白脂素在代谢综合征、雄性生殖和运动中的研究进展*
2023-02-16吕红艳衣雪洁
吕红艳 杨 旸 常 波 衣雪洁
(1)辽宁师范大学体育学院,大连 116029;2)上海体育学院运动科学学院,上海 200438;3)沈阳体育学院运动人体科学学院,沈阳 110102;4)沈阳体育学院运动与健康研究中心,沈阳 110102)
代谢综合征(metabolic syndrome,MetS)是指人体的蛋白质、脂肪、碳水化合物等物质发生代谢紊乱的病理状态,是一组复杂的代谢紊乱症候群,主要包括腹部肥胖、胰岛素抵抗(insulin resistance,IR)、血脂异常和血压升高等因素,并显著增加了患2型糖尿病(type 2 diabetes mellitus,T2DM)和心血管疾病的风险[1]。此外,MetS还会对男性生殖潜能产生负面影响[2]。最近研究显示,代谢性疾病死亡人数逐年上升,已呈现年轻化趋势[3]。肥胖和IR是MetS的中心环节,脂肪组织过度积累引起机体代谢失衡是诱发肥胖和IR的重要因素。脂肪组织不仅是储存和动员脂质的重要调节器官,而且还能通过内分泌作用,作为重要的传感器参与调控全身能量平衡,以及通过合成和分泌一系列的脂肪细胞因子与其他代谢器官相互交流,参与调节其他组织的功能。近年来,随着肥胖率和不孕率的平行上升,以及多种脂肪因子(如瘦素、脂联素、鸢尾素、抵抗素)的发现,脂肪因子在参与调节MetS和生殖中的潜能受到了更多的关注。本课题组前期关于瘦素和脂联素的研究也证实了脂肪因子在肥胖症和雄性生殖系统中的调控作用[4-6]。
2016年美国贝勒医学院Atul R. Chopra团队在研究新生儿早衰症(neonatal progeroid syndrome,NPS)时,发现了白色脂肪细胞分泌的一种新蛋白质分子,将其命名为白脂素(asprosin,ASP)。ASP的发现引起了广泛关注,为MetS的治疗提供了新的思路。此外,ASP与生殖功能也具有一定联系。运动通过调控机体的能量平衡,可以预防和缓解MetS的发生和发展以及提升男性/雄性生殖潜能。因此,通过对ASP与代谢综合征、生殖及运动干预的文献进行系统梳理,从ASP的生物学功能,ASP对代谢综合征的调控机制和研究现状,ASP对男性/雄性生殖的影响,运动与ASP的关系等方面进行阐述,为改善代谢综合症和提升男性/雄性生殖潜能提供新的思路。
1 ASP概述
1.1 ASP的结构和功能
2016年,Romere等[7]研究发现,NPS患者FBN1基因突变,患者脂肪代谢障碍,血糖正常,但胰岛素和血清ASP水平显著性降低。ASP是原纤维蛋白(FBN1编码)的C端裂解产物,由FBN1的65号和66号外显子编码。ASP有3个N连接的糖基化位点和其他潜在的翻译后修饰,哺乳动物中ASP的分子质量为30 ku,重组ASP(recombinant asprosin,rASP)分子质量为17 ku[7]。研究证实ASP参与葡萄糖稳态、食物摄入量和能量稳态的调节[7-8]。循环中的ASP浓度随禁食而升高,随进食而急剧下降,表现出了昼夜节律性。单次注射rASP可立即引起血糖升高,引起高胰岛素血症,但rASP对血液分解代谢激素(如胰高血糖素、儿茶酚胺和糖皮质激素)水平没有影响[7]。在IR的小鼠模型和肥胖人群中发现,ASP水平呈病理性增加,使用ASP特异性抗体可以降低IR小鼠的血清ASP浓 度,并 改 善 胰 岛 素 敏 感 性[7,9]。Duerrschmid等[8]研究发现,循环中的ASP可以穿过血脑屏障,通过Gα蛋白(Gαs)-环磷酸腺苷(cyclic adenosine monophosphate,cAMP)-蛋白激酶A(protein kinase A,PKA)轴,直接激活下丘脑中的摄食AgRP+神经元活性,并间接抑制厌食POMC神经元活性,引起食欲增加,促使肥胖发生,因此,ASP参与维持机体的能量平衡。此外,ASP还可以作用于其他组织发挥调控作用,如抑制白色脂肪褐化、促进成脂[10],抑制胰腺β细胞自噬和促进β细胞炎症[11],降低骨骼肌胰岛素敏感性[12],以及调控雄性生殖功能[13-14]。
1.2 ASP及其受体Olfr734的来源和表达
研究发现,人体脂肪组织中的FBN1mRNA表达最高,细胞实验证实脂肪是ASP的主要来源[7]。利用CRISPR/Cas9技术制备小鼠和兔子FBN1NPS/+模型,结果显示,FBN1NPS/+表型动物的体重和脂肪量显著性减少[8,15],而FBN1NPS/+表型小鼠可以免受高脂饮食诱导的肥胖症和糖尿病(DM)的发展[8],表明了ASP在维持体重、脂肪和葡萄糖稳态中的重要作用。尽管脂肪组织是ASP的主要来源,但ASP发挥作用的靶器官为肝脏。研究证实,ASP通过与肝脏中的未知G蛋白偶联受体(G protein-coupled receptors,GPCRs)结 合,激 活cAMP-PKA轴,导致肝糖迅速释放到循环中[7]。嗅觉受体(olfactory receptors,ORs)代表了GPCRs家族中最大的亚族,主要在嗅上皮和嗅球中高度表达,基因测序显示,ORs在外周组织(如心脏、肺、肠、皮肤和睾丸)中也高度表达[16-18]。2019年,Li等[19]研究发现,嗅觉受体734(Olfr734)是肝脏中ASP的受体,通过cAMP途径调节肝糖水平,但Olfr734基因敲除并不能完全消除ASP对葡萄糖产生的刺激作用。2020年,Kocaman等[20]检测了大鼠中ASP的定位和表达,结果表明,ASP位于肝脏中的肝细胞、肾皮质远端小管细胞、心脏心肌细胞、胃表面上皮细胞、睾丸间质细胞和大脑皮层神经元。综上所述,新型脂肪细胞因子ASP在多种组织中表达,ASP可通过血液循环与其他器官相互交流,在代谢稳态和疾病中发挥调节作用。
2 ASP与MetS
2.1 ASP与肥胖
临床研究检测了血清中ASP浓度与肥胖的关系。Wang等[21]对体脂指数(body mass index,BMI)>35 kg/m2的肥胖成年人和正常BMI成年人研究发现,肥胖者的血清ASP水平明显高于正常BMI组,肥胖者减肥手术6个月后,血清ASP下降水平只与体重下降百分比呈显著正相关;Ugur等[22]对116名不同体重级别的成年人采集血液和唾液样本检测ASP水平,发现ASP水平随BMI的增加而增加,表明成年人体重的变化是ASP水平波动的因素,唾液样本可以代替血清来研究其他代谢疾病与ASP之间的关系。对肥胖儿童血清ASP水平研究显示,肥胖组ASP浓度低于正常组,ASP水平在肥胖儿童中存在性别差异,即肥胖组男孩ASP水平低于肥胖组女孩[23-24],与成年人研究结果的不一致,可能是人群肥胖程度不同,ASP具有比食欲和体重控制更复杂的功能,以及肥胖儿童可能处于代谢平衡的代偿期,因此表现为ASP水平与BMI呈负相关;此外,机体产热、脂肪氧化、脂质代谢和遗传等因素可能是ASP在肥胖中存在性别差异的原因。儿童长期肥胖也会诱发IR和非酒精性 脂 肪 肝 (nonalcoholic fatty liver disease,NAFLD),研究报道肥胖儿童的血清ASP水平高于正常体重儿童[25],IR儿童的血清ASP水平高于非IR 儿童,血清ASP水平与腰臀比和胰岛素抵抗指数 (homeostasis model assessment of insulin resistance index,HOMA-IR)正相关,无性别差异[26];Liu等[27]研究发现,肥胖儿童血清ASP水平明显升高,特别是NAFLD儿童;这与肥胖成年人的研究结果一致,表明降低ASP水平可能预防肥胖人群发生IR和NAFLD;肥胖儿童ASP水平出现不一致的结果,这可能与实验对象年龄、肥胖程度、样本量大小以及实验试剂等存在一定关系,还需要更多的实验证明ASP在肥胖儿童中的水平。
动物实验研究中,Ma等[28]研究发现,与正常组小鼠相比,肥胖小鼠血清中ASP、肿瘤坏死因子α(tumor necrosis factor-alpha,TNF-α)和脂肪组织中ASP、TNF-α mRNA水平均显著性升高,脂肪组织中过氧化物酶体增殖物激活受体γ(peroxisome proliferators activated receptor-γ,PPARγ)蛋白质水平显著增加,脂肪褐变指标解偶联蛋白1(uncoupling protein 1,UCP1)显著性降低;Miao等[10]研究显示,与对照组相比,冷刺激(4℃)组和高脂组小鼠脂肪组织中ASP的mRNA表达显著性降低,这与肥胖小鼠和人类血清ASP升高的结果相矛盾,皮下白色脂肪过表达ASP时发现,冷刺激组小鼠代谢减弱,这可能是ASP水平负向调节白色脂肪组织;检测db/db(敲除瘦素受体)小鼠发现,血清ASP和脂肪组织中ASP的mRNA水平均显著性升高,这与以往的研究结果一致,可能是由于瘦素受体基因敲除引起负反馈调控阻断,机制研究显示,过表达ASP的小鼠在4℃环境中抑制了核因子E2相关因子(nuclear factorerythroid 2 related factor 2,Nrf2)的蛋白质表达,引起UCP1和过氧化物酶体增殖物激活受体γ辅激活 因 子 1α (peroxisome proliferator-activated receptor-γcoactivator-1α,PGC1α)的mRNA和蛋白质表达水平显著性下降,并促进成脂基因表达,因此,ASP可以抑制脂肪组织的褐变和产热,加剧脂肪细胞中的脂质沉积。
综上所述,血清和唾液均可以检测到ASP,不同肥胖人群中ASP水平波动趋势有所不同,ASP可能存在负反馈调节,这表明了ASP在肥胖中的复杂作用。目前,ASP对肥胖的调控机制研究较少,仅有的研究显示ASP通过降低脂肪褐变,引起肥胖,但尚不清楚ASP水平升高是肥胖的后果还是病态肥胖后的保护性反馈机制,有待深入研究。
2.2 ASP与糖尿病(DM)
机体能量平衡的维护和葡萄糖稳态的保持离不开脂肪组织的参与和调节,因此脂肪因子在DM的发病病程中扮演着重要的角色。临床研究显示,糖耐量受损者和T2DM患者血清ASP水平与空腹血糖(fasting blood glucose,FBG)、甘 油 三 酯(triglyceride,TG)、糖化血红蛋白(hemoglobin A1c,HbA1c)和IR等呈正相关[29-31];T2DM患者服用降糖药物后显著性降低血清ASP水平,改善糖脂代谢[32-33];Zhang等[34]研究显示,糖耐量正常人的血清ASP水平下降与葡萄糖耐量试验开始时一致,而在T2DM患者中ASP下降幅度减弱并出现节律紊乱,这表明血清ASP对葡萄糖波动的反应减弱可能是T2DM致病的原因之一。ASP在禁食或空腹时会刺激肝糖释放,预期低血糖时ASP会有代偿性升高,在低血糖发作期起到调节血糖作用。Groener等[35]诱导1型糖尿病(type1 diabetes mellitus,T1DM)患者可控低血糖症状,发现患者未出现生理性ASP升高现象,患者胰岛素敏感性较低,伴有IR和NAFLD,这表明未能预期升高血糖是因为患者伴有IR和NAFLD后导致ASP反应迟缓;Ke等[36]研究表明,NAFLD患者存在IR会出现血清ASP水平升高。以上研究表明,T1MD和T2DM中的ASP浓度与患者发生IR和血脂异常有关,并且相关性是一致的,因此未来更详尽的研究ASP与IR、NAFLD之间的关联,可能在DM及其并发症的治疗中起到有效作用。
在动物实验中,敲除FBN1基因构建杂合子FBN1NPS/+小鼠,发现小鼠血清ASP水平下降,高脂饮食喂养不会引起FBN1NPS/+小鼠肥胖和DM症状[8],这表明ASP是促进DM发展的重要因子。Hekim等[37]对小鼠腹腔注射rASP,发现ASP提高了对照组小鼠血糖水平,但DM组小鼠血糖无变化,降低了DM组小鼠肝脏中的TG、总胆固醇(total cholesterol,TC)和低密度脂蛋白(low density lipoprotein cholesterol,LDL-C)水平,这表明病理状态下,血糖平衡紊乱,ASP不能发挥生理上的升糖作用,这与人体实验结果一致,但ASP治疗可以减弱血脂异常。Kocaman等[20]研究显示,单次注射链脲佐菌素(streptozocin,STZ)诱导大鼠发生T1DM后,酶联免疫吸附试验(ELISA)和免疫组化分析显示,血清和肝脏中ASP水平显著性降低。而Ko等[38]同样方式诱导大鼠T1DM,却发现肝脏ASP蛋白水平显著性升高,肝脏中磷酸化腺苷酸活化蛋白激酶(phosphorylated AMP-activated protein kinase,p-AMPK)、磷酸化蛋白激酶B(phosphorylated protein kinase B,p-Akt)、PGC1α和过氧化物酶体增殖物激活受体γ辅激活因子1β(peroxisome proliferator-activated receptor-γ coactivator-1β,PGC1β)蛋白质水平显著降低,这表明调节ASP的水平或其下游的通路可能是治疗T1DM靶点,二者肝脏中ASP水平出现了矛盾的结果,这可能与STZ用量、检测方式具有一定关系。单次诱导大鼠T1DM可能出现过高的胰岛素血症,为了保护机体免受高胰岛素血症的影响,可能出现了ASP减少的现象。总之,这些差异的原因尚不清楚,解决ASP在代谢中的争议,可能要从ASP产生的途径进行全面深入的研究。此外,ASP还与糖尿病肾病指标呈正相关[39-41],并且T2DM和糖尿病肾病患者血清白介素-6(interleukin-6,IL-6)和TNF-α水平显著性升高[42],探讨ASP与炎症因子的上下游关系及其机制研究可能是治疗T2DM的途径之一。
综上所述,DM患者中,血清ASP水平升高,与IR、糖脂代谢标志物和炎症因子相关,循环ASP可能是成为DM前期、T2DM和糖尿病肾病的潜在治疗靶点,但以上研究限于因果关系,其机制研究还有待完善。动物实验中T1DM大鼠的ASP水平变化结果并不一致,这可能与实验设计有一定关系;病理状态下肝脏中升高的ASP水平影响了肝脏的能量代谢通路,今后还需要更多的研究去揭示ASP作用的靶点。
2.3 ASP与胰岛素抵抗(IR)
不同器官对胰岛素刺激的反应减弱引发了IR。脂肪因子可以增强或削弱胰岛素的作用。人体血液中大部分葡萄糖被骨骼肌利用,因此骨骼肌对葡萄糖的摄取能力是改善高血糖的重要条件。Jung等[12]对小鼠骨骼肌和C2C12细胞研究显示,ASP干预通过促进蛋白激酶Cδ(protein kinase Cδ,PKCδ)磷酸化和核转位,抑制了肌浆网Ca2+ATPase 2b(sarcoplasmic reticulum Ca2+-ATPase 2b,SERCA2b)的mRNA水平,引发了炎症反应和内质网应激(endoplasmic reticulum stress,ERS),降低了胰岛素受体底物1(insulin receptor substrate 1,IRS-1)和Akt的蛋白质水平,抑制了葡萄糖摄取,发生了骨骼肌IR,这表明通过抑制ASP可以治疗IR;Zhang等[43]研究表明,ASP激活了C2C12细胞中p-AMPK蛋白表达,上调的p-AMPK增强了葡萄糖转运蛋白4(glucose transporters 4,GLUT4)的蛋白质水平,促进了葡萄糖的摄取,AMPK抑制剂可降低ASP介导的葡萄糖摄取效应,表明AMPK-GLUT4途径介导了骨骼肌细胞生理上的ASP升糖作用。
胰岛β细胞功能受损是诱发DM的重要因素。Lee等[11]研究显示,棕榈酸和rASP干预胰腺β细胞,发现ASP诱导Toll样受体4(Toll like receptor-4,TLR4)表达和c-Jun氨基末端激酶(c-Jun N-terminal kinases,JNK)磷酸化,促进β细胞炎症和活性氧(ROS)的产生,降低了胰岛素的分泌,证明内源性ASP升高影响胰岛β细胞功能,在内源性ASP基础上,外源性ASP增加会进一步加剧β细胞功能障碍,充分说明了ASP与IR密切相关;Wang等[44]研究显示,与对照组相比,高糖和ASP处理β细胞引起p-AMPK mRNA和蛋白质表达降低,磷酸化雷帕霉素靶蛋白(phosphorylated mammalian target of rapamycin,p-mTOR)mRNA和蛋白质表达升高,抑制β细胞自噬,促进β细胞凋亡,这表明了自噬参与了ASP对β细胞的调节。临床研究显示,血清ASP水平与IR、MetS和IL-6显著正相关[45]。Nam等[46]通过生物信息学、体外和体内实验研究人胰腺癌细胞发现,胰腺癌细胞可以分泌ASP,并且ASP水平显著高于对照组,血清ASP水平可能成为胰腺癌的早期标志物。
综上所述,骨骼肌发生IR可能是ASP引起了内质网应激,减弱了葡萄糖摄取,从骨骼肌摄取葡萄糖的角度来看,针对ASP治疗可能对IR和T2DM具有有益的作用;可通过提升β细胞自噬机制,改善ASP诱发的胰岛β细胞功能受损;肝脏和脂肪组织同样对胰岛素有较强的敏感性,因此对于局部组织发生IR的研究可能有助于了解ASP更多的分子靶点和生理功能。
2.4 ASP与多巢卵囊综合征(polycystic ovary syndrome,PCOS)
PCOS是常见的内分泌、代谢异常引起的疾病,PCOS可分为IR和高雄激素血症(hyperandrogenemia,HA)两种亚型,会增加患T2DM的风险。临床研究表明,PCOS和T2DM女性患者的血清ASP水平显著高于健康受试者,PCOS组血清ASP与BMI、FBG、HbA1c、HOMAIR、睾酮(testosterone,T)、游离雄激素指数(free androgen index,FAI)呈正相关,血清ASP水平是PCOS的独立危险因素[47-49];但也有研究显示了不同的结果,Chang等[50]研究显示,与对照组相比,PCOS患者的血清ASP无显著变化,血清ASP水平与PCOS患者的IR和HA均无关,这种结果的不同,可能肥胖才是ASP变化的根本原因,而非PCOS引起的差异;Jiang等[51]研究显示,血清ASP水平在PCOS组和对照组之间没有差异,在PCOS组,血清ASP与BMI、空腹胰岛素、HOMA-IR、甘油三酯、T和黄体生成素(luteinizing hormone,LH)呈负相关,调整 BMI后,这种相关性不显著,ASP仅与催乳素(prolactin,PRL)呈正相关;将PCOS患者分为HA(PCOS-NHA和PCOS-HA)组 和IR组(PCOS-NIR和PCOS-IR),与PCOS-NHA和PCOSNIR组相比,PCOS-HA和PCOS-IR组的血清ASP水平显著性降低,这与之前的研究结果也矛盾,可能是血清ASP水平受到性激素、特定人群条件和样本混杂因素的影响,也可能是PRL通过某些反馈机制影响PCOS患者的ASP水平。因此,ASP是否可以作为预测和诊断PCOS的生物标志物还有待进一步研究。
综上所述,PCOS患者的血清ASP水平变化并不能得出统一的结论,相互矛盾的结果可能是由于受试者年龄、测量学差异、样本量、疾病严重程度和遗传等因素之间的差异引起的,需要更大的样本量和长时间随访跟踪调查去进一步探究ASP和PCOS的关系,并通过机制研究去揭示ASP在PCOS中的作用。
3 ASP与雄性生殖功能
代谢综合征引起的能量失衡也参与了调节了男性/雄性生殖功能,因此,脂肪分泌的ASP在男性生育能力中的作用也引起了人们的关注。Kocaman等[20]研究显示,与对照组相比,DM组大鼠睾丸间质细胞(leydig cell)的ASP水平显著性增加,睾丸间质细胞是睾酮合成的场所,DM通常伴随低睾酮水平,因此,病理状态下ASP与睾酮水平可能具有负向关系。但目前关于ASP与睾酮生物合成的机制研究还未见报道;ASP受体之一Olfr734在睾丸中高表达[14,19]。Wei等[14]研究表明,与野生型(wild type,WT)小鼠相比,10周龄雄性Olfr734-/-小鼠精子渐进性运动显著性减弱,将雄性WT小鼠和Olfr734-/-小鼠分别与雌性WT小鼠交配,发现Olfr734-/-小鼠的受精能力显著性降低,但小鼠的血清睾酮水平和交配频率不受影响,这说明Olfr734介导的生殖途径不同于睾酮介导的途径,也可能睾丸中还存在ASP的其他受体;进一步研究显示,rASP干预WT小鼠和Olfr734-/-小鼠后,ASP只提高了WT小鼠的血糖和精子中的ATP和cAMP的mRNA水平以及精子渐进性运动能力和受精率,Olfr734-/-小鼠无变化,rASP干预还可以将40周龄WT小鼠的以上指标恢复至10周龄小鼠水平,这表明ASP或Olfr734的激动剂是潜在的提高生育能力的药物,Olfr734也是肝脏中ASP的受体,激活Olfr734信号可能会增加高血糖的风险。因此,寻找特异性激活睾丸中Olfr734的信号很重要。另外,高脂饮食降低了WT和Olfr734-/-小鼠的精子渐进性运动,Olfr734-/-小鼠下降的更显著[14],这表明ASP-Olfr734信号轴对高脂饮食引起的精子活力降低有保护作用。Keskin等[13]对雄性大鼠侧脑室注射ASP,研究显示,与对照组和假手术组相比,ASP治疗组(34 nmol/L rASP和68 nmol/L rASP)增加了下丘脑促性腺激素释放激素(gonadotropin releasing hormone,GnRH)mRNA和蛋白质表达,提高了血清卵泡刺激素(follicle stimulating hormone,FSH)、LH和T水平,还提高了睾丸中精子密度、精子活动率和渐进性精子活力。从整体上看,ASP通过激活下丘脑-垂体-睾丸(hypothalamic-pituitary-testicular,HPT)轴提升了生殖功能。
综上所述,生理状态下,ASP可以提高雄性小鼠生殖功能,而病理状态下(如肥胖)ASP的增多会抑制生殖潜能,但这种生殖潜能的下降还需要敲除FBN+/-基因进一步确认,Olfr734受体的敲除并不能完全解释ASP基因对生殖的作用;病理状态下,ASP参与调节了生殖功能下降可能是机体发生了ASP抵抗,病理状态下再给予ASP干预可能通过负反馈机制提升生殖功能,但需要实验研究证实这种猜想;此外,ASP对睾酮的影响是否只通过中枢,而不通过外周,也需要更多的实验研究进一步探讨。
4 运动与ASP
运动或身体活动通过能量消耗产生能量赤字来影响能量平衡。Schumann等[52]研究显示,与健康成年人相比,肥胖受试者血清ASP水平与之相似,力竭运动对健康成年人和肥胖成年人血清ASP水平无影响。Wiecek等[53]研究了单次20 s自行车冲刺运动对健康成年男性和女性血清ASP的影响,研究显示,运动后只有女性血清中ASP和鸢尾素水平升高,瘦素水平下降;无论性别,血清ASP与脂联素呈正相关,与瘦素呈负相关,这表明多种脂肪因子相互关联,运动对脂肪因子具有一定影响,ASP水平可能在个体和性别中存在一定差异,因此,不同形式的无氧运动对ASP水平的影响还有待进一步研究。Ceylan等[54]研究显示,与对照组相比,超重/肥胖组(男性)血清ASP、脂质运载蛋白2(lipocalin-2,LCN2)和胰岛素水平升高,神经肽Q(spexin,SPX/NPQ)水平下降,早、晚30 min中等强度有氧运动后,两组(正常组和超重/肥胖组))体温升高,血清ASP、LCN2和胰岛素水平均显著性下降,SPX水平无差异,超重/肥胖组各指标变化更显著,此外,晚间有氧运动比早上运动效果更明显,这表明中等强度运动可以减少ASP水平,晚间效果更显著可能是有氧运动和晚间心理上抑制能量摄入共同导致能量负平衡,伴随体温升高,抑制了食欲;Zarei等[55]研究显示,12周联合抗阻有氧运动降低了T2DM男性患者血清ASP浓度和BMI、FBG、IR和HbA1c水平;Kantorowicz等[56]研究显示,8周最大脂肪氧化强度的步行训练降低了患有代谢障碍的女性血清中的ASP浓度,改善内脏型肥胖症状。这些研究表明,长期运动可以改善肥胖状态下的ASP水平,运动训练后ASP浓度的下降可能是由于体重和脂肪百分比下降所致。
在动物实验中,Ko等[38]研究显示,与对照组相比,T1DM大鼠肝脏中ASP蛋白水平升高,p-AMPK、p-Akt、PGC1α和PGC1β蛋白水平降低,8周跑台运动后,T1DM大鼠肝脏ASP、PKA和转化生长因子β(transforming growth factor-β,TGF-β)蛋白质水平,增加了p-AMPK、p-Akt、PGC1β和锰超氧化物歧化酶(manganese superoxide dismutase,MnSOD)蛋白质水平,这表明有氧运动通过激活AMPK信号通路(运动诱导的主要能量感受器),减少ASP和PKA/TGF-β,降低血糖,改善 T1DM;Nakhaei等[57]研究了持续性和间歇性8周游泳训练对MetS雄性大鼠血清ASP水平的影响,研究显示,与MetS组相比,运动训练组大鼠血清ASP水平和血脂(TG、TC、LDL-C)水平显著性降低,间歇训练组对ASP浓度的降低作用大于持续性训练,但没有统计学差异。
综上所述,力竭运动对成年人不同状态(生理/病理)的血清ASP水平影响并不一致,还可能存在性别差异,这可能与运动方案(如运动方式、持续时间和强度、参与者特征、评估参数等)有关;而有氧运动和联合抗阻运动会降低疾病状态下血清ASP水平,但还需要更多的实验去探讨运动对ASP的机制研究。
5 总结与展望
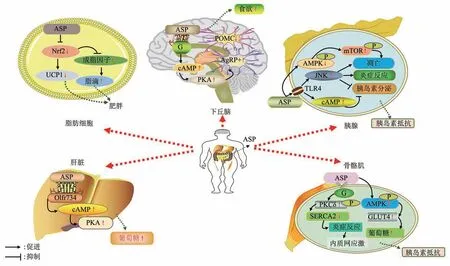
Fig. 1 Schematic diagram of ASP regulatory signal pathways in different tissues of the human body图1 人体不同组织中ASP调控信号通路的示意图
ASP作为新发现的脂肪因子,调节脂肪功能,还经循环到达肝脏、下丘脑、胰腺、肌肉等多个组织器官,调控机体多种生命活动(图1)。大多数研究表明,成年人病理状态下(如肥胖和T2DM)血清ASP水平较高,降低ASP表达可能为代谢性疾病的治疗提供了潜力和新思路。脂肪因子诱发的能量失衡对男性/雄性生殖功能具有一定负面影响。目前,关于ASP对男性/雄性生殖系统的影响研究较少,因此,还需要更多的实验探究ASP在生殖中的作用机理。
运动在改善代谢综合征中的作用已经得到充分的认可。因此,运动对ASP的影响在防治代谢性疾病方面可能具有广阔前景,但目前运动对ASP影响的研究文献较少,其机制也尚未明确,增加运动对ASP影响的研究数量可能有助于理解运动后ASP减少或增加的机制。此外,探究ASP与其他脂肪因子(如脂联素和瘦素)之间的联系,以及它们在代谢综合征和生殖系统中是否存在交叉对话,将有助于更好理解能量失衡对生殖功能的调控机制。