由图3可以看出,茎秆抗折力与株高呈负相关,而与折断部位至穗顶鲜质量及距离、茎壁厚呈正相关,但均未达到显著水平。折断弯矩与抗折力、弯曲力矩、弯曲应力、断面系数和茎壁厚度呈正相关,其中抗折力与断面系数间的相关性达到极显著(P<0.01)、显著(P<0.05)水平。倒伏指数与株高、断面系数呈正相关,与穗颈高度、抗折力、折断弯矩、弯曲应力、可溶性糖和淀粉含量呈负相关,且除NSCs含量外均达显著水平(P<0.05)。 2.2 喷施乙烯利对水稻产量及其构成因素的影响 由表4可以看出,喷施乙烯利导致水稻产量小幅下降,不同浓度处理对产量的影响不同,其中ET2、ET3处理的水稻产量显著下降了12.13%~15.74%(P<0.05),而与CK处理相比,ET1处理的差异不显著。ET1处理与CK处理的有效穗数差异不显著,而ET2、ET3处理的有效穗数比CK处理显著减少了10.39%(P<0.05)。喷施乙烯利能够显著影响水稻的每穗粒数,各处理的每穗粒数比CK处理降低了16.13%~26.06%。ET1、ET2处理的结实率有提高趋势,而ET3处理的结实率略有降低,差异均不显著。喷施乙烯利增加了水稻千粒质量,ET1、ET2、ET3处理的千粒质量分别比CK处理显著增加了6.02%、5.06%、4.69%(P<0.05)。 3 讨论 近年来,稻渔综合种养以其高生态经济效益而在全国迅速推广,其中稻虾共作模式是长江中下游地区的主推模式,发展速度最快[19-20]。然而,由于长期淹水和土壤潜育化等问题,稻虾共作模式能否稳定粮食生产和确保粮食安全引起了农业农村部门和国土部门的担忧[21]。因此,研究稻虾共作模式下水稻稳产、抗倒伏调控措施十分必要。乙烯利对作物产量影响的相关研究发现,水稻、玉米、大麦叶面喷施或土壤施用适宜浓度乙烯利可在提高作物抗逆性的同时保持作物产量,对产量无显著负作用[12,22]。在本试验中,不同浓度乙烯利均降低了水稻产量,但喷施低浓度乙烯利处理水稻产量减幅仅为0.74%,喷施中、高浓度乙烯利处理的水稻产量减幅为12.13%~15.74%。乙烯利处理后水稻产量降低主要与有效穗数、每穗粒数降低有关,随喷施浓度升高,有效穗数呈降低趋势。 在稻虾共作过程中,土壤氮含量较高且长期淹水,更易造成水稻生长过盛、茎秆徒长而使得机械特性减弱,茎秆倒伏风险增加。Zhang等的研究表明,矮化株高、缩短节间长度是降低倒伏指数、提高茎秆强度的有效措施[23]。本研究结果表明,乙烯利处理使得水稻株高显著下降了6.60%~12.81%,穗颈高度下降了5.98%~11.07%,且降低幅度随喷施浓度增加而增大。各喷施乙烯利处理的N1~N3基部节间长度伸长,N4~N7中高部节间缩短,但整体株高显著低于CK处理。基部节间伸长可能是由于在淹水环境下,内源乙烯受到胁迫响应,促使水下基部节间伸长[24],而外源乙烯利对于植株的矮化作用主要体现为中上部节间缩短,这与文廷刚等的研究结果[13,15]一致,乙烯利主要通过降低中上部节间来降低株高及重心高。此外,乙烯利处理也影响了节间的长轴直径、短轴直径,低浓度乙烯利处理的茎壁厚比CK处理增加了10.46%,中浓度乙烯利处理的茎秆粗度增加了6.47%,而高浓度乙烯利同时增加了13.07%茎壁厚度和6.44%茎秆粗度。相关分析结果表明,茎壁厚度与节间抗折力呈正相关。喷施低浓度乙烯利有助于增加水稻茎秆节间抗折力,具体原因还有待进一步研究。茎鞘组织中NSCs含量同样被证实与茎秆强度有关[25]。本研究中,节间抗折力与NSCs含量呈正相关,而与茎秆形态指标相关性更强,说明控制植株形态可以更有效地降低水稻茎秆倒伏风险。茎秆力学特性是茎秆机械强度的重要指标。本研究还发现,喷施乙烯利降低了折断部位至穗顶鲜质量和距离、第2节间弯曲力矩和倒伏指数,且随浓度的升高而降低。其中喷施低浓度乙烯利处理的倒伏指数降低最为显著(-31.28%),这与其较低的弯曲力矩和较高的折断弯矩有关。此外可知,喷施低浓度乙烯利处理的茎秆抗倒伏能力增强与抗折力和弯曲应力的提高密切相关,这与Zhang等关于高产水稻的抗倒伏特性的结论[26]相似。乙烯利调控玉米茎秆抗倒伏特性的研究表明[27],弯曲应力和抗折力的增加可能与茎秆内纤维素含量和半纤维素含量、以及解剖结构相关,而在稻虾共作模式中的水稻抗倒伏特性调控机理有待进一步深入研究。 4 结论 稻虾共作模式下拔节初期叶面喷施低浓度乙烯利处理显著降低了水稻倒伏指数,其通过改善植株形态及力学特性,提高水稻茎秆的抗倒伏能力。与CK处理相比,叶面喷施低浓度乙烯利处理的水稻产量基本维持不变。稻虾共作模式下,喷施低浓度(200 mL/hm2)乙烯利处理可在水稻稳产的前提下提高水稻抗倒伏特性,为最佳喷施浓度。 参考文献: [1]梁玉刚,陈奕沙,陈 璐,等. 垄作稻-鱼-鸡共生对水稻茎秆倒伏、穗部性状及产量的影响[J]. 中国生态农业学报(中英文),2021,29(2):379-388. [2]Liu Q,Yin C S,Li X,et al. Lodging resistance of rice plants studied from the perspective of culm mechanical properties,carbon framework,free volume,and chemical composition[J]. Scientific Reports,2022,12(1):20026. [3]雷小龍,刘 利,苟 文,等. 种植方式对杂交籼稻植株抗倒伏特性的影响[J]. 作物学报,2013,39(10):1814-1825. [4]Ishimaru K,Togawa E,Ookawa T,et al. New target for rice lodging resistance and its effect in a typhoon[J]. Planta,2008,227(3):601-609. [5]张玉山,黄晓声,梁志辉,等. 稻虾共作模式下水稻品种和施肥方式的筛选试验[J]. 江苏农业科学,2023,51(7):66-73. [6]喻召雄,陶先法,贾 睿,等. 2种稻虾共作模式对土壤有机氮矿化作用的影响[J]. 南方农业学报,2022,53(5):1357-1367. [7]杨智景,顾海龙,顾 明,等. 稻虾种养模式对土壤肥力的影响[J]. 江苏农业科学,2020,48(23):245-249. [8]Waadt R,Seller C A,Hsu P K,et al. Plant hormone regulation of abiotic stress responses[J]. Nature Reviews Molecular Cell Biology,2022,23:680-694. [9]Kaur P,Mal D,Sheokand A,et al. Role of plant growth regulators in vegetable production:a review[J]. International Journal of Current Microbiology and Applied Sciences,2018,7(6):2177-2183. [10]Zhang Y S,Wang Y B,Ye D L,et al. Ethephon improved stalk strength of maize (Zea mays L.) mainly through altering internode morphological traits to modulate mechanical properties under field conditions[J]. Agronomy,2019,9(4):186. [11]Ma L B,Leibovitch S,Maloba W E,et al. Spring barley responses to nitrogen fertilizer and ethephon in regions with a short crop growing season[J]. Journal of Agronomy and Crop Science,2008,169(3):151-160. [12]Cho S R,Verma P P,Das S,et al. A new approach to suppress methane emissions from rice cropping systems using ethephon[J]. Science of the Total Environment,2021,804:150159. [13]文廷剛,王伟中,杨文飞,等. 水稻茎秆形态特征与抗倒伏能力对外源植物生长调节剂的响应差异[J]. 南方农业学报,2020,51(1):48-55. [14]孙万纯,吴建中,仇广灿,等. 乙烯利等生长调节剂在直播水稻上的应用效果研究[J]. 安徽农学通报(下半月刊),2011,17(4):70-72. [15]解振兴,张居念,林 祁,等. 植物生长调节剂对再生稻头季抗倒伏能力和两季产量的影响[J]. 中国水稻科学,2019,33(2):158-166.[HJ2mm] [16]徐 强,李京咏,戴林秀,等. 氮肥管理对稻虾共作模式水稻产量和抗倒伏特性的影响[J]. 中国稻米,2022,28(6):30-36. [17]Gao H,Dou Z,Cheng L R,et al. Effects of semi-deep water irrigation on hybrid indica rice lodging resistance[J]. Frontiers in Plant Science,2022,13:1038129. [18]Ookawa T,Ishihara K. Varietal difference of physical characteristics of the culm related to lodging resistance in paddy rice [Oryza [JP3]sativa][J]. Japanese Journal of Crop Science,1992,61(3):419-425. [19]Xu Q,Dai L,Shang Z Y,et al. Application of controlled-release urea to maintain rice yield and mitigate greenhouse gas emissions of rice-crayfish coculture field[J]. Agriculture,Ecosystems & Environment,2023,344:108312. [20]Wei Y B,Lu M A,Yu Q Y,et al. Understanding the dynamics of integrated rice-crawfish farming in Qianjiang county,China using Landsat time series images[J]. Agricultural Systems,2021,191:103167. [21]曹凑贵,江 洋,汪金平,等. 稻虾共作模式的“双刃性”及可持续发展策略[J]. 中国生态农业学报,2017,25(9):1245-1253. [22]Avat S,Emam Y. Plant growth regulator (ethephon) alters maize (Zea mays L.) growth,water use and grain yield under water stress[J]. Journal of Agronomy,2008,7(1):41-48. [23]Zhang X B,Wang Y,Wang X W,et al. A very-long-chain fatty acid synthesis gene,SD38,influences plant height by activating ethylene biosynthesis in rice[J]. The Plant Journal,2022,112(4):1084-1097. [24]赵 赫,陈受宜,张劲松. 乙烯信号转导与植物非生物胁迫反应调控研究进展[J]. 生物技术通报,2016,32(10):1-10. [25]Kashiwagi T,Ishimaru K. Identification and functional analysis of a locus for improvement of lodging resistance in rice[J]. Plant Physiology,2004,134:676-683. [26]Zhang J,Li G H,Song Y P,et al. Lodging resistance characteristics of high-yielding rice populations[J]. Field Crops Research,2014,161:64-74. [27]Zhao Y T,Lv Y J,Zhang S A,et al. Shortening internodes near ear:an alternative to raise maize yield[J]. Journal of Plant Growth Regulation,2022,41:628-638.
