黄土高原种子植物多样性分布格局与环境解释
2023-02-10安克丽李育榕熊东阳李国庆
安克丽,李育榕,熊东阳,刘 冠,李国庆①
(1.西北农林科技大学林学院,陕西 杨凌 712100;2.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100;3.中国科学院水利部水土保持研究所,陕西 杨凌 712100)
物种丰富度地理分布格局及其影响因素一直是宏观生态学和生物地理学研究的核心问题之一[1-2],随着全球生物多样性的急剧丧失[3],其也成为保护生物学关心的问题[4]。物种丰富度地理格局受到许多环境因素(如气候、地形、土壤等因素)影响[5],也与研究尺度[6]、物种类群[7]以及物种的生理生态需求等有关[8]。在此基础上,很多研究者将人为因素对物种分布格局的影响也纳入其中[9]。在区域尺度上探究物种分布规律及其影响因素,能为识别物种优先保护区和制定物种保护规划提供参考[10]。
黄土高原位于我国中部偏北,海拔在800~3 000 m之间,总面积约为64万km2,包括中国太行山以西、青海省日月山以东、秦岭以北、长城以南的广大地区。黄土高原年平均气温在4~14 ℃之间,年降水量在150~650 mm之间且由东南向西北递减,属于干旱大陆季风气候区,是东南湿润季风气候向西北内陆干旱气候过渡、暖温带落阔叶林向典型草原和荒漠草原过渡的地带[11]。该区植物种类丰富且起源古老,黄土高原种子植物约有130科864属3 224 种(包括亚种、变种、变型)[12-13]。长期以来,针对黄土高原植物分布格局的研究主要集中在省级和自然保护区尺度。比如,王翠红等[14]研究了山西省种子植物多样性格局与环境因子的关系;汤萃文等[15]研究甘肃省特有种子植物的分布格局并划定其多样性优先保护区;周哲等[16]研究了太白山自然保护区攀援植物物种丰富度及其沿海拔梯度的分布格局等。近年来,在黄土高原尺度上陆续开展研究。比如,董雪蕊等[17]研究了基于系统发育的黄土高原地区木本植物热点区域;刘丽君等[18]基于系统保护规划研究了黄土高原木本植物分布格局及其优先保护区的识别。到目前为止,黄土高原物种分布格局研究主要聚焦于小尺度,在黄土高原尺度上的研究仍然比较缺乏,且聚焦的种子植物类型比较单一。因此,还需要在黄土高原尺度上对比较丰富的种子植物类群分布格局进行研究,从而为黄土高原种子植物保护提供建议。
黄土高原是世界上最大的黄土堆积区,也是我国关键生态脆弱区和国家重点土壤保持功能区[19]。长期以来,由于人为破坏和自然环境的变迁,水土流失严重,使得该地区生境破碎化严重的同时生物多样性保护也面临着严峻形势[20]。建立自然保护区是全球范围内公认的应对生物多样性下降危机的战略[21]。自1965年,黄土高原建立第1个自然保护区——太白山自然保护区以来,已建成148个自然保护区,其面积约占黄土高原土地面积的9.8%。自然保护区保存了较为完整的生态系统,是生物多样性聚集的核心区域,也是研究物种分布格局及其驱动因子的理想自然试验地,已有的自然保护区考察报告等研究文献也为自然保护区内动植物物种丰富度数据获取提供了便利[22-23]。
基于此,以黄土高原为研究区域,通过保护区文献、自然保护区标本馆等收集到黄土高原108个自然保护区的种子植物调查数据,结合气候、生境异质性以及人类干扰等数据进行研究,拟解决以下两个科学问题:(1)探讨黄土高原种子植物丰富度地理格局及其影响因素;(2)明确气候、生境异质性以及人为干扰对黄土高原种子植物丰富度地理格局影响的相对重要性,确定维持其格局的环境解释。在气候变化和人为干扰加剧的背景下,笔者研究能为黄土高原生物多样性保护、自然保护区建设提供基础数据和理论支持。
1 材料与方法
1.1 自然保护区数据
黄土高原自然保护区数据主要来源于中华人民共和国生态环境部发布的《2017年全国自然保护区名录》,根据黄土高原矢量边界最终统计出黄土高原148个自然保护区(附录Ⅰ)。其中,国家级36个,省级97个,市级5个,县级10个,涵盖森林生态、草原草甸、内陆湿地、荒漠生态、野生动物、野生植物、地质遗迹和古生物遗迹等类型。在148个保护区中,能收集到物种数据的有108个,主要是国家级和省级自然保护区,包括森林生态、野生动物、野生植物、荒漠生态、内陆湿地和草原草甸等生态类型,108个保护区中具有矢量边界的保护区有44个,矢量边界缺乏、以中国生态环境部发布的自然保护区名录中的面积为基准、以自然保护区的中心经纬度坐标为圆心画圆来表示的有64个(图1)。黄土高原矢量边界数据来源于国家地球系统科学数据中心黄土高原分中心(http:∥loess.geodata.cn)。

图1 黄土高原148个自然保护区分布格局
1.2 物种分布数据
种子植物分布数据主要来源于相关自然保护区文献、中国自然保护区标本资源共享平台(http:∥www.papc.cn/)和保护区调查规划报告等实际调查数据,共搜集108个自然保护区的物种数据,建立了“黄土高原自然保护区种子植物分布数据库”(完整的数据源清单见附录Ⅱ)。未收集到数据的保护区有40个,主要是市县级、古生物遗迹和地质遗迹类型保护区。这些保护区级别较低,建设目的主要是为了保护古生物遗迹和地质遗迹,因此,保护区物种调查数据缺失。由于面积会影响物种丰富度,为了比较各个自然保护区种子植物丰富度,采用种子植物密度表征不同自然保护区种子植物丰富程度。物种数量与面积之间通常呈指数关系,基于面积、物种丰富度,采用以下方程计算每个自然保护区的种子植物密度[24]。
(1)
式(1)中,S为自然保护区种子植物丰富度,种;A为自然保护区面积,km2;D为自然保护区种子植物密度,种·km-2。黄土高原108个自然保护区种子植物密度见图2。

图2 黄土高原种子植物密度空间分布格局
1.3 环境因子数据
所使用的环境因子变量有3类11个(表1):(1)气候因子,年平均温度(AMT)、年降水量(AP)、最干月降水(PDM)、温度季节性(TSD)、降水范围(Ap.R)、温度范围(Amt.R);(2)生境异质性因子:海拔范围(Alt.R)、净初级生产力范围(Npp.R);(3)人类干扰因子:人口密度(HPD)、人类足迹(HF)、国内生产总值(GDP)。

表1 黄土高原种子植物丰富度以及各环境变量的基本统计信息
统计每个自然保护区的上述环境数据,气候数据来源于世界气候数据网站(https:∥www.worldclim.org/),其中,AMT、AP、PDM和TSD提取的是保护区内的均值,Amt.R和Ap.R来源于自然保护区内AMT、AP的最大值与最小值间差值,用于表征一个地区的气候复杂程度[25]。生境异质性因子中海拔数据、净初级生产力数据来源于中国科学院资源环境科学与数据中心(https:∥www.resdc.cn/),海拔数据空间分辨率为250 m×250 m,Alt.R和Npp.R分别为自然保护区内海拔及净初级生产力最大值和最小值之差,Alt.R用于表征一个地区地形复杂程度[8],而Npp.R可用于表征一个地区的能量变化[26]。人类干扰因子中的HPD数据来源于Diva GIS网站(http:∥www.diva-gis.org/gdata); HF是利用1960到2001年收集的人口密度、土地转化和基础设施等代表人类活动的空间数据集构建的指数,代表人类活动对地表影响的强度,数据来源于中国科学院资源环境科学与数据中心(https:∥www.resdc.cn/);GDP数据主要来源于资源学科创新平台数据库(http:∥www.data.ac.cn/),提取自然保护区均值作为每个自然保护区的人类干扰数据。
1.4 统计分析
首先,采用单因素相关分析研究各环境因子对黄土高原种子植物丰富度和密度格局的独立影响程度;其次,通过多元逐步回归得出最优线性模型并筛选出决定种子植物丰富度和密度格局的主要环境因子;最后,利用方差分解法(variation partitioning analysis)进一步分解气候、生境异质性、人类干扰对黄土高原种子植物丰富度和密度格局的相对作用。
在进行统计分析前,对种子植物丰富度和密度进行平方根(sqrt)转换,对生境异质性因子中海拔范围、降水范围、温度范围,人为干扰因子中国内生产总值、人口密度进行自然对数(ln)转换,以解决数据的非正态分布问题。由于物种分布数据和环境数据之间普遍存在显著的空间自相关,在进行显著性检验时会得到偏向于更加显著的结果[27]。因此,在进行单因素相关分析时,采用改进的t检验方法检验相关系数的显著性[28]。最优线性回归模型采用R中的step函数,结合赤池信息准则(Akaike information criterion,AIC)确定[29]。以校正后的模型拟合优度(adjustedR2)作为最终解释率,通过模型中各变量的偏决定系数(partialR2)比较不同环境因子对模型的贡献。最后,通过方差分解探讨气候、生境异质性和人类干扰3组变量对种子丰富度和密度的独立解释率及共同解释率。
采用R.3.6.1进行统计分析,其中,多元逐步回归采用step函数,改进的t检验方法采用spatial pack程序包中的modified.ttest函数,方差分解采用vegan程序包中的varpart函数。
2 结果与分析
2.1 种子植物丰富度及密度与环境因子的单因素相关分析
黄土高原种子植物密度由东南向西北递减(图2)。总体上,在陕西和河南南部边界秦岭一带较高,宁夏和内蒙古西北片区较低。种子植物丰富度及密度与各环境因子的单因素相关分析结果(表2[27])显示:黄土高原种子植物丰富度及密度与AP、PDM、TSD、Ap.R、Amt.R、Alt.R、Npp.R、HPD、HF呈显著正相关,而与AMT、GDP相关性不显著。其中,种子植物丰富度与净初级生产力范围相关性较强(Npp.R:R=0.622,P<0.001),与年降水量、最干月降水、温度季节性、海拔范围相关性次之(AP:R=0.588,P<0.001;TSD:R=0.580,P<0.001;PDM:R=0.567,P<0.001;Alt.R:R=0.517,P<0.001),与降水范围、温度范围、人口密度和人类足迹相关系数低于上述环境因子(Ap.R:R=0.412,P<0.001;Amt.R:R=0.461,P<0.001;HPD:R=0.410,P<0.001;HF:R=0.334,P<0.001)。种子植物密度与年降水量、温度季节性、净初级生产力范围相关性较强(AP:R=0.624,P<0.001;TSD:R=0.618,P<0.001;Npp.R:R=0.600,P<0.001),与最干月降水和海拔范围相关性次之(PDM:R=0.586,P<0.001;Alt.R:R=0.507,P<0.001),与降水范围、温度范围、人口密度和人类足迹相关系数低于上述环境因子(Ap.R:R=0.396,P<0.001;Amt.R:R=0.443,P<0.001;HPD:R=0.456,P<0.001;HF:R=0.367,P<0.001)。

表2 黄土高原种子植物丰富度及密度与环境因子的相关分析[27]
总体上,气候因子中与降水相关的环境因子与种子植物丰富度和密度相关性较强,生境异质性因子中的净初级生产力范围和海拔范围与种子植物丰富度和密度显著相关,人为干扰因子整体与种子植物丰富度和密度相关系数较低,影响较小。
2.2 种子植物丰富度及密度与环境因子的最优线性模型
最优线性模型分析结果(表3)表明,所有环境因子中,PDM、Alt.R、Npp.R和HF的线性组合对黄土高原种子植物丰富度格局的解释率最高,达53.4%。偏回归分析结果显示,在种子植物丰富度的最优线性模型中,Alt.R、PDM和Npp.R影响最为强烈,其偏决定系数分别为0.158、0.145和0.142(表3),此外,HF也有一定贡献。对于种子植物密度的分布格局,所有环境因子中,AP、PDM、Alt.R、Npp.R和HF解释率共达52.0%。偏回归分析结果显示,在种子植物密度的最优线性模型中,AP、Alt.R和Npp.R影响最强烈,其偏决定系数分别为0.389、0.257和0.360(表3)。此外,PDM和HF对种子植物密度的分布格局也有一定程度的贡献。

表3 黄土高原种子植物丰富度及密度与环境因子的最优多元线性模型
综上所述,气候因素中降水因子及生境异质性是黄土高原种子植物丰富度和密度格局的主导因子,人为干扰有一定程度的影响。
2.3 黄土高原气候、生境异质性和人为干扰对种子植物丰富度格局的相对影响
方差分解分析结果显示,气候、生境异质性和人为干扰3者的组合能够解释黄土高原种子植物丰富度格局52.0%~53.4%的变异,有46.6%~48.0%的变异不能解释(图3)。
在3组因子中,生境异质性在种子植物丰富度格局中起着重要作用(独立解释率为12.3%~17.2%);而气候因子的单独解释率为7.3%~9.3%;相较于气候和生境异质性,人类干扰因子的独立解释重要性较低(独立解释率为2.4%~4.0%)(图3)。气候和生境异质性之间的交互作用最为重要(解释率为17.7%~18.6%),3者之间的交互作用能解释13.2%~15.0%的总变异(图3)。
3 讨论
近几十年来,随着全球生物多样性保护研究的深入,在不同区域尺度上物种分布格局及其影响因素受到生态学家的广泛关注[30]。物种多样性的地理分布格局形成过程是一个复杂的生态过程,受到许多因素影响,比如面积[6]、气候[22]和景观异质性[7]等。黄土高原种子植物密度最高的自然保护区主要集中在东南部秦岭狭长地带,总体上,种子植物密度由东南向西北递减。笔者选取包括气候、生境异质性和人为干扰因素的11个变量对黄土高原种子植物多样性格局进行研究。结果表明,AP、PDM、Alt.R、Npp.R和HF是影响黄土高原种子植物分布的最主要因素(表3)。这说明黄土高原种子植物分布格局受多种环境因子的共同影响,它们共同决定了黄土高原种子植物丰富度格局。
3.1 气候因子对黄土高原种子植物多样性分布格局的解释
气候因子主要包括热量和水分两个基本组成部分:热量能为生物体提供能量,也是维持其基本生命活动的基础;水分是生命体的基本组成部分,也是生理代谢活动的必要物质[31]。因此,这在很大程度上决定了生物的分布及其生理活动,从而影响其分布格局。在大尺度上,气候因子决定植物多样性分布格局,已得到广泛认可[32],但在特定区域尺度和生物类群中,起决定性作用的是哪类气候因子还未达成共识[33]。在气候因素方面,笔者研究结果表明与水分相关的环境因子以及温度季节性与种子植物多样性呈显著正相关(表2)。在气候因子中,在最优线性模型中只有与水分相关的因子被纳入,且能解释黄土高原种子植物密度39.0%的变异(表3,图3)。在干旱环境中,水分因子是限制植物分布格局的关键因素[34]。黄土高原处于我国半干旱和干旱地带,热量充足,降水分布不均匀,水分匮乏[35],水分成为制约该区域植被生长的关键因素。SHI等[36]对黄土高原草本植被进行年轮解剖发现,其生长对干旱胁迫有较为敏感的响应,说明水分条件对黄土高原植被生长有着重要作用;董雪蕊等[17]提出水分是黄土高原半干旱地区植物生长与植被恢复的主要限制因子。黄土高原种子植物密度分布格局由东南向西北递减,与等雨量线相对应。在400 mm等雨量线东南部种子植物密度比西北部高,尤其是靠近秦岭一带(陕西南部和河南北部)(图2)。位于黄土高原南部的秦岭山脉,年降水量在600 mm以上,水热条件良好,是我国生物多样性保护的热点区域之一[34],所以,此区域的自然保护区拥有黄土高原最高的种子植物密度。
3.2 生境异质性因子对黄土高原种子植物多样性分布格局的解释
以往研究表明,生境异质性也会对生物多样性分布格局产生深刻影响。生境异质性越高的地区,其栖息地类型就越多样,可利用的资源也越丰富,从而能提供更多的生态位促进物种生存[37];当外界环境改变或生存条件不利时,生境异质性高的地区能够为生物提供避难所,提高物种存活的几率[38];生境异质性越复杂的地区还可以通过生境隔离使物种分化几率增加[39]。在生境异质性因素方面,Alt.R和Npp.R所表征的生境异质性可解释种子植物密度43.1%的变异。黄土高原Alt.R较大的自然保护区主要分布在600 mm等雨量线的南部边界、青海和宁夏西北部边界以及整个山西(图4)。Alt.R 较大的山区能为物种提供类型多样的栖息地和资源,导致生态位多样性增加,从而促进物种形成[37];黄土高原植物类群丰富,起源古老且地理成分复杂[12],Alt.R较大的山区能够缓冲外界环境和不利条件的变化,为物种提供避难所,提高存活几率[38],保留更多古老的植物类群。其次,环境能量的增加能够提高一个地区的净初级生产力(NPP),增加生物量积累,能为动物提供更多的食物;其最终结果使得该区域更多生物个体能共存,动植物种群规模增大,最终提高该区域物种多样性[40]。Npp.R能用于表征一个地区能量的复杂状况,因此,能显著影响黄土高原种子植物分布格局(表2~3)。笔者研究结果与孙远等[41]利用被子植物和陆栖脊椎动物潜在分布范围数据对黄河流域进行研究得出的结果相似,他们也发现生境异质性(海拔范围、冠层高度范围、NPP范围)相对于其他因素对黄河流域潜在的植物和动物分布格局的环境解释贡献最高。LI等[42]也证明生境异质性(海拔范围)是影响全国省域尺度上种子植物分布格局的最重要因素。XU等[43]基于全国2 376个县30 519种维管植物和565种哺乳动物的研究结果也表明,在全国县域尺度上Alt.R表征的生境异质性对物种丰富度至关重要,建议在生物多样性保护工作中要重视栖息地的异质性。另外,笔者利用自然保护区生物多样性实际调查数据与孙远等[41]利用的物种潜在分布范围叠加后的物种多样性数据得出了相似的生物多样性格局,说明黄土高原和黄河流域生物多样性分布格局的一般规律没有被强烈的人类活动干扰破坏。

图4 黄土高原自然保护区海拔高差
3.3 人为干扰因子对黄土高原种子植物多样性分布格局的解释
人类活动对生物多样性的影响包括自然资源的不合理利用、森林的滥砍滥伐、外来物种的引入、环境污染、城市化以及气候变化等,这些都对生物多样性产生巨大压力,导致生物多样性急剧下降[44]。在人为干扰因素方面,笔者研究结果表明,HF显著影响黄土高原种子植物密度的分布格局,其表征的人为干扰因素能够解释种子植物密度12.5%的变异,并产生负面影响(表2~3,图3)。并且, HF分别与HPD和GDP呈显著正相关关系(R2=0.333,P<0.001;R2=0.079,P=0.003)(图5),表明HPD增长与GDP总值提高对黄土高原HF增加有直接影响,可用HF表征人类干扰。人口增加和城市扩张使生态系统遭到破坏、野生动植物生境面积减少,也导致生境破碎化严重进而降低连通性,最终使生态系统变得脆弱,从而增大生态系统被入侵和生物多样性丧失几率[45]。同时,在退耕还林等国家政策驱动下,黄土高原建立了大面积人工林和果园林[46-47],从而导致区域生境同质化,这在一定程度上会降低地区间beta多样性和物种多样性的周转,从而对生物多样性保护产生负面影响。
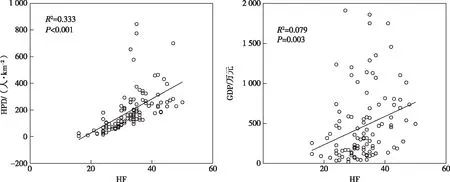
图5 人类足迹(HF)与人口密度(HPD)和国内生产总值(GDP)的相关关系
3.4 研究的不确定性、应用和建议
尽管气候、生境异质性和人为干扰解释了黄土高原种子植物丰富度大部分的变异,但是仍然有46.6%~48.0%的变异未得到解释(图3)。这说明气候和生境异质性是黄土高原种子植物多样性的决定因子,人为干扰有一定影响,但其他未知潜在因素也起重要作用。杨涛等[48]对大理苍山种子植物多样性分布格局影响因子的研究得出该区域种子植物多样性主要受气候、土壤及地形的共同影响;历史气候变化也可能会影响物种分布格局,邹东廷等[49]发现末次盛冰期以来的气候对中国蔷薇科物种分布和物种多样性格局具有显著影响,且对西北、东北和青藏高原地区的影响要大于南部地区。李亦超等[50]也发现末次盛冰期以来的气候变化显著影响欧亚大陆东部毛茛科植物多样性格局。方差分解结果表明其他未被引入的土壤、地形、进化以及地质历史气候因素等都可能对黄土高原种子植物丰富度有重要作用。其次,黄土高原到目前为止没有较新且系统的植被数据集,而自然保护区网络较为完善且数据相对齐全,基于自然保护区的方式是目前研究物种多样性格局最好的方式。自然保护区生物多样性高,是保护生物多样性和维持物种多样性的关键区域,适合于研究植物物种丰富度与环境因子的关系。未来应对自然保护区多样性数据进行系统收集并优化环境数据,进一步引入土壤、历史气候等因素,深入探究其他未知因素的作用过程,从而更准确、全面地研究黄土高原种子植物多样性格局。
黄土高原已建立的自然保护区面积占比还远低于全国水平,在现存保护区的基础上进行调整和补充是黄土高原生物多样性保护的重要途经。笔者研究中,生境异质性、气候和人类干扰对黄土高原种子植物密度的影响分别为43.1%、39.0%和12.5%。因此,对黄土高原未来生物多样性的保护策略应该以增加生境异质性和保护自然植被资源为主,同时要减少人为干扰。笔者研究结果表明,黄土高原山地、水热条件良好的地区有较高的物种丰富度,在这些区域建立自然保护区并最大程度减轻人为干扰,是保护种子植物多样性的有效途径。同时,笔者研究与孙远等[41]和XU[43]等的研究内容类似,都是关于保护区域alpha多样性目标的研究,对于beta多样性较高而alpha多样性较低的区域启示意义较小。因此,认为对于黄土高原区域beta多样性分布规律和保护政策方面还需要进一步研究。
4 结论
基于自然保护区种子植物丰富度的研究表明,黄土高原植物多样性由东南向西北逐渐递减,气候、生境异质性和人为干扰能够用于揭示黄土高原植物分布格局的大部分变异;降水因子、海拔范围、净初级生产力范围和人类足迹是影响黄土高原植物多样性分布的最主要因素。在黄土高原自然保护区建设和生物多样性保护规划中,应该科学地考虑和纳入生境异质性研究其对生物多样性的影响,自然保护区应建立在水热条件良好且人为干扰较少的山地,将有利于保护植物多样性。笔者研究是关于保护区域alpha多样性目标的研究,对于beta多样性较高而alpha多样性较低的区域启示意义较小。建议加强自然保护区物种名录或植被调查数据库的建设,为黄土高原区域beta多样性分布规律和保护政策的研究提供数据支撑。
