不同番茄品种叶片理化性状对南美番茄潜叶蛾寄主选择性的影响
2023-02-06薛增昇张生存郄杏桃吕治燊张东霞郑卫锋闫喜中
薛增昇, 王 韩, 张生存, 马 力, 郄杏桃, 吕治燊,张东霞, 郑卫锋, 闫喜中,*, 郝 赤,*
(1. 山西农业大学植物保护学院, 太谷 030801; 2. 山西省植物保护植物检疫中心, 太原 030001)
南美番茄潜叶蛾Tutaabsoluta属鳞翅目(Lepidoptera)麦蛾科(Gelechiidae),又名番茄麦蛾、番茄潜麦蛾或番茄潜叶蛾(Povolny, 1994; 马菲等, 2011)。南美番茄潜叶蛾起源于南美洲西部的秘鲁,20世纪50年代以来一直是南美洲国家露地和保护地番茄的重要害虫(Desneuxetal., 2010)。2006年传入西班牙(Urbanejaetal., 2007),随后迅速遍布欧洲各国并快速扩散至亚洲和非洲大陆(Desneuxetal., 2010, 2011)。截至2017年5月,该虫已在包括南美洲、欧洲、非洲、中美洲、亚洲在内的85个国家/地区发生和22个国家/地区疑似发生(Biondietal., 2018; 张桂芬等, 2018)。2017年8月在我国新疆地区首次监测到南美番茄潜叶蛾(张桂芬等, 2019),之后迅速向我国其他地区扩散,对我国番茄产业造成严重威胁。
南美番茄潜叶蛾主要危害茄科植物,尤其嗜食番茄,包括鲜食番茄和加工番茄;该虫还可危害马铃薯、茄子、甜椒、烟草和人参果等茄科作物(Desneuxetal., 2010)。南美番茄潜叶蛾主要以幼虫进行危害,可以在番茄植株的任一发育阶段和任一地上部位进行危害(张桂芬等, 2018)。幼虫一经孵化便潜入寄主植物组织中,取食叶肉,并在叶片上形成细小的潜道,通常早期不易被发现,隐蔽性极强;当虫口密度比较高、幼虫龄期比较大时,还可蛀食顶梢、腋芽、嫩茎以及幼果。在该虫发生地,倘若不予防治或防治不及时,将造成80%~100%的番茄产量损失(Desneuxetal., 2010),对当地番茄产业造成毁灭性打击。
番茄是世界上重要的蔬菜作物,全球年总产量达1.7亿吨,在蔬菜作物中位居首位(李君明等, 2021)。不同番茄品种由于其理化性状的不同也造就了其抗虫能力的差异。例如研究人员发现,南美番茄潜叶蛾在不同番茄品种上的生命表参数有显著差异,其更喜欢取食对其敏感性较弱的番茄品种(Gharekhani and Salek-Ebrahimi, 2014;Cekin and Yasar, 2015);同时也有研究表明不同番茄品种上的南美番茄潜叶蛾种群丰度也有差异(El-Kadyetal., 2018),然而,最近的一项研究表明不同番茄品种对南美番茄潜叶蛾产卵选择性影响不显著(Sawadogoetal., 2022)。我国是鲜食和加工番茄生产大国,鲜食番茄产量稳居世界第一,加工番茄产量位居世界第二或者第三(李君明等, 2021)。我国番茄有许多栽培品种,而国内关于不同番茄品种叶片理化性状对南美番茄潜叶蛾寄主选择性影响的研究还鲜有报道。千禧、大粉、大黑、小绿和黄矮生5个番茄品种在我国种植广泛且理化性状差异显著,因此,本研究以南美番茄潜叶蛾为研究对象,测试了南美番茄潜叶蛾对千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的取食选择性和产卵选择性,并检测了5个番茄品种叶片的理化性状,最后分析了南美番茄潜叶蛾寄主选择性与番茄叶片理化性状的内在联系,为番茄抗虫品种选育以及南美番茄潜叶蛾的绿色精准防控提供理论依据。
1 材料与方法
1.1 供试昆虫
南美番茄潜叶蛾采自山西省晋中市太谷区申奉村试验田(37°25′51.7584″N, 12°36′16.362″E),并在山西农业大学昆虫神经行为与感官生物学实验室饲养3代以上,饲养温度为(25±1) ℃,相对湿度为65%,光周期为16L∶8D。将幼虫养到3龄用于取食选择性测定。将蛹区分雌雄之后分装于干净指型管中,每个指型管分装1头蛹,待其羽化后用于产卵选择性测定。
1.2 供试植物
5个番茄品种(千禧、大粉、大黑、小绿和黄矮生)种苗均购自山东寿禾种业有限公司,并放置于山西农业大学昆虫神经行为与感官生物学实验室培养,培养温度为(25±1) ℃,相对湿度为65%,光周期为16L∶8D,待其长到40 cm时用于试验。
1.3 南美番茄潜叶蛾取食选择性测定
参照赵瑞等(2022)的方法,采用叶碟法测定南美番茄潜叶蛾幼虫对千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的取食选择性。在同一时间剪取5个番茄品种植株中层嫩度一致的健康叶片,将各番茄品种叶片叶柄用带水脱脂棉包住保湿,随机排放在铺有网格纸且直径为20 cm的玻璃培养皿(均分为5个区域)中围成一圈,中心放置一个直径为5 cm的培养皿,将饥饿处理12 h后的3龄幼虫接入培养皿中,分别于接虫后6, 12, 24 和48 h时观察记录各品种番茄叶片区域内的虫口数量,计算取食选择率,取食选择率(%)=(各品种番茄叶片区域内的虫口数量÷培养皿内虫口总数)×100,并使用网格法统计48 h时的取食面积,即将被取食过的各品种番茄叶片放在长×宽=1 mm×1 mm的网格纸上,画出取食面积,进而通过网格数量估算取食面积。每个处理30头幼虫,3次生物学重复。
1.4 南美番茄潜叶蛾产卵选择性测定
参照梅文娟等(2021)的方法,采用笼罩法测定南美番茄潜叶蛾对5个番茄品种的产卵选择性。选取5个番茄品种长势一致的番茄苗各1盆,随机放入养虫笼(长×宽×高=50 cm×50 cm×50 cm)内,记录每个养虫笼中各番茄品种摆放的具体位置,将羽化48 h且未交配的南美番茄潜叶蛾成虫雌雄各30头逐一从笼门小心释放到养虫笼内任其自由扩散。接虫后紧闭笼门,以防成虫逃逸,分别在接虫后24, 48和72 h时记录每株植物上的产卵量和叶片数量,试验重复3次。
1.5 番茄叶片化学性状测定
采用比色法测定番茄叶片的化学性状。
1.5.1含水量:分别称取0.1 g(准确至0.001 g)去除叶中脉的各品种中层番茄叶片放到已烘干并称重的烘皿中,置于103 ℃烘箱内(皿盖打开斜至皿边),加热4 h,加盖取出,于干燥器内自然冷却至室温称重。再置于烘箱内加热1 h,冷却后称重,重复加热1 h后称重,直至样品达到恒定重量(前后两次重量差≤0.005 g,以最小重量为准),每处理重复3次。
1.5.2可溶性糖含量:称取0.1 g去除叶中脉的各品种中层番茄叶片置于1.5 mL离心管中,加入1 mL蒸馏水研磨成匀浆,沸水浴10 min(盖紧,以防止水分散失),冷却后,8 000 g常温离心10 min,取上清液于1.5 mL离心管中,按照生工生物工程股份有限公司植物可溶性糖含量检测试剂盒(D799391-0050)说明书进行操作,使用上海元析仪器有限公司生产的UV-5500紫外分光光度计在620 nm处测定吸光值,利用标准曲线法计算各番茄品种叶片可溶性糖含量,试验重复3次。
1.5.3蛋白质含量:称取0.1 g去除叶中脉的各品种中层番茄叶片置于1.5 mL离心管中,加入1 mL蒸馏水研磨成匀浆,8 000 g常温离心10 min,取上清液于1.5 mL离心管中,按照生工生物工程股份有限公司BCA法蛋白质浓度测定试剂盒(C503021)说明书进行操作,使用上海元析仪器有限公司生产的UV-5500紫外分光光度计在562 nm处测定吸光值,利用标准曲线计算各番茄品种叶片蛋白质含量。设置3次生物学重复3次。
1.5.4氨基酸含量:称取0.1 g去除叶中脉的各品种中层番茄叶片置于1.5 mL离心管中,加入1 mL试剂1研磨成匀浆,置于沸水浴提取15 min;自来水冷却后,10 000 r/min 4 ℃离心10 min,取上清液于1.5 mL离心管,按照生工生物工程股份有限公司氨基酸(AA)含量检测试剂盒(D799583-0050)进行操作,使用上海元析仪器有限公司生产的UV-5500紫外分光光度计在570 nm处测定吸光值。设置3次生物学重复。
1.6 番茄叶片物理性状测定
采用网格法和比色法测定番茄叶片的物理性状。
1.6.1叶片面积:在同一时间分别剪取5个番茄品种上、中和下层嫩度一致的番茄叶片,用网格法测量叶片面积,即将各品种番茄叶片放在1 mm×1 mm的网格纸上,画出叶片面积,通过网格数量估算叶片面积,最终取3个部位叶片面积的均值。设置3次生物学重复。
1.6.2叶片绒毛密度:在同一时间分别在5个番茄品种上、中、下层嫩度一致的番茄叶片上取1 cm×1 cm的叶片面积,在高倍体视镜下观察记录绒毛数量,最终取3个部位绒毛数量的均值。设置3次生物学重复。
1.6.3叶绿素含量:剪取5个番茄品种中层新鲜叶片,蒸馏水清洗干净后用滤纸吸干表面水汽,然后去掉叶中脉后在避光条件下液氮研磨成粉末,称取0.1 g植物叶片粉末,按照南京建成生物工程研究所植物叶绿素试剂盒(A147-1-1)说明书进行操作,使用上海元析仪器有限公司生产的UV-5500紫外分光光度计分别在645和663 nm处读取吸光度值。设置3次生物学重复。
1.7 数据分析
参照刘欢等(2021)的方法,利用Pearson法对南美番茄潜叶蛾幼虫取食选择率和成虫产卵量与不同番茄品种理化性状进行相关性分析。数据初步整理使用Excel 2019,采用SPSS 24进行差异显著性分析和Pearson相关性分析,利用Graphpad Prism 9和ChiPlot(https:∥www.chiplot.online/)进行图形绘制。
2 结果
2.1 南美番茄潜叶蛾3龄幼虫对不同番茄品种的取食选择性
南美番茄潜叶蛾3龄幼虫对千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的取食选择性结果表明:在取食6 h时,南美番茄潜叶蛾3龄幼虫对大粉和千禧番茄叶片的取食选择率均在32%~33%之间,显著高于对其他3个番茄品种叶片的取食选择率(P<0.05),对其他3个番茄品种叶片的取食选择率都小于20%,从高到低依次为:大黑>小绿>黄矮生; 在取食12 h时, 南美番茄潜叶蛾3龄幼虫对大粉和千禧番茄叶片的取食选择率在 27%~30%之间,显著高于对其他3个番茄品种叶片的取食选择率(P<0.05),对其他3个番茄品种叶片的取食选择率都小于20%,从高到低依次为:大黑>小绿>黄矮生;在取食24 h时,南美番茄潜叶蛾3龄幼虫对大粉和千禧番茄叶片取食选择率在34%~38%之间,显著高于对其他3个番茄品种叶片的取食选择率(P<0.05),对其他3个番茄品种叶片的取食选择率都小于20%,从高到低依次为:大黑>小绿>黄矮生;在取食48 h时,南美番茄潜叶蛾3龄幼虫对大粉和千禧番茄叶片的取食选择率与取食24 h时保持一致,显著高于对其他3个番茄品种叶片的取食选择率(P<0.05),对其他3个番茄品种叶片的取食选择率都小于20%,从高到低依次为:大黑>小绿>黄矮生。对于千禧和大粉番茄来说,随着取食时间的增加,南美番茄潜叶蛾幼虫对叶片取食选择率整体呈上升趋势;对于小绿和黄矮番茄来说,随着取食时间的增加,南美番茄潜叶蛾幼虫对叶片取食选择率整体呈下降趋势;对于大黑番茄来说,随着取食时间的增加,南美番茄潜叶蛾幼虫对叶片取食选择率基本保存不变(图1)。

图1 南美番茄潜叶蛾3龄幼虫在不同时间段对不同番茄品种叶片的取食选择率Fig. 1 Feeding selection rates of the 3rd instar larvae of Tuta absoluta to the leaves of different tomato varieties in different time periods图中数据为平均值±标准误;折线上不同字母表示在同一时间不同番茄品种差异显著(P<0.05, Duncan氏检验)。Data in the figure are mean±SE. Different letters above the broken lines indicate significant difference among tomato varieties at the same time (P<0.05, Duncan’s test)
南美番茄潜叶蛾3龄幼虫在取食48 h时对千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的取食面积测定结果表明:南美番茄潜叶蛾幼虫在48 h时对大粉番茄叶片取食面积最大,为(156.67±6.67) mm2,与对小绿和黄矮生番茄叶片的取食面积[分别为(46.67±8.82)和(23.33±6.67) mm2]差异显著(P<0.05),但与对千禧和大黑番茄叶片的取食面积[分别为(133.33±13.33)和(116.67±12.02) mm2]差异不显著(P>0.05)(图2)。
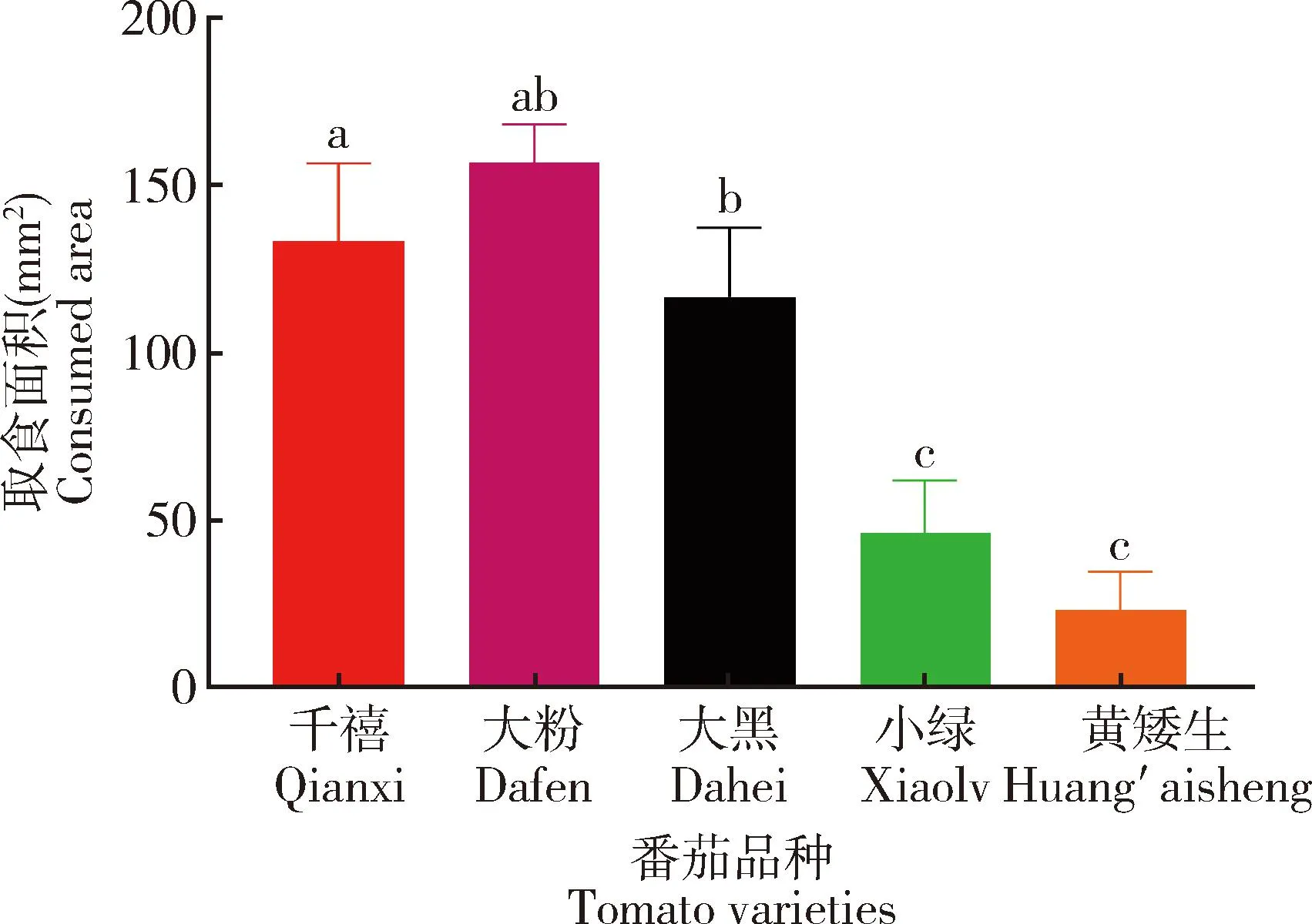
图2 南美番茄潜叶蛾3龄幼虫在取食48 h时对不同番茄品种叶片的取食面积Fig. 2 Consumed area of the 3rd instar larvae of Tuta absoluta on the leaves of different tomatovarieties at 48 h after feeding图中数据为平均值±标准误;柱上不同字母表示同一参数在不同番茄品种间差异显著(P<0.05, Duncan氏检验)。图4和5同。Data in the figure are mean±SE. Different letters above bars represent significant difference in the same parameter among different tomato varieties (P<0.05, Duncan’s test). The same for Figs.4 and 5.
2.2 南美番茄潜叶蛾成虫对不同番茄品种的产卵选择性
南美番茄潜叶蛾成虫对千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的产卵选择性结果表明:在产卵24 h时,南美番茄潜叶蛾成虫在大黑番茄每叶片上的产卵量为(7.53±0.73)粒,显著高于在其他番茄品种每叶片上的产卵量(P<0.05),在其他番茄品种每叶片上的产卵量从高到低依次为: 千禧[(2.58±1.18)粒]>大粉[(2.22±0.91)粒]>黄矮生[(1.50±0.15)粒]>小绿[(1.08±0.20)粒];在产卵48 h时,南美番茄潜叶蛾成虫在大黑番茄每叶片上的产卵量[(10.83±0.58)粒]显著高于在其他番茄品种每叶片上的产卵量(P<0.05),在其他番茄品种每叶片上的产卵量从高到低依次为:黄矮生[(5.30±0.70)粒]>千禧[(3.80±0.20)粒]>大粉[(3.52±1.03)粒]>小绿[(3.40±0.06)粒];在产卵72 h时,南美番茄潜叶蛾成虫在大黑番茄每叶片上的产卵量为(12.55±0.95)粒,显著高于在其他番茄品种每叶片上的产卵量(P<0.05),在其他番茄品种的每叶片上的产卵量从高到低依次为:黄矮生[(5.90±0.10)粒]>大粉[(4.85±0.78)粒]>千禧[(4.83±0.43)粒]>小绿[(3.89±0.11)粒];随着产卵时间增加,南美番茄潜叶蛾成虫在各个番茄品种每叶片上的产卵量都呈现上升趋势(图3)。
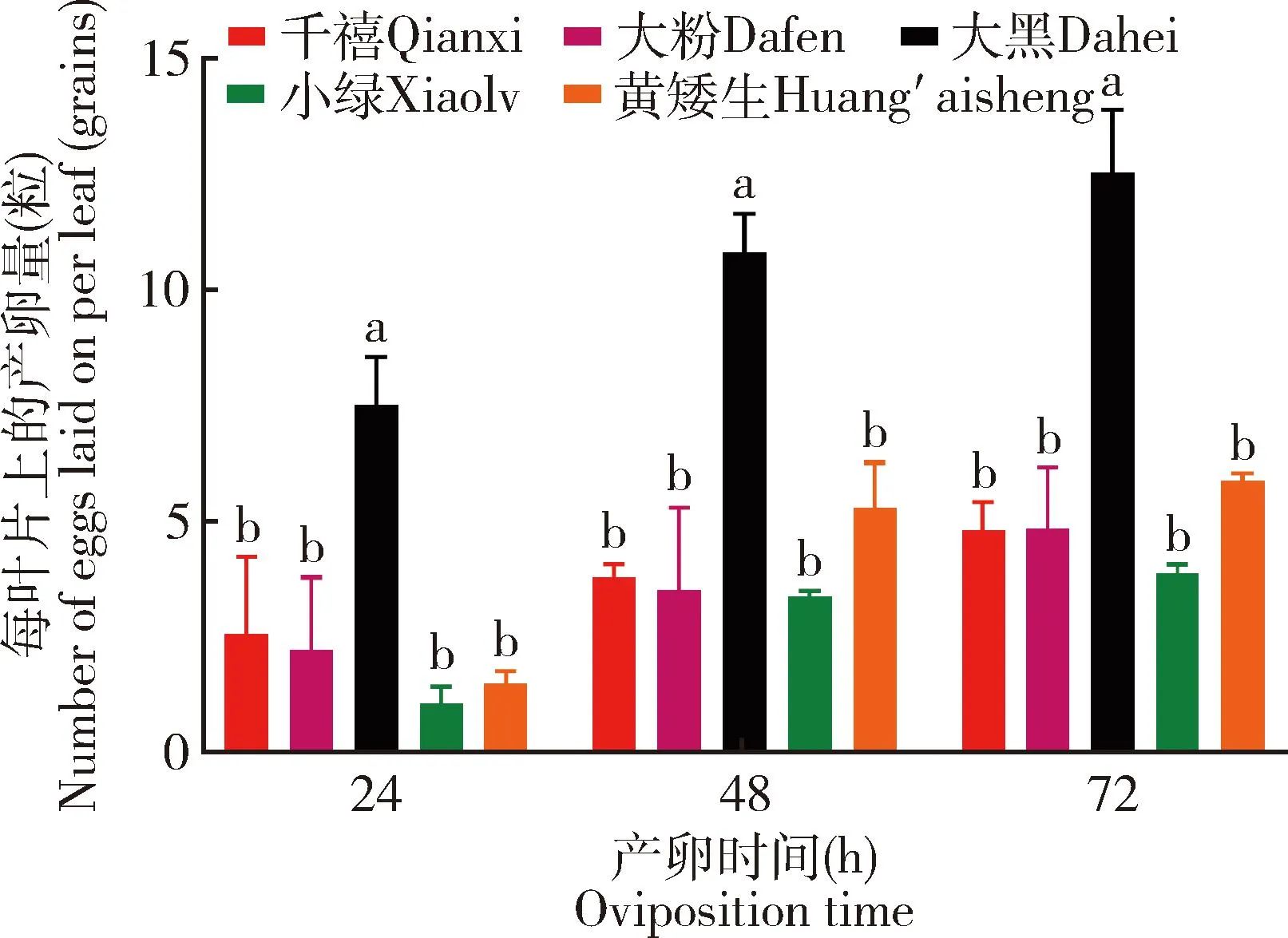
图3 南美番茄潜叶蛾成虫在不同时间段在不同番茄品种每叶片上的产卵量Fig. 3 Numbers of eggs laid by Tuta absoluta adults on per leaf of different tomato varieties in different time periods图中数据为平均值±标准误;柱上不同字母表示在同一时间不同番茄品种间差异显著(P<0.05, Duncan氏检验)。Data in the figure are mean±SE. Different letters above bars indicate significant difference among different tomato varieties at the same time (P<0.05, Duncan’s test)
2.3 不同番茄品种叶片的化学性状
千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的化学性状结果显示:黄矮生番茄叶片含水量(89.49%±0.33%)显著高于其他番茄品种叶片含水量(P<0.05),其他番茄品种叶片含水量从高到低依次为:大粉(87.63%±0.45%)>小绿(85.66%±0.64%)>千禧(84.97%±0.65%)>大黑(84.31%±0.17%)(图4: A);千禧番茄叶片可溶性糖含量[(0.22±0.04) mg/mL]显著高于其他番茄品种叶片可溶性糖含量(P<0.05),其他番茄品种叶片可溶性糖含量从高到低依次为:大黑[(0.11±0.04) mg/mL]>大粉[(0.10±0.03) mg/mL]>小绿[(0.06±0.02) mg/mL]=黄矮生[(0.06±0.03) mg/mL](图4: B);黄矮生番茄叶片蛋白质含量最高,为(309.80±12.77) μg/mL,显著高于小绿、大粉和千禧番茄叶片蛋白质含量[分别为(222.67±4.51), (209.70±30.11)和(164.21±11.12) μg/mL](P<0.05),但是与大黑[(247.55±23.96) μg/mL]叶片蛋白质含量差异不显著(P>0.05)(图4: C);不同番茄品种叶片氨基酸含量差异不显著(P>0.05),各番茄品种叶片氨基酸含量从高到低依次为:大黑[(46.00±3.31) μmol/mL]>小绿[(43.30±4.87) μmol/mL] >千禧[(37.56±3.83) μmol/mL]>黄矮生[(36.85±3.43) μmol/mL]>大粉[(32.30±3.46) μmol/mL](图4: D)。
2.4 不同番茄品种叶片的物理性状
千禧、大粉、大黑、小绿和黄矮生5个番茄品种叶片的面积、茸毛密度、叶绿素a含量、叶绿素b含量及总叶绿素含量5个物理性状的测试结果表明:黄矮生番茄叶片面积最大,为(481.33±48.31) mm2,显著高于小绿番茄叶片面积[(314.56±36.39) mm2](P<0.05),但是与大黑、千禧和大粉番茄叶片面积[分别为(407.11±25.58), (384.67±57.10)和(376.78±36.22) mm2]差异不显著(P>0.05)(图5: A); 大粉番茄叶片茸毛密度最大, 为(216.89±27.90)根/cm2,显著高于大黑番茄叶片茸毛密度[(112.22±13.14)根/cm2](P<0.05),但是与黄矮生、千禧和小绿番茄叶片茸毛密度[分别为(177.22±11.35), (167.78±18.02)和(155.44±21.96)根/cm2]差异不显著(P>0.05)(图5: B);大黑番茄叶片叶绿素a含量最高,为(1.12±0.00) mg/g, 显著高于大粉番茄叶片叶绿素a含量[(1.05±0.00) mg/g](P<0.05),但与千禧、黄矮生和小绿番茄叶片叶绿素a含量[分别为(1.10±0.00) , (1.09±0.02)和(1.07±0.03)]差异不显著(P>0.05);大黑番茄叶片叶绿素b含量[(0.99±0.05) mg/g]显著高于其他番茄品种叶片叶绿素b含量(P<0.05),其他番茄品种叶片叶绿素b含量从高到低依次为:千禧[(0.71±0.03) mg/g]>小绿[(0.68±0.10) mg/g]>黄矮生[(0.66±0.07) mg/g]>大粉[(0.57±0.01) mg/g]; 大黑番茄叶片总叶绿素含量[(2.13±0.04) mg/g]显著高于其他番茄品种叶片总叶绿素含量(P<0.05),其他番茄品种叶片总叶绿素含量从高到低依次为:千禧[(1.83±0.02) mg/g]>小绿[(1.77±0.14) mg/g]>黄矮生[(1.76±0.08) mg/g]>大粉[(1.63±0.03) mg/g](图5: C)。

图5 不同番茄品种叶片的物理性状Fig. 5 Physical characteristics of the leaves of different tomato varietiesA: 不同番茄品种叶片面积Leaf area of different tomato varieties; B: 不同番茄品种叶片茸毛密度Leaf pubescence density of different tomato varieties; C: 不同番茄品种叶片叶绿素含量 Leaf chlorophyll contents of different tomato varieties.
2.5 南美番茄潜叶蛾3龄幼虫取食选择率与不同番茄品种叶片理化性状的关系
利用Pearson法对南美番茄潜叶蛾3龄幼虫取食选择率与不同番茄品种叶片理化性状进行的相关性分析结果表明:在取食6 h时,南美番茄潜叶蛾幼虫取食选择率与不同番茄品种叶片可溶性糖含量(r=0.75,P=0.144)和叶片茸毛密度(r=0.34,P=0.571)呈非显著正相关,与叶片含水量(r=-0.35,P=0.558)、蛋白质含量(r=-0.81,P=0.098)、氨基酸含量(r=-0.44,P=0.457)、叶片面积(r=-0.27,P=0.664)、叶绿素a含量(r=-0.15,P=0.808)、叶绿素b含量(r=-0.12,P=0.854)及总叶绿素含量(r=-0.12,P=0.845)呈非显著负相关;在取食12 h时,南美番茄潜叶蛾3龄幼虫取食选择率与不同番茄品种叶片蛋白质含量(r=-0.89,P=0.041)呈显著负相关,与叶片可溶性糖含量(r=0.63,P=0.254)和叶片茸毛密度(r=0.29,P=0.633)呈非显著正相关,与叶片含水量(r=-0.48,P=0.410)、氨基酸含量(r=-0.30,P=0.622)、叶片面积(r=-0.56,P=0.330)、叶绿素a含量(r=-0.28,P=0.650)、叶绿素b含量(r=-0.12,P=0.848)及总叶绿素含量(r=-0.14,P=0.816)呈非显著负相关;在取食24 h时,南美番茄潜叶蛾3龄幼虫取食选择率与不同番茄品种叶片可溶性糖含量(r=0.80,P=0.102)和叶片茸毛密度(r=0.33,P=0.592)呈非显著正相关,与叶片含水量(r=-0.33,P=0.588)、蛋白质含量(r=-0.77,P=0.124)、氨基酸含量(r=-0.46,P=0.441)、叶片面积(r=-0.16,P=0.792)、叶绿素a含量(r=-0.07,P=0.909)、叶绿素b含量(r=-0.09,P=0.884)及总叶绿素含量(r=-0.09,P=0.886)呈非显著负相关;在取食48 h时,南美番茄潜叶蛾3龄幼虫取食选择率与不同番茄品种叶片可溶性糖含量(r=0.72,P=0.172)和叶片茸毛密度(r=0.39,P=0.519)呈非显著正相关,与叶片含水量(r=-0.27,P=0.660)、蛋白质含量(r=-0.73,P=0.157)、氨基酸含量(r=-0.50,P=0.390)、叶片面积(r=-0.17,P=0.785)、叶绿素a含量(r=-0.16,P=0.800)、叶绿素b含量(r=-0.14,P=0.829)及总叶绿素含量(r=-0.13,P=0.822)呈非显著负相关(图6)。

图6 南美番茄潜叶蛾3龄幼虫在不同时间的取食选择率与不同番茄品种叶片理化性状相关性分析Fig. 6 Correlation analysis between the feeding selection rates of the 3rd instar larvae of Tuta absoluta at different time and the physical and chemical characteristics of the leaves of different tomato varieties图中星号代表显著相关(*0.01 利用Pearson法对南美番茄潜叶蛾成虫产卵量与不同番茄品种叶片理化性状进行的相关性分析结果表明:在产卵24 h时,南美番茄潜叶蛾成虫在每叶片上的产卵量与不同番茄品种叶片可溶性糖含量(r=0.18,P=0.770)、氨基酸含量(r=0.46,P=0.440)、面积(r=0.13,P=0.832)、叶绿素a含量(r=0.59,P=0.292)、叶绿素b含量(r=0.82,P=0.086)及总叶绿素含量(r=0.80,P=0.101)呈非显著正相关,与叶片含水量(r=-0.54,P=0.346)、蛋白质含量(r=-0.01,P=0.992)和茸毛密度(r=-0.59,P=0.295)呈非显著负相关;在产卵48 h时,南美番茄潜叶蛾成虫在每叶片上的产卵量与不同番茄品种叶片叶绿素b含量(r=0.92,P=0.028)和总叶绿素含量(r=0.91,P=0.031)呈显著正相关,与叶片蛋白质含量(r=0.36,P=0.546)、氨基酸含量(r=0.65,P=0.235)、面积(r=0.35,P=0.558)及叶绿素a含量(r=0.77,P=0.128)呈非显著正相关,与叶片含水量(r=-0.38,P=0.525)、可溶性糖含量(r=-0.06,P=0.920)和茸毛密度(r=-0.78,P=0.120)呈非显著负相关;在产卵72 h时,南美番茄潜叶蛾成虫在每叶片上的产卵量与不同番茄品种叶片叶绿素b含量(r=0.91,P=0.034)和总叶绿素含量(r=0.90,P=0.039)呈显著正相关,与叶片蛋白质含量(r=0.29,P=0.631)、氨基酸含量(r=0.60,P=0.284)、面积(r=0.33,P=0.589)和叶绿素a含量(r=0.74,P=0.152)呈非显著正相关, 与叶片含水量(r=-0.41,P=0.493)、 可溶性糖含量(r=-0.01,P=0.991)和茸毛密度(r=-0.74,P=0.155)呈非显著负相关(图7)。 图7 南美番茄潜叶蛾成虫在不同时间段在每叶片上的产卵量与不同番茄品种叶片理化性状相关性分析Fig. 7 Correlation analysis between the numbers of eggs laid by Tuta absoluta adults on per leaf in different time periods and the physical and chemical characteristics of the leaves of different tomato varieties 在长期的协同进化过程中,植食性昆虫与寄主植物形成了相互适应的生存策略。对于植食性昆虫而言,凭借其自身灵敏的嗅觉、味觉、视觉、触觉等感觉器官的综合作用进行寄主植物的选择(Sunetal., 2012; Britoetal., 2016)。对于寄主植物而言,则演化出了抵御植食性昆虫攻击的防御策略,这些策略可以是物理性状的改变,例如叶片面积、叶片茸毛密度以及叶片颜色等等(Charles-Dominiqueetal., 2017; Liuetal., 2017; Belete, 2018),当然也可以是化学成分的改变,例如营养物质、次生代谢物以及植物激素等等(Mithöfer and Boland, 2012; Bruce, 2015; Belete, 2018)。自从农业出现以来,由于人类对农作物的驯化,影响了农作物的物理性状和化学成分,在一定程度上改变了植物-昆虫-天敌的三级营养关系,造成了同种植物不同品种对有害生物抗性的差异(Barrios-Masias and Jackson, 2014; Fentik, 2017)。 在本研究中,我们发现南美番茄潜叶蛾对不同品种番茄叶片取食选择性和产卵选择性均有明显差异(图1, 3),这在其他昆虫的研究中也得到了证实,如葛超美等(2018)的研究表明灰茶尺蠖Ectropisgrisescens对不同茶树品种的取食选择性有显著差异;Li等(2012)的研究表明,相较于野生烟草品种,斜纹叶蛾Spodopteralitura更喜欢在栽培烟草品种上产卵。但出乎我们意料的是南美番茄潜叶蛾幼虫取食选择性与成虫产卵选择性结果并不一致,可能是由于南美番茄潜叶蛾幼虫在进行取食选择时味觉系统起到主导作用(Yangetal., 2021),而南美番茄潜叶蛾成虫在进行产卵选择时嗅觉系统和视觉系统起到主导作用(Liuetal., 2020),所以即使是同种昆虫,在不同发育阶段味觉系统、嗅觉系统及视觉系统对同种寄主植物的感知也存在一定的差异。 随后,我们测试了不同番茄品种叶片的理化性状(图4, 5),并分析了南美番茄潜叶蛾幼虫取食选择率与不同番茄品种叶片理化性状的相关性,发现在取食12 h时,南美番茄潜叶蛾幼虫取食选择率与不同番茄品种叶片蛋白质含量呈显著负相关,与叶片含水量、可溶性糖含量、氨基酸含量、面积、茸毛密度、叶绿素a含量、叶绿素b含量和总叶绿素含量没有显著相关性,且在其他取食时间段南美番茄潜叶蛾幼虫取食选择率与不同番茄品种叶片理化性状均没有显著相关性(图6),这表明南美番茄潜叶蛾幼虫更偏向取食蛋白质含量低的番茄叶片,这与地红蝽Pyrrhocoristibialis和杜鹃冠网蝽Stephanitispyriodes喜食蛋白质含量低的寄主植物的研究结果(段宇杰等, 2020; 赵瑞等, 2022)相似,而且在前12 h南美番茄潜叶蛾幼虫取食时间越长,其取食选择率与叶片蛋白质含量的负相关显著性越大,造成这种情况的原因可能是昆虫的取食诱导了植物抗虫蛋白的表达(McKenzieetal., 2002),而这种由多种抗虫蛋白组成的防御网络是植物抵御昆虫摄食的重要机制(Fink and Scandalios, 2002; Rubioetal., 2002)。 植食性昆虫产卵选择常与其自身或后代营养需求密切相关,在一定程度上反映了昆虫与植物的相互作用关系(钦俊德和王琛柱, 2001; 梅文娟等, 2021)。在本研究中我们发现在产卵48和72 h时,南美番茄潜叶蛾成虫产卵量与不同番茄品种叶片叶绿素b含量和总叶绿素含量呈显著正相关,与叶片含水量、可溶性糖含量、蛋白质含量、氨基酸含量、面积、茸毛密度和叶绿素a含量无显著相关性(图6),这表明南美番茄潜叶蛾雌性成虫更喜欢在叶绿素含量高的寄主植物上产卵,这在植食性昆虫界是一个普遍的现象,目前已有大量的研究为这一结论提供了充分的证据,例如雒珺瑜等(2011)的研究表明绿盲蝽Polyguslucorum寄主选择行为与棉花叶片叶绿素含量呈显著正相关;刘晨曦(2004)的研究表明南美斑潜蝇Liriomyzahuidobrensis更喜欢寄生颜色浓绿的寄主植物叶片;Anjani等(2017)和戴小华等(2011)的研究表明,对于三叶草斑潜蝇Liriomyzatrifolii来说绿叶型的蓖麻是高感品种,而紫叶型的蓖麻则是抗性品种。造成这种情况的原因主要有两方面:一是视觉在昆虫产卵选择过程中发挥着极其重要的作用(Reeves, 2011),南美番茄潜叶蛾对绿色有显著趋性(张桂芬等, 2021);二是叶绿素是绿色植物的主要光受体,在植物光合作用中举足轻重,植物中叶绿素的含量在一定程度上反映了植物合成有机化合物的能力(Banu and Yadav, 2022),所以昆虫可能为最大限度保证后代的生存,优先将卵产在叶绿素含量丰富的寄主植物上(Jaenike, 1978),此外,也有研究表明,植物在受到昆虫取食之后叶绿素含量会有显著降低(Rostamietal., 2020),这可能是植物逃避昆虫再次为害的重要防御策略。 总之,本研究初步探讨了不同番茄品种叶片理化性状对南美番茄潜叶蛾幼虫取食选择行为以及成虫产卵行为的影响,发现南美番茄潜叶蛾幼虫取食选择性主要与寄主植物蛋白质含量有关,成虫产卵选择性主要与寄主植物叶绿素含量有关,为深入探究不同番茄品种对南美番茄潜叶蛾的抗生性奠定了理论基础。但是植物抗虫机制非常复杂,后续我们将进一步研究不同番茄品种挥发性物质、次生代谢物以及叶片防御酶对南美番茄潜叶蛾寄主选择行为的影响,并探究相关分子机制,以期为昆虫-植物协同进化及番茄抗虫品种选育提供新的参考。2.6 南美番茄潜叶蛾成虫在每叶片上的产卵量与不同番茄品种叶片理化性状的关系

3 讨论
