次生林栽针保阔效果及影响因子探讨
2023-02-20叶林李巍巍于添邓长贺
叶林 李巍巍 于添 邓长贺
(1.黑龙江省林业科学院伊春分院,黑龙江 伊春 153000;2.伊春职业学院,黑龙江 伊春 153000;3.国家林业和草原局重点国有林区森林资源监测中心 黑龙江 大兴安岭 165000)
研究表明,红松作为建群种在原始阔叶红松林的树种组成上占40%~90%,而经破坏后形成的次生林中红松种群数量极度缺乏,成为制约阔叶红松林生态系统恢复的瓶颈[1]。我国老一辈林学家和林业工作者在研究及实践的基础上提出了“栽针保阔”的经营思想。即人工栽植以红松为主的针叶树,保留天然更新的多种阔叶树,把阔叶树的天然更新和针叶树的人工更新密切结合起来,以形成符合于地带性特征的针阔混交林。栽针是缩短森林自然(演替)恢复过程的重要手段;保阔是迅速形成(或恢复)地带性顶极森林的可靠保证[2,3]。
然而,虽然“栽针保阔”经营思想得到广泛认可,但由于没有完整合理的应用技术规范,使该技术在应用实践过程中的效果良莠不齐。本文通过红松林冠下更新造林跟踪调查结合红松幼树光合速率测定及影响净光合速率环境因子研究,探讨次生林林冠下红松更新造林保存率的主要影响因子,旨在为完善“栽针保阔”科学途径及工艺提供参考及支撑。
1 研究区概况
伊春市位于黑龙江省东北部,地处127°37′~130°46′E,46°28′~49°26′N。属北温带大陆性季风气候,年平均气温-1~1℃,最冷为1月份,气温-20~-25℃,最热为7月份,气温20~21℃,极端最高温为35℃。全年≥10℃活动积温1800~2400℃,无霜期90~120天。年平均日照数2355~2400h。年降雨量550~670mm,多集中在夏季。干湿指数1.13~0.92,属湿润地区。伊春地貌特征为“八山半水半草一分田”,整个地势西北高、东南低,南部地势较陡,中部较缓,北部较平坦,海拔高度平均600m。境内千米以上高峰77座,最高山为平顶山,海拔1423m。伊春林区经营总面积382.4万hm2,有林地面积318.7万hm2,森林覆被率83.4%,山地土壤以暗棕壤为主。试验区位于伊春市翠峦林业局翠峦河经营所和解放经营所,2010年进行次生林林冠下红松更新造林,造林密度3300株/hm2,造林后进行了5年的幼林抚育。
2 材料与方法
观测样地面积50m×50m, 样地处于山坡下部,坡度5°~8°,3个重复样地坡向分别为南坡、西南坡和西坡。次生林树种组成3白3云1柳1冷1杨1榆,林分平均胸径10.9~11.5cm,郁闭度0.7~0.8。2010、2013年和2022年进行保存率调查,调查试验地的造林保存率、成活树苗的生长量和健康状况。2011年对林冠下造林2年的红松幼树进行光合速率观测,用CIRAS-2光合作用测定系统,对红松幼树上的2年生针叶进行林地现场环境活体测定,采用林地自然光结合LED红外光源进行光合速率测定。光源设置自低到高的光合有效辐射强度0、20、50、100、200、400、600、800、1000、1200、1400μmol/m2·s。测定时段在5月中旬至6月下旬的上午,记录各光照强度下光合参数,包括光合有效辐射(Q)、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(E)、细胞间隙C02浓度(Ci)、相对湿度(rH)、叶片温度(Ti)。2022年进行样地光辐射空间分布规律调查,在观测样地内按10m×10m均匀布点,在保持灌层和草本植被自然状态的条件下,观测地面草本层、灌木层以及林冠下的光合有效辐射强度,并记载每个观测点的乔木冠层状态,灌木层和草本层的高度、盖度和稠密度。
3 结果与分析
3.1 自然森林环境条件下红松净光合速率数学模型建立
首先建立红松净光合速率与光合有效辐射强度(光强)的回归模型作为自然森林环境条件下红松光合响应的导向曲线。通过模型拟合初算,选择更符合拟合散点图趋势的对数曲线方程作为红松光合响应导向曲线的数学模型,模型参数见表1,拟合曲线图见图1:

图1 红松光合响应导向曲线图

表1 红松光响应导向曲线参数

式中,fPnd(Q)为光合响应导向曲线方程函数;Q为光合有效辐射强度;A、B、C为模型参数。
红松光响应导向曲线方程函数的回归相关系数达到0.9(表1)取得良好效果,但图1显示,回归拟合还存在一定的偏差,说明其他环境因子对红松幼树的净光合速率有很大的影响。
通过多元相关初算确定对红松光合速率有显著影响的环境因子为相对湿度和叶片温度。因此我们假设红松光响应导向曲线方程估值的变异主要来源于相对湿度和叶片温度,用公式(1)对原始计算数据的净光合速率进行估算,并将估算值与实测值的差值作为因变量,相对湿度(定量因子)和叶片温度(定性因子)为自变量,建立排除光强因素外其他环境因子对净光合速率影响的回归数量化模型。
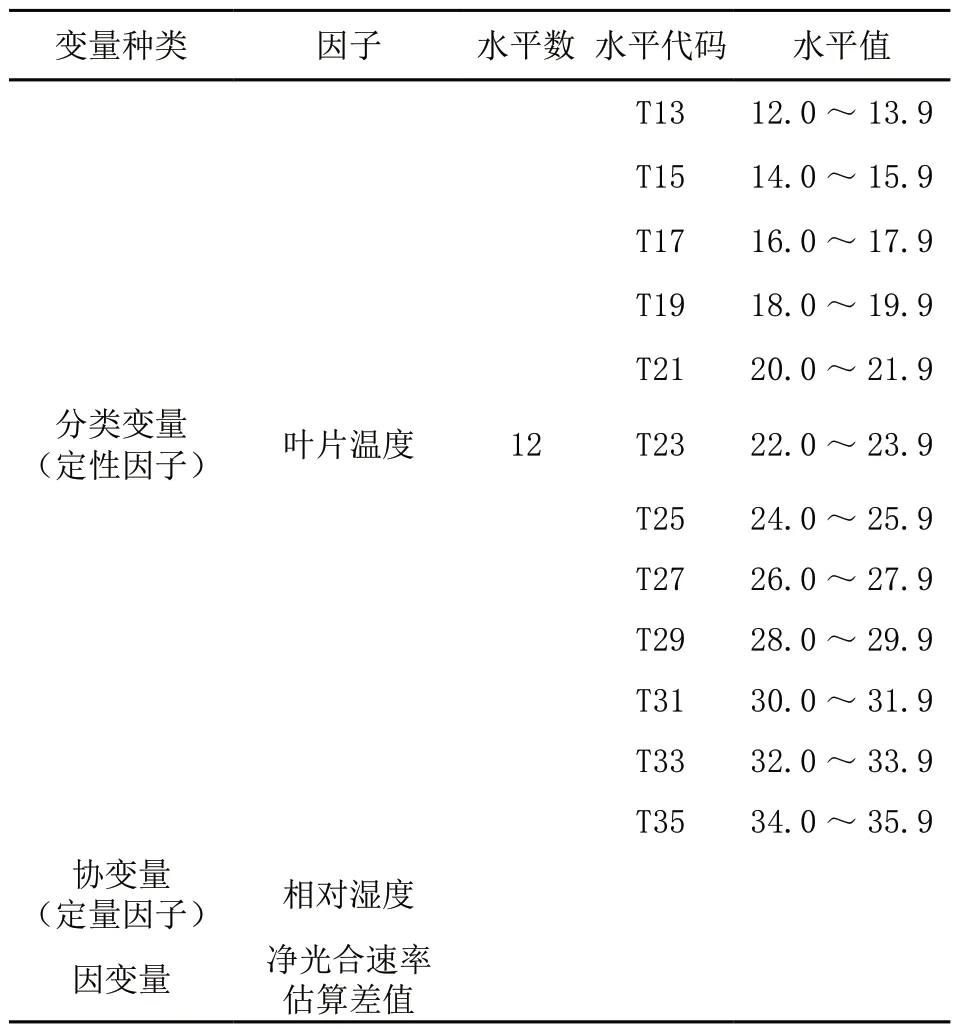
表2 数量化模型变量因子及水平
方差分析显示(表3),叶片温度和相对湿度对红松净光合速率影响显著,数量化模型有效。数量化模型因子得分表(表4)显示,红松净光合速率与相对湿度呈线性正相关。

表3 数量化模型方差分析

表4 数量化模型因子(水平)得分
由表4可知,叶片温度在13~19℃范围的得分系数处于较高的平台,从21℃开始随着温度上升得分系数急速下跌。说明叶片温度对红松幼树光合速率的影响是分阶段、非线性的,高温是影响红松幼树光合速率的限制因子。
采用多项式模型对表4中叶片温度得分系数进行回归拟合,回归方程:

式中,fT(T)为叶片温度数量化模型得分系数方程函数;T为叶片温度。
相关系数R = 0.9842。
表明回归效果良好。回归曲线图见图2,曲线呈倒S形。

图2 数量化模型得分系数拟合曲线图
以叶片温度数量化模型得分系数方程函数代替叶片温度因子得分系数值,形成连续型数量化模型回归方程:

式中,fΔPn(rH,T)为数量化模型方程函数;fT(T)为数量化模型得分值方程函数;rH为相对湿度;T为叶片温度。
将式(1)与式(3)组合形成红松净光合速率回归复合模型:

式中,fPn(Q,rH,T)为净光合速率模型方程函数;fPnd(Q)为光合响应导向曲线方程函数fΔPn(rH,T)为数量化模型方程函数;Q为光合有效辐射强度;rH为相对湿度;T为叶片温度。
3.2 次生林光辐射调查
由表5可知,次生林的林冠类型因树种、林隙和郁闭度的不同,形成不同的透光率,对林冠下灌木层和草本层的发育产生复杂的影响,同时也形成各林层光辐射的重新分配。林分中局部密集的针叶树和阔叶树,形成郁闭度0.8~0.9主林层,使主林层下的光辐射强度非常微弱,灌木层和草本层的光辐射强度均小于10.0μmol/m2·s;郁闭度0.6~0.7的阔叶树林冠下,灌木层和草本层的光辐射强度可达到60.0~120.0μmol/m2·s;郁闭度0.5的针阔混交树冠层和小林隙,草本层和灌木层的光辐射强度可达到较高的150.0~420.0μmol/m2·s;而大林隙下由于乔大层缺失使主林层的光辐射强度可达到800.0μmol/m2·s以上,充足的光照使灌木层过度发育形成浓密的屏障,使灌木层和草本层的光辐射在15.0μmol/m2·s以下。

表5 不同树冠层类型光合有效辐射强度统计

表6 次生林林内光合有效辐射统计
表7的统计数据显示,次生林内光合有效辐射非常微弱,草本层的平均辐射强度只有51.0μmol/m2·s,集中分布范围0.1~39.9μmol/m2·s占70%,辐 射 强 度 大 于100.0μmol/m2·s的观测点只占14%;灌木层的平均辐射强度69.5μmol/m2·s,集中分布范围仍然在20.0~39.9μmol/m2·s,占56%的比例,辐射强度大于100.0μmol/m2·s的观测点占22%;主林层下平均辐射强度130.6μmol/m2·s,辐射强度大于100.0μmol/m2·s的观测点占62%。

表7 各林层下光合有效辐射强度分布频率
3.3 次生林内环境因子调查
统计光合测定记录中对光合速率有显著影响的环境因子温度和湿度的分布规律,结果显示叶片温度分布范围为8~34℃,平均值为21.0℃,集中分布区18~24℃占据77.5%的比例(图3);相对湿度的分布范围为5%~16%,平均值为12.1%,集中分布呈双波峰状态,5%~7%和15%~16%的相对湿度分别占36.6%和46.3%的比例(图4)。

图3 中片温度分布图

图4 相对温度分布图
3.4 自然环境条件下红松幼树光补偿点计算
温度和湿度对红松幼树的光合速率均有显著影响,因此在自然环境中红松幼树的光补偿点是随环境因子的变化而不断变化的,结合环境因子的主要分布范围,选取环境因子平均值时(叶片温度21.0℃、相对湿度12.1%)与环境因子最差时(叶片温度24.0℃、相对湿度6.0%),用红松净光合速率回归复合模型公式(4)计算绘制自然环境红松光响应曲线拟合图(图5)。

图5 自然环境红松光响应曲线
图5显示,选取环境因子平均值时(叶片温度21.0℃、相对湿度12.1%)光响应曲线与X轴的交点(光补偿点)约在光强0μmol/m2·s处。选取环境因子最差时(叶片温度24.0℃、相对湿度6.0%)光响应曲线与X轴的交点(光补偿点)约在光强108.0μmol/m2·s处。因此,小兴安岭次生林下红松幼树在5~6月份的光补偿点为0~108μmol/m2·s。此期间红松幼树在100μmol/m2·s光辐射强度以下处于生长衰弱和停滞状态。
3.5 红松造林保存率跟踪调查
汇总次生林下红松更新造林当年、第4年、第13年的保存率及生长量。
造林保存率调查(表8)显示,次生林中人工更新的红松在幼林抚育条件下保存率达到87.6%~90%,而造林13年的保存率调查结果平均只有19.8%。分析其原因红松造林的幼林抚育年限是5年,造林5年后林分中清林作业清除的小乔木和灌木迅速萌生,恢复对红松幼树的遮盖状态,使林分重新回到满足红松幼树生长的植苗点只占林分面积14%~22%的状态。同时调查显示,红松造林苗2~3年内顶芽被狍、鹿啃食断头率高达28.4%~38.1%,反复的啃食掐头严重影响红松造林苗生长演替的进程。

表8 红松造林保存率统计
4 讨 论
4.1 通过建立红松净光合速率回归复合模型在提高模型估算精度的同时,合理梳理出环境因子光辐射强度、叶片温度、相对湿度对红松净光合速率影响的趋势及权重。红松幼树的净光合速率与光合有效辐射强度呈自然对数曲线正相关、与相对湿度呈线必正相关。叶片温度对净光合速率的影响较大,与净光合速率呈倒S形负相关。以该模型依据自然环境因子主要组合估算的红松幼树光补偿点为0~108μmol/m2·s,次生林中光合有效辐射为100μmol/m2·s以下的地段不适宜红松幼树的生长演替。
4.2 次生林主林层的树种组成树冠分布及郁闭度,影响林内灌木层和草本层的盖度和光辐射强度,光辐射强度大于100.0μmol/m2·s适合红松幼树生长的地段仅占林地面积的14%~22%。造林的清林作业及造林后经过5年的幼林抚育,红松幼树的树高还在50.0cm以下,还没有超出1.0~1.5m灌木层的影响,缺乏后续透光抚育使红松幼林在造林13年后的保存率仅有8%~26.8%,形成社会资源的巨大浪费。同时,狍、鹿啃食红松嫩枝反复形成28.4%~38.1%断头率,影响红松幼树生长演替进程也是造成保存率降低的重要因素。
4.3 毋庸置疑栽针保阔是恢复阔叶红松林生态系统最有效的途径,但次生林内红松人工更新造林的保存率受立地适生性、动物干扰等生态因素的制约,需要完善更合理的工艺措施来确保更新造林的保存率。森林演替过程与红松更新过程的契合性有其内在的自然规律,受森林演替阶段的深刻影响同时也是一个漫长的过程,采用简单的林冠下红松人工植苗更新造林,很难一次性达到栽针保阔的目标。采取包括林分选择、植苗点选择、鼠害生态防治在内的红松直播造林技术进行高效低碳的更新造林方式更具可操作性。或者通过向次生林引入红松种源扩散林方式,以近自然的方式人工促进红松天然更新,形成红松种群恢复的长效机制。
