茶树光响应曲线模型的筛选与光响应特征比较
2023-02-24刘东娜李兰英龚雪蛟黄藩尧渝胥亚琼高远罗凡
刘东娜 李兰英 龚雪蛟 黄藩 尧渝 胥亚琼 高远 罗凡


摘要:通过筛选茶树最适的光响应曲线模型,比较分析茶树光响应特征的种间差异特征,为揭示茶树的光能利用特性提供数据支持。利用Li-6800便携式光合仪测定4个品种茶树的光响应曲线及参数,运用3种光合模型和Photosynthesis系统拟合光响应曲线,并通过拟合参数的比较,筛选出茶树最适合的光响应曲线模型,比较分析茶树不同品种的光强-光合响应特征。结果表明,直角双曲线的修正模型-叶子飘模型(YEM)拟合茶树光强-光合响应曲线的确定系数(R2)、残差平方和(RSS)及均方误差(MSE)均在合理范围,拟合得出的最大净光合速率、光饱和点、光补偿点等特征参数与实测值最为接近;4个茶树品种中,绿色系茶树品种福鼎大白茶(FD)的Pnmax(10.89)、LSP(988.10)、α(0.06)和AQE(0.10)最大,LCP(27.93)最小。综合分析,叶子飘模型(YEM)是茶树叶片光响应的最佳数学模型;与黄色系茶树品种相比,绿色系茶树品种对弱光和强光的利用能力均较高,利用的光合有效辐射范围较大,生态适应能力较强。
关键词:茶树;光响应模型;光响应特性
中图分类号:S571.101 文献标志码:A
文章编号:1002-1302(2023)23-0146-06
光合作用是茶树生长发育、生理过程和物质代谢的基础,也是决定茶树生产力和鲜叶品质的重要因素之一。叶片净光合速率是反映茶树光合作用能力的重要指标,光合有效辐射(PAR)是影响叶片净光合速率最直接、最根本的环境因子[1]。茶树光响应曲线是指其叶片净光合速率(Pn)随光合有效辐射(PAR)变化而变化的基本规律,从中可获得光饱和点、光补偿点、暗呼吸速率以及光合量子效率等光合生理参数,进而了解茶树光合利用能力及光响应特征,推断茶树的光合潜力和环境适应能力[2-3]。可见,探究茶树光强-光合响应特征对评价茶树的环境适应性、选育新品种及配套栽培管理技术等具有重要意义。
近年来,有关植物光强-光合响应曲线的研究较多。陈可可等对金荞麦不同种质光响应曲线特性研究结果表明,种质JQM4的光补偿点(LCP)、光饱和点(LSP)相对较高,对强光的利用能力较强;种质JQM5的LCP较低,LSP较高,对光能利用范围较广[4]。叶英林等通过对不同叶色辣椒光响应特征研究发现,紫色辣椒具有良好的强光适应性,光合潜力较大,在选育耐强光的高产品种方面具有潜在的利用价值[5]。然而,不同植物光响应特征及适宜的光响应曲线模型等差异较大,同一植物不同光响应曲线模型拟合效果及参数结果亦有所不同[6-7]。而光响应曲线模型拟合结果中各参数的合理性会影响植株光合生理过程的判断,最优拟合模型的选择变得尤为重要。
常用的光响应数学模型有直角双曲线模型(RHM)、非直角双曲线模型(NRHM)、直角双曲线的修正模型-叶子飘模型(YEM)、指数函数模型(EFM)等[8-9]。本研究以4个茶树品种为材料,测定其光响应曲线和参数,并利用RHM、NRHM、YEM 3种数学模型及Photosynthesis系统进行拟合,筛选出茶树叶片光响应的最适光合模型;同时对4个品种茶树的光响应曲线和参数进行比较分析,以期对茶树的优质高效栽培和种质评价提供数据支持。
1 材料与方法
1.1 试验材料
供试茶树品种为福鼎大白茶、金凤1号、金凤2号和中黄1号,树龄为5年生。品种详细介绍见表1。试验区位于四川省雅安市名山区茅河鎮龙兴村(103.368 813°E,30.217 201°N)。试验时间为2019年5月24日。
1.2 试验方法
选择5月下旬晴朗天气,在09:00—11:00时测定,每个品种随机选取长势一致的新梢,取当年生新梢成熟叶(芽下第5叶)为测定叶,每个品种取3株光合数据的平均值。
采用北京力高泰科有限公司生产的Li-6800便携式光合仪Light-Curve曲线测定功能,测定各个茶树品种叶片的光响应曲线。测定前用 1 000 μmol/(m2·s) 光强对茶树叶片诱导20 min。测定时叶室温度、相对湿度、气流速度、气压和CO2浓度等分别设定为28 ℃、60%、500 μmol/s、0.1 kPa和400 μmol/mol,光合有效辐射(PAR)从0~1 500 μmol/(m2·s)分为9个梯度[1 500、1 200、900、600、300、150、100、50、0 μmol/(m2·s)]进行光合(Pn-PAR)响应曲线测定。光照度改变后,稳定 3 min,由仪器自动记录光合参数。
1.3 模型
本研究选用的茶树叶片光响应曲线拟合数学模型分别为RHM、NRHM和YEM。各模型的数学表达式见表2。表2中:Pn为净光合速率;I为光合有效辐射;Pnmax为最大净光合速率;Rd为暗呼吸效率;α为初始量子效率;θ为非直角双曲线的凸度,0≤θ≤1,凸度与曲线的弯曲程度正相关;β为修正系数;γ=α/Pnmax;AQE为表观量子效率,通过对弱光强条件下[I≤200 μmol/(m2·s)]的光响应数据进行直线(Pn=AQE×I+Rd)回归,直线方程的斜率即为AQE;该直线和直角双曲线模型、非直角双曲线模型拟合的Pnmax与X轴平行的直线(Y=Pnmax)相交,其交点在X轴上的数值为LSP[17]。各模型参数初始值设定为:直角双曲线模型α=0.05,Pnmax=30,Rd=2;非直角双曲线模型α=0.05,θ=0.5,Pnmax=30,Rd=2;叶子飘新模型α=0.01,β=0.0001,γ=0.001,Rd=0.5。
用Excel 2010和SPSS 19软件对数据进行整理和方差统计学分析,用Duncans新复极差法进行多重比较;用SPSS 19软件和Photosynthesis软件进行曲线拟合。根据拟合出的光响应曲线,得出确定系数(R2)、均方误差(mean square error,简称MSE)和平均绝对误差(mean absolute error,简称MAE)等拟合度参数,计算出最大净光合速率(Pnmax)、初始量子效率(α)、暗呼吸速率(Rd)、光饱和点(LSP)以及光补偿点(LCP)等光响应特征参数[18-20]。用Origin 2019作图。
2 结果与分析
2.1 茶树光响应曲线的拟合
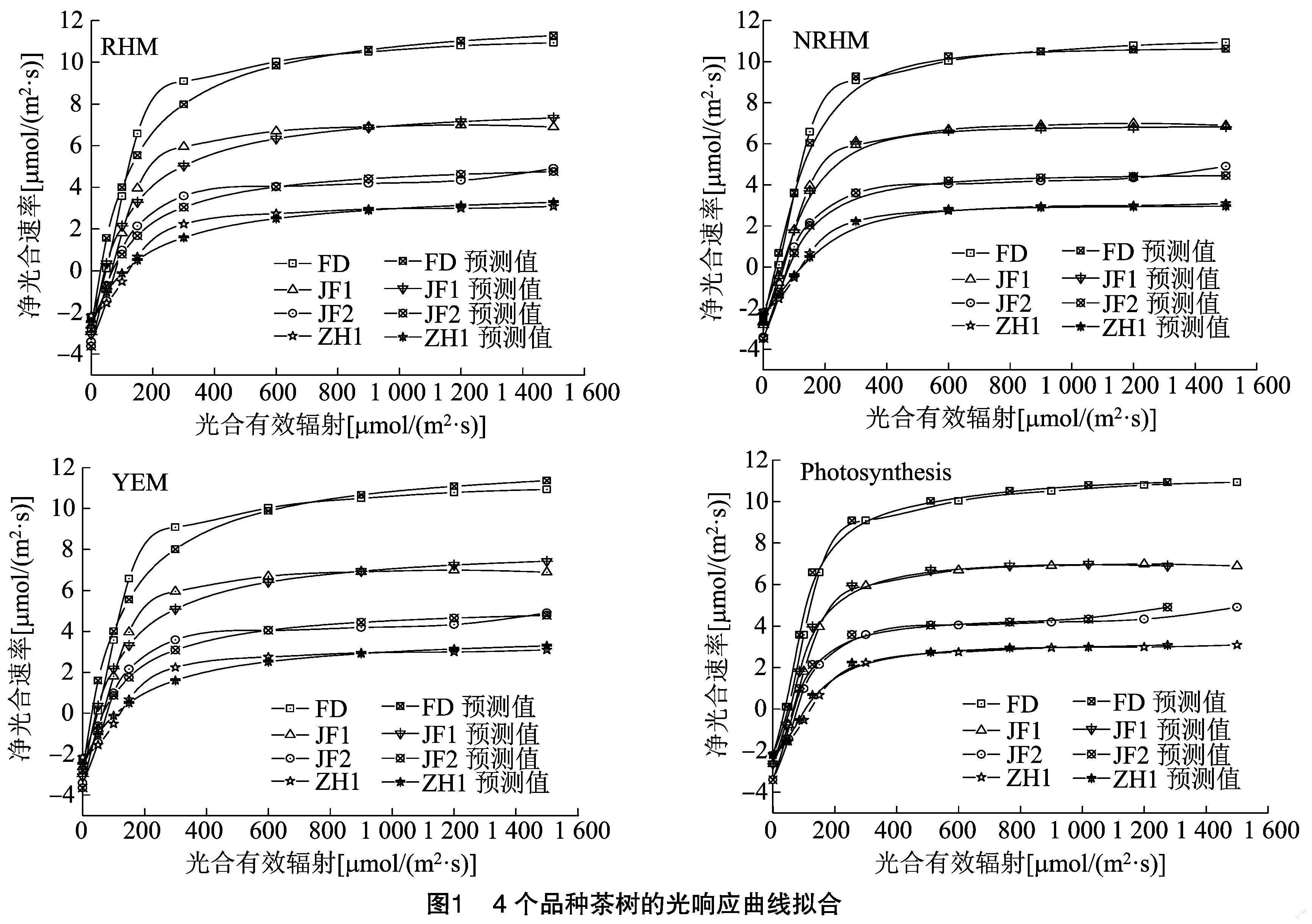
光照强度直接影响茶树的光合效率。4个品种茶树采用不同模型进行光强-光合响应曲线拟合(图1),结果显示,各个模型拟合所得的4个茶树品种的光响应曲线与实测曲线均较为接近,符合植物叶片的光响应基本特征,均具有较好的拟合效果。不同品种茶树光响应曲线有一定差异,但大体表现为随光合有效辐射(PAR)增强茶树净光合速率(Pn)迅速增大,且增幅随着PAR增强而逐渐减缓,在达到光饱和点后Pn趋于平稳。各个模型所拟合的光响应曲线变化趋势与实测值类似,当PAR为0时,各品种Pn均为负值;当PAR为0~200 μmol/(m2·s) 时,各品种茶树叶片的Pn均呈现直线上升的趋势;当PAR为200~900 μmol/(m2·s)时,各品种茶树叶片的Pn增长趋势减缓;光强超过900 μmol/(m2·s)后,各品种茶树叶片的Pn逐渐趋于稳定。PAR达到 900 μmol/(m2·s) 时,各品种茶树叶片的净光合速率差异显著(P<0.05),表现为福鼎大白茶(FD)>金凤1号(JF1)>金凤2号(JF2)>中黄1号(ZH1)。
确定系数R2的大小可判定方程拟合精度的高低,R2越大,模型的精确度越高[19]。由表3可知,3种光响应曲线模型拟合度均较优,R2均在0.969以上,其中非直角双曲线模型(NRHM)的R2最高,达0.995以上。残差平方和(RSS)和均方误差MSE的值可判断预测值与实测值之间的差异,其值越小表明模型的拟合值越接近观测值[20]。同一品种不同拟合模型的残差平方和均方误差有一定差异,但均在合理范围,其中以非直角双曲线模型(NRHM)的残差平方和(RSS)和均方误差MSE值较低。
2.2 光响应特征参数
光响应曲线参数模型拟合值与实测值见表4。不同品种茶树的最大净光合速率Pnmax测量估计值为3.06~10.87 μmol/(m2·s),在模拟过程中,通过RHM模型、NRHM模型和Photosynthesis系统拟合曲线所得的各品种茶树的Pnmax分别为6.42~15.25、5.34~13.35、4.67~13.04 μmol/(m2·s),均远大于实测值;YEM模型拟合的茶树Pnmax值与测量估计值较为接近,为3.10~10.89 μmol/(m2·s)。Pnmax在各品种间的差异表现为福鼎大白茶(FD)>金凤1号(JF1)>金凤2号(JF2)>中黄1号(ZH1)(P<0.05)。
不同品种茶树光饱和点LSP估计值为600~900 μmol/(m2·s),且以福鼎大白茶品种的光饱和点最高。不同模型拟合所得茶树叶片的光饱和点差异较大,其中RHM和NRHM模型拟合4个品种茶树光饱和点分别为304.56~468.23 μmol/(m2·s)[JP+1]和266.65~402.42 μmol/(m2·s),以中黄1号茶树品种的光饱和点最高,与估计值相差甚大。这可能与RHM、NRHM模型本身特点有关;RHM和NRHM模型是一条渐近线,不存在极值,所以无法直接估算出茶树叶片的Pnmax和LSP;而现在常用的,在低光强条件下[≤200 μmol/(m2·s)],植物叶片的净光合速率响应光强的直线方程与RHM和NRHM模型拟合的曲线相交,其与Pnmax的交点在x轴上的数值为LSP;但这种方法计算得出的茶树LSP与估计值相差较大;这种现象在结球甘蓝、胡萝卜、冬小麦等植物的拟合过程中也存在[21-23]。Photosynthesis拟合各茶树品种的光饱和点为432.00~522.00 μmol/(m2·s),以福鼎大白茶品种的光饱和点最高,与估计值有一定差异。YEM模型拟合各茶树品种的光饱和点为445.81~988.10 μmol/(m2·s),与估计值较为接近;各品种间差异表现为福鼎大白茶(FD)>金凤1号(JF1)>金凤2号(JF2)>中黄1号(ZH1)(P<0.05)。
不同品种茶树初始量子效率α各模型拟合值有一定差异,其中NRHM模型和Photosynthesis系统拟合值较为接近,为0.02~0.08;RHM和YEM模型拟合值较为接近,为0.03~0.12;各品种间差异表现为福鼎大白茶(FD)>金凤1号(JF1)≈金凤2号(JF2)>中黄1号(ZH1)(P<0.05)。不同品种茶树暗呼吸速率Rd与光补偿点LCP在各模型中的拟合值与估计值较为一致,其中Rd值在各品种间差异表现为金凤2号(JF2)>金凤1号(JF1)>福鼎大白茶(FD)>中黄1号(ZH1)(P<0.05),LCP值在各品种间差异表现为中黄1号(ZH1)>金凤2号(JF2)>金凤1号(JF1)>福鼎大白茶(FD)(P<0.05)。
3 讨论
植物光强-光合响应曲线最适模型的确定,对揭示植物光反应过程、光合效率、光适应性等光合生理生态特征具有重要的意义[24-27]。本研究利用3种数学模型和Photosynthesis系统对4个茶树品种的光强-光合响应曲线进行拟合,并通过决定系数R2、残差平方和(RSS)和均方误差(MSE)等参数判定模型的优劣和精确度,通过最大净光合速率(Pnmax)、光饱和点(LSP)等参数与实测值的比较分析判断模型的适用性,进而保证拟合所得的光响应特征参数的准确性和可比性。3个光响应曲线模型决定系数R2、残差平方和(RSS)和均方误差(MSE)均在合理范围,均可以用于茶树光响应曲线的拟合;其中以NRHM模型决定系数R2最大(≥0.99),残差平方和(RSS)和均方误差(MSE)最小。然而,RHM模型和NRHM模型是一条没有极值的渐近线,拟合得到的各品种茶树叶片Pnmax分别较估计值高22.85%~74.35%和40.33%~109.77%,且通过计算得出的茶树叶片LSP与实测值相差甚大;Photosynthesis系统拟合得到各茶树品种的最大净光合速率与光饱和点与估计值亦有一定差異。可见,采用RHM、NRHM和Photosynthesis系统拟合茶树光响应曲线所得光响应特征参数的准确性并不理想。YEM模型是一条有极点的曲线,可拟合出光合响应曲线的最大净光合速率和光饱和点等特征参数,且该模型拟合得出的茶树叶片Pnmax、LSP、LCP等特征参数与实测值较为接近,是茶树的最适光强-光合响应曲线模型。该结果与张赟齐等、刘子凡等、南吉斌等的研究结果较为相似,认为YEM模型是无患子、木薯、沙棘等植物叶片光响应曲线的最佳模型[18,20,25]。
植物光强-光合响应曲线大体有2种表现:一种是随光合有效辐射增加光合速率快速增强,达光饱和点后,光合速率趋于平稳;另一种是光合速率达光饱和点后,随光合有效辐射增加而略有下降,表现出光抑制现象[28-29]。4个茶树品种光强-光合响应曲线趋势基本一致,均有快速响应阶段和平稳阶段;其中,各品种茶树叶片的Pn值快速响应阶段的PAR为0~200 μmol/(m2·s),平稳阶段的PAR为≥900 μmol/(m2·s),PAR介于200~900 μmol/(m2·s) 时,各品种茶树叶片的Pn值缓慢增加。本试验条件下,4个茶树品种的净光合速率均未出现光抑制现象,这可能与茶树自身的生理节奏或其他影响因子有关。茶树是常绿植物,喜光耐阴,其净光合速率除受光合有效辐射变化影响外,还与叶片色泽、气温、相对湿度、蒸腾速率、水分利用效率、气孔导度及胞间CO2浓度等因素密切相关[3,30]。
茶树不同品种的光响应特征参数差异较大。其中,最大净光合速率Pnmax是反映茶树叶片光合潜力的重要指标之一。各品种的Pnmax差异表现为福鼎大白茶(FD)>金凤1号(JF1)>金凤2号(JF2)>中黄1号(ZH1)(P<0.05),这可能与各品种的叶片色泽有关,韩楠等研究表明,绿色系茶树品种福鼎大白茶的Pnmax显著高于郁金香、黄金芽、金光等黄色系茶树品种[3]。暗呼吸速率(Rd)则是指有效光辐射为0时呼吸速率的值[31],金凤1号(JF1)和福鼎大白茶品种Rd值较低,说明其耐阴性相对较强。表观量子效率(AQE)为弱光条件下净光合速率与相应光量子通量密度的比值,可反映植物叶片的光能转化效率和弱光利用能力[17]。初始量子效率(α)和光补偿点LCP可用于评价植物叶片对弱光的利用能力,光饱和点LSP可用于判断植物叶片对强光的利用能力[8,31]。4个茶树品种中,福鼎大白茶(FD)的α(0.06)、AQE(0.10)和LSP(988.10)最大,LCP(27.93)最小;说明福鼎大白茶品种利用弱光和强光的能力均较高,利用的光合有效辐射范围较大,生态适应能力较强;这可能与福鼎大白茶属于绿色系品种有关,绿色系茶树品种叶片叶绿素、类胡萝卜素等光合色素含量较黄色系品种增多,具有较强的吸收和转化光能的能力,进而使得其自身具有较强的光合能力、生长势和生态适应性等[32]。
4 结论
叶子飘模型(YEM)拟合茶树光强-光合响应曲线的确定系数R2、残差平方和(RSS)和均方误差(MSE)均在合理范围,拟合得出的茶树叶片最大净光合速率、光饱和点、光补偿点等光合特征参数与实测值较为接近,是茶树的最适光强-光合响应曲线模型。
茶树4个品种中,最大净光合速率(Pnmax)和光饱和点(LSP)的拟合值排序为福鼎大白茶(FD)>金凤1号(JF1)>金凤2号(JF2)>中黄1号(ZH1);暗呼吸速率(Rd)排序为金凤2号(JF2)>金凤1号(JF1)>福鼎大白茶(FD)>中黄1号(ZH1),初始量子效率(α)和表观量子效率(AQE)排序为福鼎大白茶(FD)>金凤1号(JF1)≈金凤2号(JF2)>中黄1号(ZH1);光补偿点(LCP)值排序为中黄1号(ZH1)>金凤2号(JF2)>金凤1号(JF1)>福鼎大白茶(FD)。与金凤1号、金凤2号和中黄1号等黄色系茶树品种相比,绿色系茶树品种福鼎大白茶(FD)的Pnmax(10.89)、LSP(988.10)、α(0.06)和AQE(0.10)最大,LCP(27.93)最小,对弱光和强光的利用能力均较高,利用的光合有效辐射范围较大,生态适应能力较强。
参考文献:
[1]盖淑杰,王奕雄,李 兰,等. 茶树生长光调控研究进展[J]. 茶叶科学,2022,42(6):753-767.
[2]沈守艮,郑 征. 西双版纳胶-茶群落中茶树的光合特性及其影响因子[J]. 应用与环境生物学报,2008,14(1):32-37.
[3]韩 楠,唐 茜,赖云松,等. 四川引进的茶树特色品种黄金芽、金光、郁金香的光合特性[J]. 西南农业学报,2015,28(4):1600-1605.
[4]陈可可,黄 宣,黄莉娟,等. 8个金荞麦种质光合及光响应曲线特性研究[J]. 耕作与栽培,2022,42(3):1-6,18.
[5]叶英林,欧立军,邹学校. 辣椒不同紫色程度叶片的光合特性及光响应模型拟合研究[J]. 园艺学报,2018,45(6):1101-1114.
[6]李仕杰,朱润军,杨 巧,等. 不同生境下木棉幼苗光响应特性及最适模型拟合[J]. 西部林业科学,2020,49(5):149-154.
[7]魏炳康,李根柱,王贺新,等. 软枣猕猴桃的光合模型筛选与光响应特征比较[J]. 科学技术与工程,2021,21(17):7034-7039.
[8]刘佳哲,余小红,樊东函,等. 广西4种珍贵树种在桉树林下间种的光合响应[J]. 中南林业科技大学学报,2023,43(1):96-103.
[9]金建新,李株丹,黄建成,等. 宁夏引黄灌区不同灌水处理下春小麦光响应曲线模型研究[J]. 中国农机化学报,2022,43(9):182-190.
[10]江昌俊. 茶树育种学[M]. 北京:中国农业出版社,2011.
[11]李兰英,堯 渝,龚雪蛟,等. 茶树叶色黄化型新品种金凤1号选育研究[J]. 安徽农业科学,2022,50(19):20-24.
[12]李兰英,胥亚琼,刘东娜,等. 茶树新品种‘金凤2号[J]. 园艺学报,2022,49(增刊2):283-284.
[13]杨亚军,陈 明,王新超,等. 特异优质茶树新品种“中黄1号”选育研究[J]. 中国茶叶,2014,36(9):8-9,17.
[14]Baly E C.The kinetics of photosynthesis[J]. Proceedings of the Royal Society of London Series B-Biological Sciences,1935,117(804):218-239.
[15]Thornley J H M. Mathematical models in plant physiology:a quantitative approach to problems in plant and crop physiology[M]. London:Academic Press,1976:86-110.
[16]Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica,2007,45(4):637-640.
[17]叶子飘,于 强. 光合作用光响应模型的比较[J]. 植物生态学报,2008,32(6):1356-1361.
[18]张赟齐,高世轮,卫星杓,等. 光合模型对无患子叶片光合响应参数计算结果的影响[J]. 北京林业大学学报,2019,41(4):32-40.
[19]蒋高明,何维明. 一种在野外自然光照条件下快速测定光合作用-光响应曲线的新方法[J]. 植物学通报,1999,34(6):712-718.
[20]刘子凡,魏云霞,黄 洁. 木薯光合-光响应曲线的模型拟合比较[J]. 云南农业大学学报(自然科学),2018,33(4):611-616.
[21]蔡 霞,张德军,郭惊涛,等. 结球甘蓝的光合模型筛选与光响应特征比较[J]. 种子,2021,40(11):117-121,136.
[22]Kyei-Boahen S,Lada R,Astatkie T,et al. Photosynthetic response of carrots to varying irradiances[J]. Photosynthetica,2003,41(2):301-305.
[23]Yu Q,Zhang Y Q,Liu Y F,et al. Simulation of the stomatal conductance of winter wheat in response to light,temperature and CO2 changes[J]. Annals of Botany,2004,93(4):435-441.
[24]薛 晴,陈 斌,杨小梅,等. 不同光强下4种鸭跖草科植物的生物量分配、水分生理及光响应特征[J]. 草业学报,2022,31(1):69-80.
[25]南吉斌,杨广环,赵玉文,等. 5种光合模型对沙棘属3种植物叶绿素荧光光响应曲线的拟合效果比较分析[J]. 西部林业科学,2019,48(2):90-96.
[26]刘启航,曹进军,付晓伟,等. 波谱光照效应对西花蓟马趋光响应变化的影响[J]. 江苏农业科学,2021,49(20):124-132.
[27]李雪飞,陈 珑,饶惠玲,等. 5种丛生竹叶片光响应曲线拟合模型比较[J]. 植物资源与环境学报,2022,31(2):88-90.
[28]劉 兴,赵 丹,杨 晓,等. 遮光对糠椴幼苗光合特性的研究[J]. 林业与生态科学,2020,35(1):86-92.
[29]杜 澜,夏 捷,李海花,等. 逐步失水过程中绿竹光响应进程及其拟合[J]. 应用生态学报,2019,30(6):2011-2020.
[30]杜旭华,马 健,彭方仁. 温室内不同茶树品种净光合速率及其生理生态因子日变化[J]. 浙江林业科技,2007,27(3):28-33.
[31]韩 晓,王海波,王孝娣,等. 不同砧木对‘87-1葡萄光合特性及荧光特性的影响[J]. 中国农业科学,2018,51(10):1972-1981.
[32]Wang L,Yue C,Cao H L,et al. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar[J]. BMC Plant Biology,2014,14(1):1-13.
