Increasing the appropriate seedling density for higher yield in dry direct-seeded rice sown by a multifunctional seeder after wheatstraw return
2023-02-03TIANJinyuLIShaopingCHENGShuangLIUQiuyuanZHOULeiTAOYuXINGZhipengHUYajieGUOBaoweiWEIHaiyanZHANGHongcheng
TIAN Jin-yu ,LI Shao-ping ,CHENG Shuang ,LIU Qiu-yuan ,ZHOU Lei ,TAO Yu ,XING Zhi-peng,HU Ya-jie,GUO Bao-wei,WEI Hai-yan,ZHANG Hong-cheng
1 Jiangsu Key Laboratory of Crop Cultivation and Physiology/Innovation Center of Rice Cultivation Technology in Yangtze Valley,Ministry of Agriculture and Rural Affairs/Co-Innovation Center for Modern Production Technology of Grain Crops,Yangzhou University,Yangzhou 225009,P.R.China
2 Agricultural College,Xinyang Agriculture and Forestry University,Xinyang 464000,P.R.China
Abstract Dry direct-seeded rice (DDR) sown using a multifunctional seeder that performs synchronous rotary tillage and sowing has received increased attention because it is highly efficient,relatively cheap,and environmentally friendly.However,this method of rice production may produce lower yields in a rice-wheat rotation system because of its poor seedling establishment.To address this problem,we performed field experiments to determine the rice yield at five seedling density levels (B1,B2,B3,B4,and B5=100,190,280,370,and 460 seedlings m-2,respectively) and clarify the physiological basis of yield formation.We selected a representative high-quality rice variety and a multifunctional seeder that used in a typical rice-wheat rotation area in 2016 and 2018.The proportion of main stem panicle increased with increasing seedling density.There was a parabolic relationship between yield and seedling density,and the maximum yield (9.34-9.47 t ha-1) was obtained under B3.The maximum yield was associated with a higher total spikelet number m-2 and greater biomass accumulation from heading to maturity.The higher total spikelet number m-2 under B3 was attributed to an increase in panicle number m-2 compared with B1 and B2.Although the panicle numbers also increased under B4 and B5,these increases were insufficient to compensate for the reduced spikelet numbers per panicle.Lower biomass,smaller leaf area,and lower N uptake per plant from the stem elongation stage to the heading stage were partially responsible for the smaller panicle size at higher seedling density levels such as B5.The higher biomass accumulation under B3 was ascribed to the increases in the photosynthetic rate of the top three leaves m-2 of land,crop growth rate,net assimilation rate,and leaf area index.Furthermore,the B3 rice population was marked by a higher grain-leaf ratio,as well as a lower export ratio and transport ratio of biomass per stem-sheath.A quadratic function predicted that 260-290 seedlings m-2 is the optimum seedling density for achieving maximum yield.Together,these results suggested that appropriately increasing the seedling density,and thereby increasing the proportion of panicles formed by the main stem,is an effective approach for obtaining a higher yield in DDR sown using a multifunctional seeder in a rice-wheat rotation system.
Keywords: seedling density,dry direct-seeded rice sown by a multifunctional seeder,rice-wheat rotation system,yield,biomass,photosynthetic capacity
1.Introduction
Dry direct-seeded rice (DDR) is a highly efficient,inexpensive,and environmentally friendly approach for rice establishment that has the potential to alleviate certain rice production problems such as a lack of labor(Chakrabortyet al.2017;Shiet al.2021),limited water resources,and excess greenhouse gas emissions (Burneyet al.2010).DDR requires less labor,time and cost than transplanted rice because some processes such as seedling production,pulling and transplantation can be skipped (Farooqet al.2011;Mahajanet al.2013;Lainget al.2018).Compared with transplanted and wet directseeded rice,DDR consumes less water because it uses dry ploughing,dry harrowing and dry sowing (Liet al.2014;Liuet al.2015).Finally,less methane is emitted with DDR because the lower soil moisture suppresses the growth of soil methanogens and supports the growth of soil methanotrophs relative to transplanted and wet directseeded rice (Hanget al.2014).In the past,DDR was sown after ploughing and harrowing (Kumar and Ladha 2011).In recent years,with the development of multifunctional seeders that perform synchronous rotary tillage and sowing(Wanget al.2017;Zhanget al.2017),DDR sown by multifunctional seeders has developed rapidly due to its labor-and time-saving features,and its planting area has expanded gradually throughout Asia,including China and India (Farooqet al.2011;Kumar and Ladha 2011;Wanget al.2017;Devkotaet al.2020).
The rice-wheat rotation system is one of the largest agricultural production systems in the world,and it occupies 24 million ha of the most productive land in South Asia and China (Ladhaet al.2003).Under the rice-wheat rotation system,DDR cannot be sown until the wheat has been harvested,and this limits DDR yields by compressing the growing season relative to that of transplanted rice(Xinget al.2017;Huang and Zou 2018).Moreover,the return of wheat-straw is widely used in this rotation system as an important agricultural measure to improve the climate,ambient air quality,and human health relative to the burning of wheat-straw in open fields (Grover and Chaudhry 2019).Wheat-straw return also ameliorates the soil structure,improves the soil organic matter content,and promotes sustainable soil development (Jianget al.2012;Xuet al.2018).Although wheat-straw return has many advantages,it is difficult to carry out high-quality mechanical tillage in large production areas for DDR sown by a multifunctional seeder that performs synchronous rotary tillage and sowing.This limitation is due to the large amounts of wheat-straw returned to the field,the low degree of chopped wheat-straw,and the uneven distribution of wheat-straw (Jianget al.2012).As a result,rice seed germination is constrained by less-than-ideal soil conditions.DDR seedlings also exhibit poor establishment because the returned wheat-straw causes microbial consumption of available soil nutrients (Zhaoet al.2017;Li Het al.2018;Nadeem and Farooq 2019),an accumulation of microbial allelochemicals such as organic acids (Qiet al.2012) and the restriction of rice seedling root development(Xuet al.2018;Wanget al.2019).Together,these production problems cause poor seedling establishment and yield loss in DDR sown by a multifunctional seeder under the rice-wheat rotation system,and rice grown in this manner has been termed “constrained direct-seeded rice”by Zhanget al.(2021).In this study,we call it constrained dry direct-seeded rice (CDDR).
Higher sowing rates present a problem during largescale production of CDDR.Higher seedling densities can partially compensate for yield loss through higher panicle numbers,but the biomass per plant and spikelet number per panicle decrease due to the effects of competition among individual plants.In addition,higher seedling densities can also aggravate the risk of weak seedlings,seedling heterogeneity,lodging,diseases,and pests(Meulen and Chauhan 2017).Nonetheless,there are also problems associated with lower sowing rates (Qiuet al.2016).Lower seedling densities are beneficial to the growth of individual plants,the accumulation of biomass per plant,and the formation of larger panicles,but it is difficult to improve the yields at low seedling densities because of insufficient panicle numbers.Therefore,determining the optimum seedling density is the most basic and important consideration for achieving higher yields in rice cultivation.Research on transplanted rice has shown that yield generally increases with seedling densities within the range of 5 to 100 seedlings m-2,and the yield remains stable through the balancing of panicle number and spikelet number per panicle at densities greater than 100 seedlings m-2(Fukushimaet al.2011;Clergetet al.2016;Huet al.2020).Previous studies have reported that 100-200 seedlings m-2was a practical optimal plant population (semi-dwarf rice variety) for both aerial-and drill-sowing methods in south-eastern Australia(Dunnet al.2020),70-118 seedlings m-2was a suitable density (hybrid rice variety) for wet direct-seeded rice in South China (Wanget al.2014),240 seeds m-2was found to be an appropriate density (inbred rice variety)for DDR in Central China (Sunet al.2015),and 160-220 seedlings m-2was a suitable density (inbredjaponicarice variety) for direct-seeded rice in rice-wheat rotation systems (Huanget al.2012;Chenet al.2015;Wuet al.2019).Some studies have shown that higher densities of 340-370 seedlings m-2produce good yields and minimize the yield loss caused by weeds (Phuonget al.2005).These previous studies were mainly carried out in soil after intensive tillage,such as puddled soil and soil after multiple rotary tillage,which means that all aspects of the soil tillage conditions were better,and this would be conducive to the growth of rice.However,the less-thanideal soil conditions caused by wheat-straw return and minimum tillage has become a common phenomenon in the large areas of CDDR production.Therefore,the results for optimal seedling density obtained using different systems may not be applicable to CDDR under a rice-wheat rotation system because this kind of rice experiences yield loss associated with poor seedling establishment.The optimum seedling density of CDDR should be re-evaluated for updating the current agronomic management practices in the rice-wheat rotation system.
Here,we performed an experiment in a typical ricewheat rotation area using a widely grown high-qualityjaponicarice variety and a representative multifunctional seeder,and measured relevant indexes of biomass and photosynthetic capacity from the levels of population to individual plants.The objectives of this study were to (1)determine the optimum seedling density for CDDR and(2) clarify the mechanism of the maximized yield at the optimum seedling density.The findings of this study provide theoretical and practical support for high-yield cultivation and regulatory approaches for CDDR in the future.
2.Materials and methods
2.1.Experimental site and weather conditions
The field experiment was performed in Taizhou City,Jiangsu Province,China (32°61´N,120°12´E,3 m altitude)during the rice growing season (from June to October).The cropping system in this region is a rice-wheat rotation system.The field soil was a sandy loam with a viscous texture and 30.36 g kg-1organic matter,1.91 g kg-1total N,31.57 mg kg-1available P,and 153.8 mg kg-1available K.
Meteorological data were collected at a weather station near the experimental site.The values for the daily mean temperature,sunshine hours,and precipitation during the two rice growing seasons were 25.58 and 25.54°C,679.4 and 857.7 h,and 1 083.9 and 485.5 mm in 2016 and 2018,respectively (Fig.1).
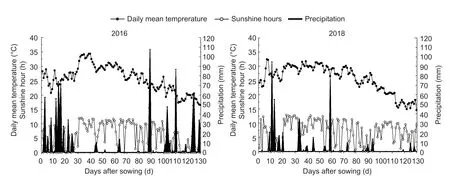
Fig.1 Daily mean temperature,sunshine hours and precipitation during the rice growth seasons in 2016 and 2018.
2.2.Rice variety and multifunctional seeder
The high-qualityjaponicarice variety Nanjing 9108 was selected for this experiment.In the planting area of conventionaljaponicarice,Nanjing 9108 was the fourthranked variety in China and the second in Jiangsu Province (CRDC 2021).A multifunctional seeder with synchronous rotary tillage and sowing (row spacing 25 cm and sowing depth 2-3 cm) was used in this experiment.The multifunctional seeder was developed by the College of Mechanical Engineering,Yangzhou University.This seeder is representative of the seeders used in largescale production to perform the one-time operation of rice sowing and improve sowing efficiency.
2.3.Experimental design and treatments
The experiment used a completely randomized design with three replications.Five target seedling density levels were used in 2016 and 2018: 100 seedlings m-2(B1),190 seedlings m-2(B2),280 seedlings m-2(B3),370 seedlings m-2(B4),and 460 seedlings m-2(B5).The sowing date in both years was 20 June.The seedling density levels were achieved by regulating the sowing rate based on the germination rate and the seedling emergence rate measured in a preliminary experiment.The actual seedling density achieved in each treatment was determined during the three-leaf stage (about 2 wk after sowing),and three representative plot areas (5 m×6 m)were selected based on the actual seedling density for each treatment.
2.4.Crop cultivation
In both years,wheat was harvested with Kubota machinery (4LBZ-145G(PRO588I-G)),and 100% of the wheat-straw was returned to the field.Next,the rice for each treatment was seeded with the multifunctional seeder.After sowing,wet irrigation management was used during the seedling period to ensure the growth and development of the rice seedlings.Thereafter,moist conditions were maintained until the 5-leaf stage.The field was flooded after the 5-leaf stage,and the water level was maintained at 2-3 cm until the middle tillering stage.The water was then drained for 7-10 d to control unproductive tillers.After the stem elongation stage,a regime of alternating wetting and moderate soil drying irrigation management was used until 1 wk before the final harvest.In this alternating wetting and moderate soil drying irrigation management regime,the fields were not irrigated until the soil water potential reached-15 kPa (soil moisture content 0.170 g g-1) at 15-20 cm depth.The soil water potential was monitored at 15-20 cm soil depth with a tensiometer consisting of a sensor of 5 cm length (Institute of Soil Science,Chinese Academy of Sciences,Nanjing,Jiangsu,China).Four tensiometers were installed in each plot,and readings were recorded at 11:00 a.m.each day.When the soil water potential reached the threshold,plots were flooded with 3 cm of water.The irrigation water level was monitored with a flow meter (LXSG-50 flow meter,Shanghai Water Meter Manufacturing Factory,Shanghai,China) installed in the irrigation pipelines.
A total of 270 kg N ha-1(as urea with an N content of 45.6%) was applied in three splits at a ratio of 3.5:3.5:3 at pre-sowing,the 4-leaf stage,and the stem elongation stage.Phosphorus (135 kg P ha-1) was applied at presowing using calcium superphosphate with a P2O5content of 12%.Potassium chloride (K2O content: 60%) was used as the potassium fertilizer,and it was applied at presowing (135 kg K ha-1) and again at the panicle initiation stage (135 kg K ha-1).
Weed,insect,and disease control followed local recommendations throughout the growing season to minimize yield losses over the two years.
2.5.Sampling and measurements
To accurately sample and understand the growth process of the rice,we recorded the dates of sowing,the stem elongation stage (panicle initiation),the heading stage,and the maturity stage.In 2018,tiller numbers were measured in each plot from three adjacent and consecutive rows (1 m length) at 5-day intervals from the 3-leaf stage until the number diminished.
Tiller number (Y) was fit with a logistic equation:

whereYmis the maximum tiller number,xis days after sowing,andAandBare rate-controlling parameters(Catonet al.2003).The fitting was performed using Origin 9.
Leaf area and biomass were determined at the stem elongation,heading,and maturity stages.Samples were collected from three adjacent rows (50 cm length) in each plot,and all samples were separated into leaves,stem-sheaths,and panicles (at the heading and maturity stages).Leaf areas were measured with a leaf area meter (LI-3100,LI-COR,USA).Each component of the rice plants was oven-dried separately at 105°C for 30 min and then in bags at 80°C to a constant weight.N uptake per plant was calculated by multiplying the N concentration per plant (%) by the biomass per plant,and N concentration was determined by the semi-micro-Kjeldahl procedure.
Five representative plants were selected and marked in each plot forin situmeasurement of the photosynthetic rate.Photosynthetic rates of the top first,second,and third leaves were measured at the heading stage and 20 and 40 days after heading.The maximum CO2assimilation rate m-2(i.e.,the photosynthetic rate;μmol m-2s-1) was measured between 9:30 and 11:30 a.m.using an LI-6400 portable photosynthetic system (LICOR,USA).At the same time,10 representative plants were selected for taking measurements of the leaf areas of the top first leaf to the third leaf (LI-3100,LI-COR,USA).
At the maturity stage,three adjacent rows (1 m length) were sampled randomly from each plot for the measurements of panicle traits and yield components,including spikelet number per panicle (SP),filled-grain percentage,and 1 000-grain weight.Three adjacent rows (5 m length) were sampled randomly from each plot for determination of the panicle number m-2(PN).Total spikelet number m-2(TSP) was calculated as PN multiplied by SP.Yield was determined from a harvest area of 8 m2in each plot and adjusted to the standard moisture content of 0.14 g H2O g-1.
2.6.Statistical analysis and formula calculations
The data were analyzed using IBM SPSS Statistics 22,and treatment means were compared by the least significant difference (LSD) test.Graphical representations of the data were produced using Origin 9,Excel 2019,and the R statistical programming language.
The following equations were used for the calculations:
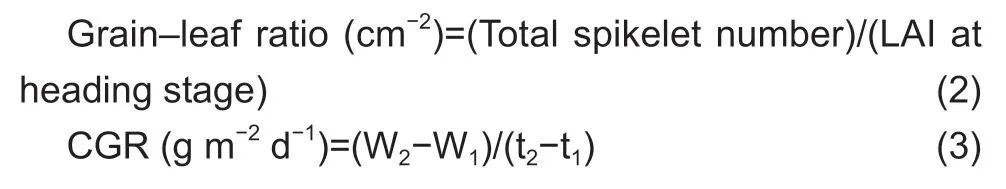
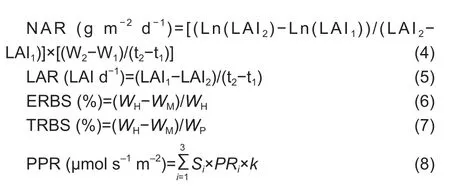
where CGR is the crop growth rate,NAR is the net assimilation rate,LAR is the leaf area attenuation rate from heading to maturity,ERBS is the export ratio of biomass per stem-sheath from heading to maturity,TRBS is the transport ratio of biomass per stem-sheath from heading to maturity,and PPR is the photosynthetic rate of the top three leaves m-2of land.W1and W2are the first and second measurements of biomass (kg ha-1),t1and t2are the first and second measurement times (d),LAI1and LAI2are the first and second measurements of leaf area index (m2m-2),Ln indicates the natural logarithm,WHandWMare the stem-sheath weights (after removal of spikes and leaves) at heading and maturity (g),WPis the single panicle weight at maturity (g),Siis the leaf area of theith leaf (from the top) per plant (m2),PRiis the photosynthetic rate of theith leaf per plant (μmol m-2s-1),andkis the number ofith leaves per m2of land (m-2) after heading.
3.Results
3.1.Yield and seedling density
Data concerning the yields and their components in 2016 and 2018 are shown in Table 1.The B3 density level (280 seedlings m-2) produced the highest yield of 9.34-9.47 t ha-1,which was 6.9-7.5% higher than B1 and 8.1-9.4%higher than B5.The relationship between seedling densityand yield was analyzed with a scatter plot (Fig.2),and in all cases,a parabolic relationship fit the data well.The relationships between grain yield (Y) (kg ha-1) and seedling density (X) (m-2) wereY2016=-0.0292X2+15.5X+7 175.1(R2=0.919) andY2018=-0.0291X2+16.4X+7 059.4 (R2=0.956)in 2016 and 2018,respectively.Based on these quadratic equations,the optimum seedling densities to achieve higher yield were 265 m-2in 2016 and 282 m-2in 2018.
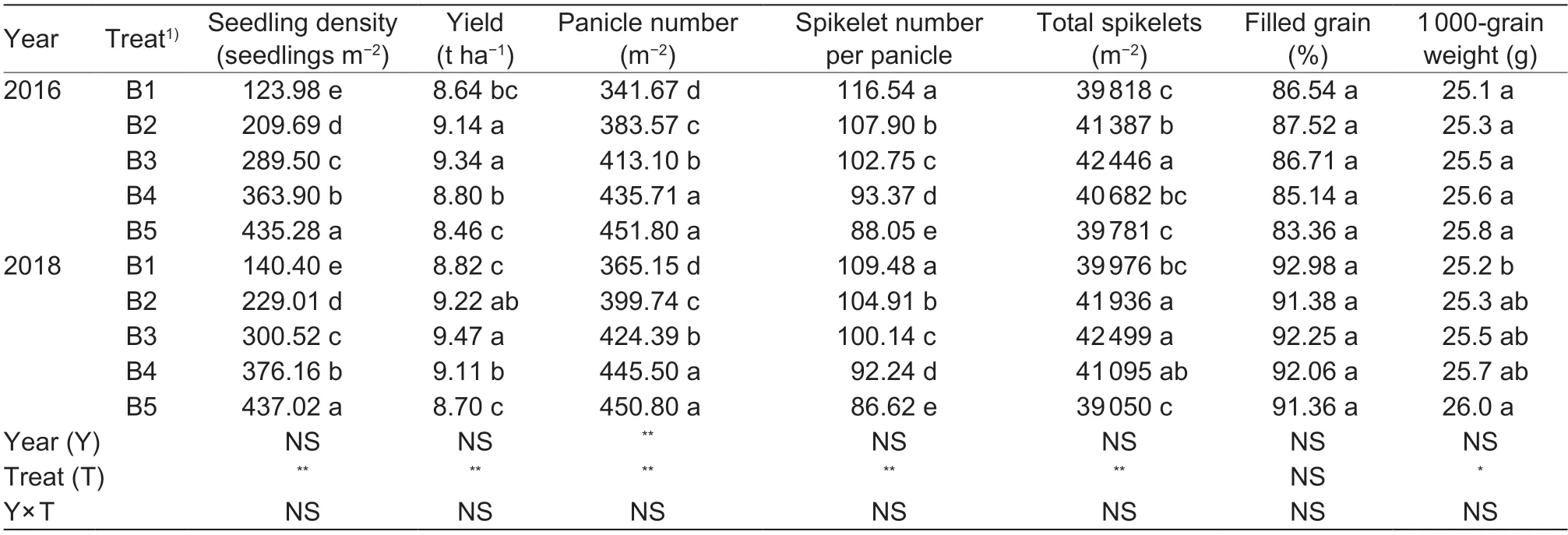
Table 1 Yield and yield components under five seedling density levels in 2016 and 2018
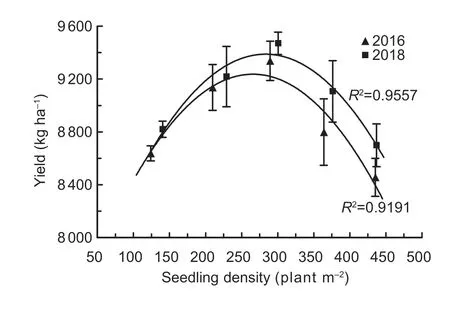
Fig.2 The relationships between seedling density and yield in 2016 and 2018.Error bars denote standard error of replicates(n=3).The actual seedling densities are shown in Table 1.
The yield components were analyzed,and consistent differences were observed in the two years (Table 1).As seedling density increased,panicle number (PN) and the proportion of panicles formed by the main stem also increased.The latter values were 37.4,56.0,70.4,84.0,and 96.4% in B1,B2,B3,B4,and B5,respectively.In contrast,spikelet number per panicle (SP) decreased significantly as seedling density increased.For example,B5 had 21.7% higher PN than B1,but it also had 22.7%lower SP than B1;as a consequence,total spikelet number (TSP) was 1.2% higher in B5 than in B1.Meanwhile,B3 had 15.7% higher PN than B1 but only 10.4% lower SP,and TSP was 6.0% higher in B3 than in B1.Although B3 had 7.2% lower PN than B5,it had 13.9% higher SP,and TSP was therefore 7.0% higher in B3 than in B5.Seedling density had a positive effect on 1 000-grain weight,but this effect was not significant.Seedling density did not have a significant effect on filledgrain percentage.
The correlation between yield and seedling density was not significant (Fig.3),nor were there significant correlations between yield and the yield components including PN,SP,filled-grain percentage,and 1 000-grain weight.However,there was a significant positive correlation between yield and TSP.Seedling density was significantly positively correlated with PN and 1 000-grain weight,and significantly negatively correlated with SP.There were significant negative correlations of 1 000-grain weight with SP,grain density,and panicle length.

Fig.3 Correlation analysis among seedling density,yield components and panicle traits (R0.05=0.878 and R0.01=0.959).SD,seedling density;PN,panicle number m-2;SP,spikelet number per panicle;TSP,the total spikelet number m-2;FG,filled-grain percentage;TGW,1 000-grain weight;GD,grain density (spikelets per panicle/PL);PL,panicle length;GWP,grain weight per panicle.
3.2.Tillering dynamics
The peak value of tillers m-2increased as seedling density increased,and the value in B5 was 171.3-172.8% and 43.3-51.4% higher than in B1 and B3,respectively.Moreover,the time to achieve maximum tillers m-2was also reduced,and it occurred about 5-10 days earlier in B5 than in B1 (Fig.4,2018).However,after the peak tiller value,it is clear from the slope of the tiller curve that the tiller number declined more rapidly at higher seedling densities (Fig.4,2016).
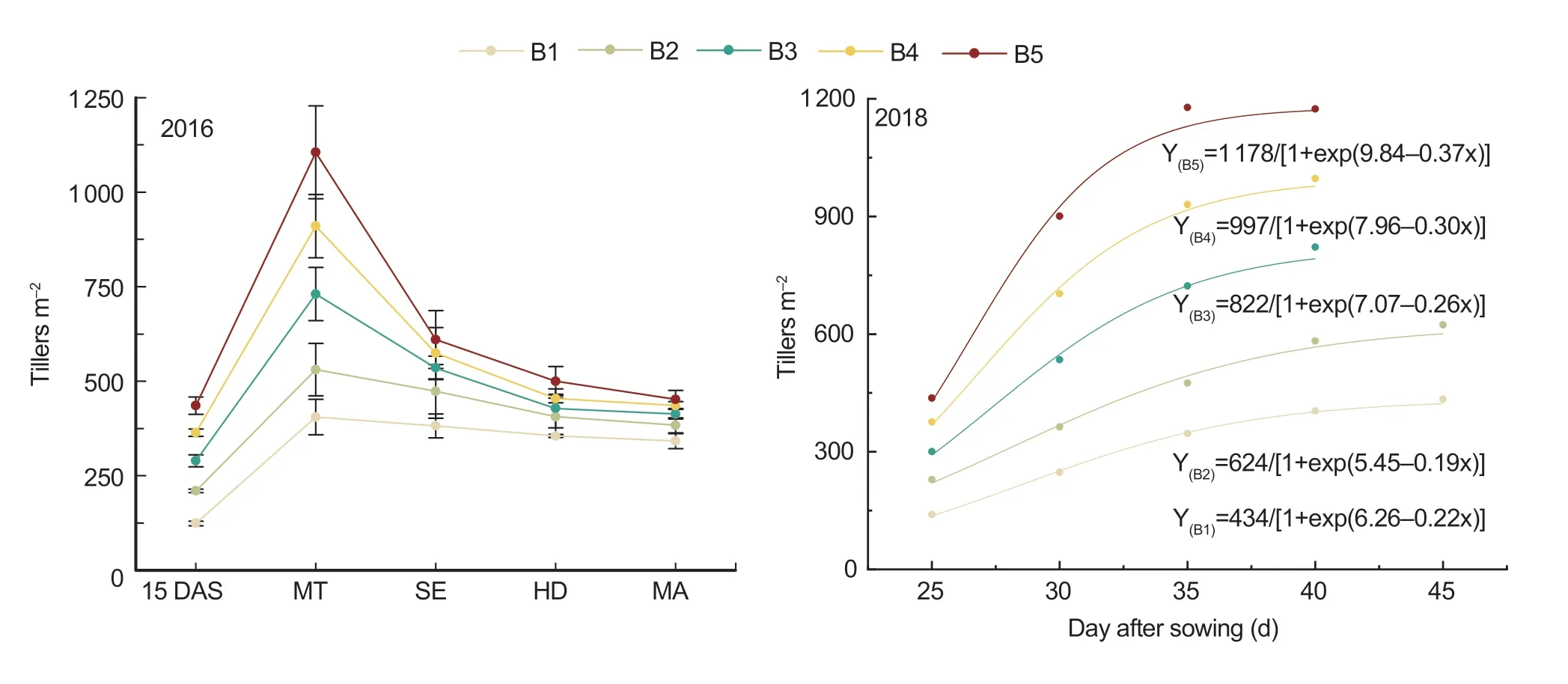
Fig.4 Tillering dynamics under five seedling density levels during the growth duration in 2016 and the early growth period in 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).15 DAS,15 days after sowing;MT,middle tillering stage;SE,stem elongation stage;HD,heading stage;MA,maturity stage.Error bars denote standard error of replicates (n=3).
3.3.Leaf area and N uptake per plant
Leaf area and N uptake per plant decreased with increasing seedling density in all growth periods (Fig.5).Leaf area per plant in B1 was 15.4-17.7% and 15.8-24.1%higher than in B3 and B5 at the stem elongation stage,11.0-11.6% and 13.0-23.0% higher than in B3 and B5 at heading,and 7.9-11.5% and 17.8-23.2% higher than in B3 and B5 at maturity,respectively.Likewise,N uptake per plant in B1 was 10.8-12.7% and 19.2-21.2% higher than in B3 and B5 at the stem elongation stage,and 17.4-20.7%and 20.7-25.1% higher than in B3 and B5 at heading,respectively.
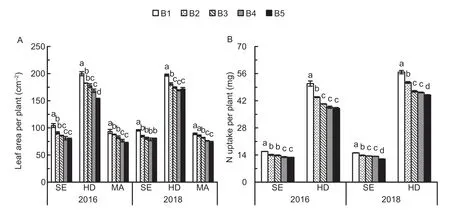
Fig.5 Leaf area per plant (A) at the stem elongation (SE),heading (HD),and maturity (MA) stages and N uptake per plant (B) at the SE and HD under the five seedling density levels in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
3.4.LAI and LAR
As shown in Fig.6,there were differences in LAI among the five seedling densities at the stem elongation,heading,and maturity stages.The LAI in B5 was 17.4-17.5%and 7.2-7.8% higher than in B1 at the stem elongation and heading stages,respectively.However,there were negligible differences in LAI between the higher seedling densities such as B3 and B5 at the heading stage.The LAI in B3 at maturity was 6.6 and 5.2% higher than in B1 and B5,respectively.The LAR increased with increasing seedling density from heading to maturity,and it was 11.4% higher in B5 than in B1.
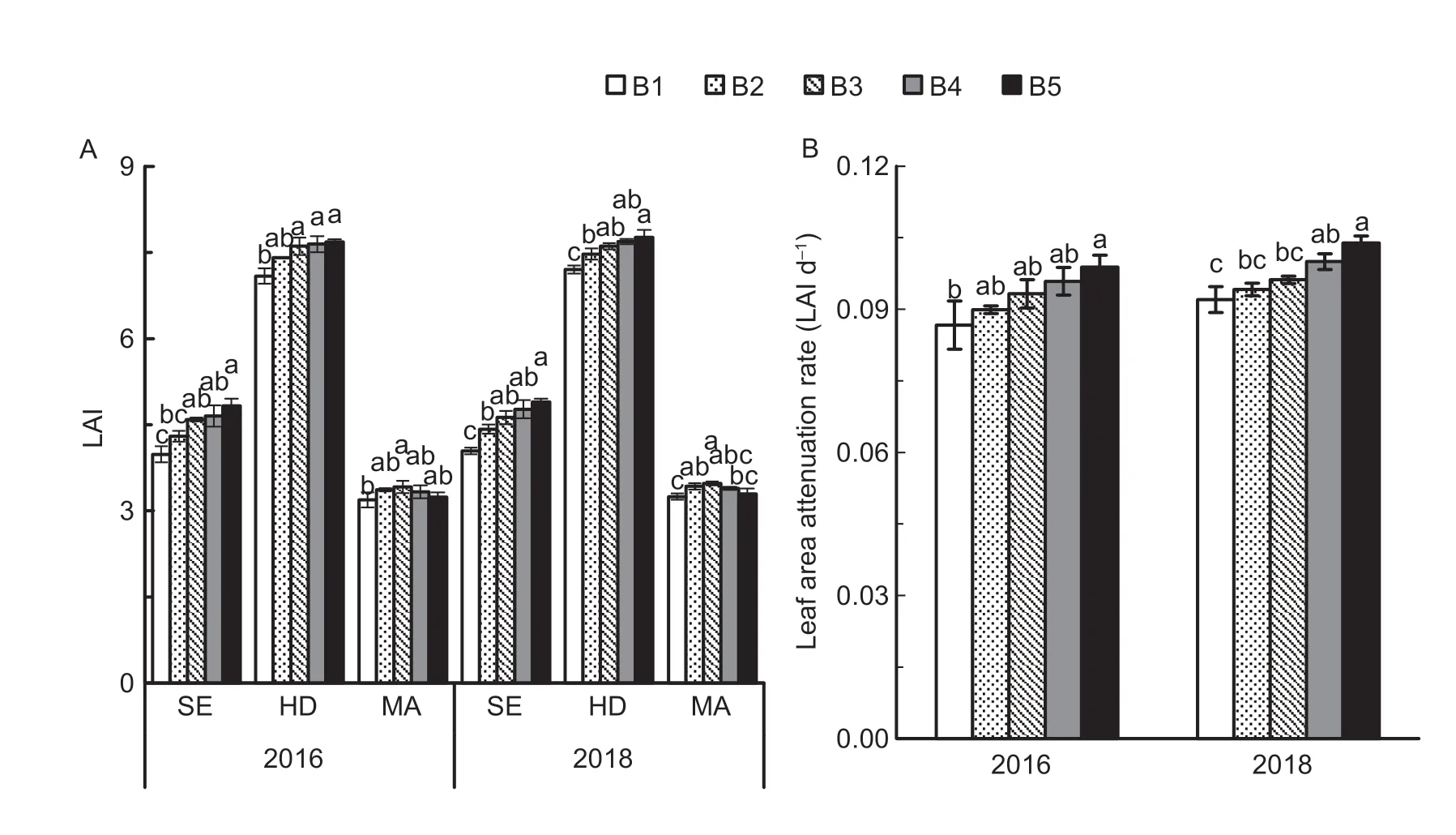
Fig.6 Leaf area index (LAI) (A) at the stem elongation (SE),heading (HD),and maturity stages (MA);and leaf area attenuation rate (B) from HD to MA under five seedling density levels in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
3.5.Biomass,CGR and NAR
As seedling density increased,biomass accumulation increased at the stem elongation and heading stages.Specifically,the biomass in B5 was 9.2-15.4% and 6.6-6.7% higher than in B1 at the stem elongation and heading stages,respectively.However,there was a parabolic relationship between seedling density and biomass accumulation from heading to maturity,and biomass in B3 was 8.0-19.5% and 13.3-20.4% higher than in B1 and B5,respectively.Finally,biomass accumulation in B3 at maturity was 5.7-11.3% and 3.2-7.4% higher than in B1 and B5,respectively.Contrary to the trend in biomass accumulation,biomass per plant in B1 was 23.5-26.0%,13.6-23.9% and 16.8-21.1% higher than in B5 at the stem elongation,heading,and maturity stages,respectively (Fig.7-B).

Fig.7 Biomass accumulation (A) at sowing stage (SO)-stem elongation (SE),SE-heading (HD) and HD-maturity (MA) stages and biomass per plant (B) at SE,HD and MA under the five seedling density levels in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
The crop growth rate and net assimilation rate from heading to maturity initially increased but later decreased with increasing seedling density (Fig.8).In the two years,the crop growth rate in B3 from heading to maturity was 13.7 and 16.9% higher than in B1 and B5,respectively.Similarly,the net assimilation rate in B3 from heading to maturity was 7.9 and 15.7% higher than in B1 and B5,respectively.
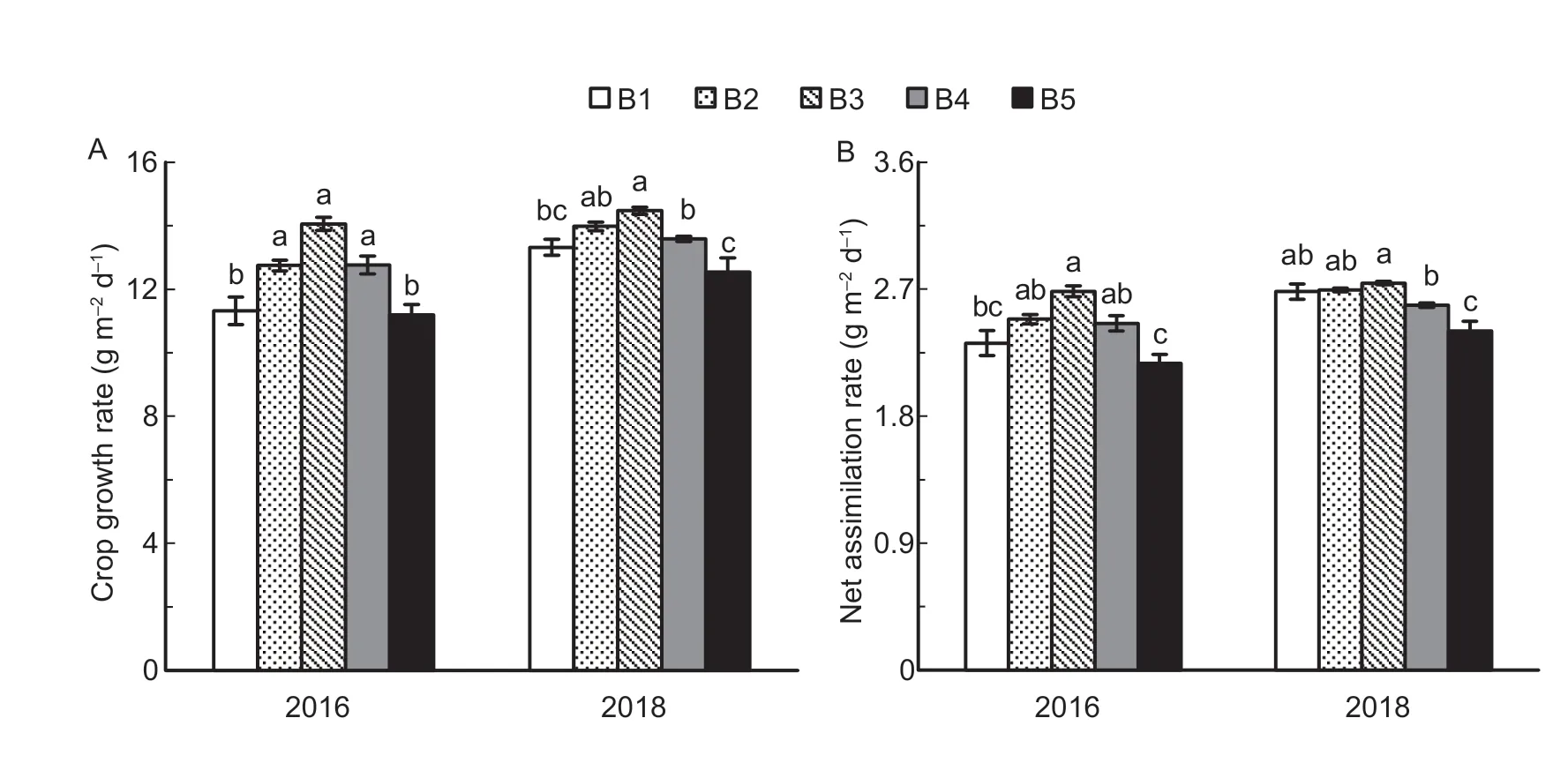
Fig.8 Crop growth rate (A) and net assimilation rate (B) under the five seedling density levels from the heading to the maturity stages in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
3.6.Photosynthetic rate
At heading,20 and 40 days after heading,the photosynthetic rates of the top three leaves decreased consistently with increasing seedling density,but the rates did not differ significantly among seedling density levels of B1 to B4 (Fig.9).The photosynthetic rates of the top three leaves were ranked the top first leaf>top second leaf>top third leaf.From heading to 40 days after heading,the photosynthetic rates of the top first,second,and third leaves in B1 were approximately 33.1,30.7 and 45.3% higher than in B5,respectively.

Fig.9 Photosynthesis rates of the top three leaves at DAH-0 d,DAH-20 d and DAH-40 d under the five seedling density levels in 2018.DAH-0 d,DAH-20 d and DAH-40 d represent 0,20 and 40 days after the heading stage (DAH),respectively.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
The photosynthetic rates of the top three leaves m-2of land (PPR) after heading increased and then decreased with increasing seedling density (Fig.10-A),and PPR was significantly lower at higher seedling density levels such as B5.PPR values in B3 were 3.0,9.1 and 1.1% higher than in B1 at 0,20 and 40 days after heading,respectively.Likewise,PPR levels in B3 were 34.2,29.9 and 27.2% higher than in B5 at 0,20 and 40 days after heading.The mean PPR in B3 across 0,20 and 40 days after heading (PPR at HD-MA) was 5.9 and 30.5% higher in B1 and B5.The scatter plot(Fig.10-B) showed that there was a significant positive correlation between biomass accumulation at HD-MA and PPR at HD-MA.
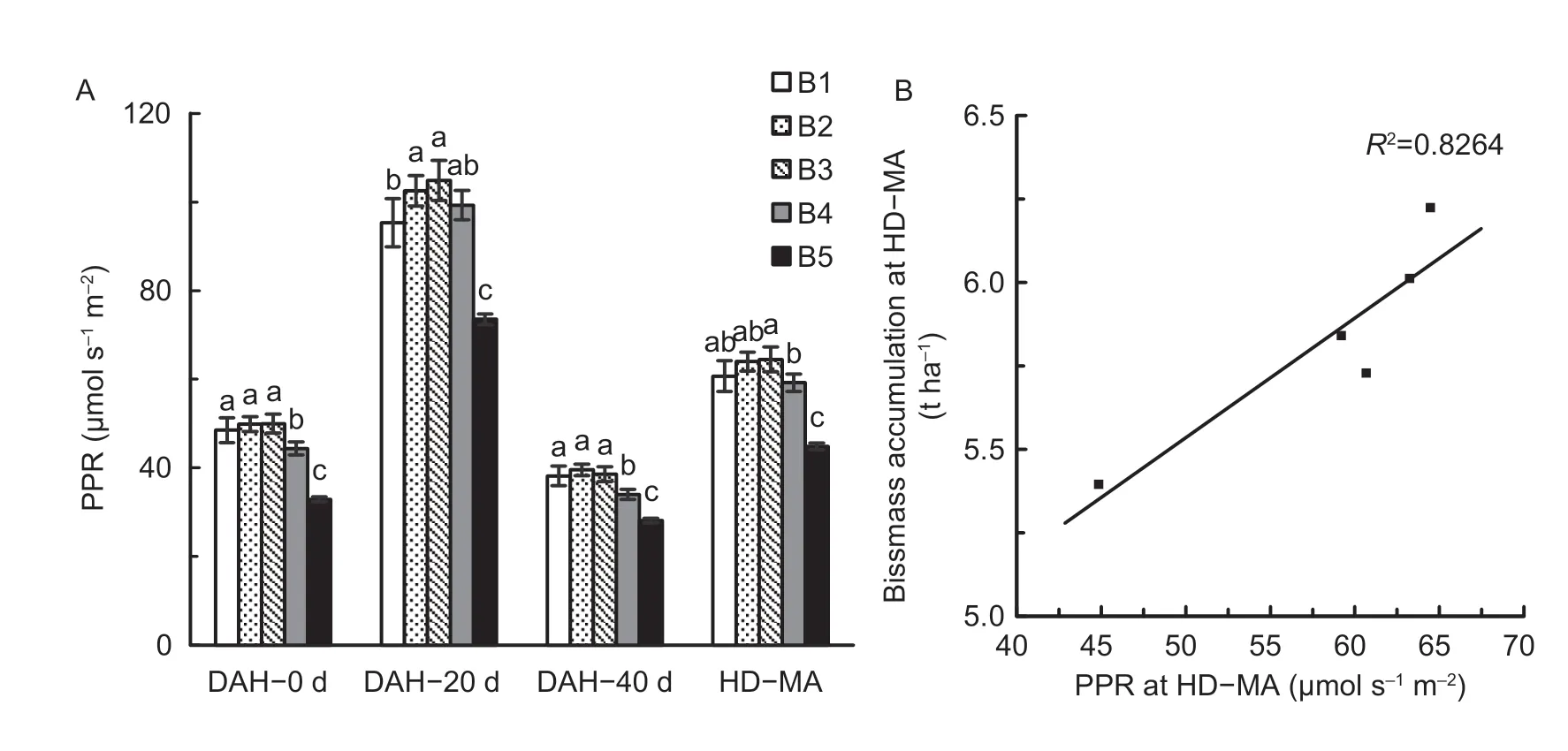
Fig.10 Photosynthetic rates (PPR) of the top three leaves m-2 of land (PPR) at DAH-0 d,DAH-20 d,DAH-40 d,and HD-MA (A)and the relationship between biomass accumulation and PPR at HD-MA (B) under the five seedling density levels in 2018.DAH-0 d,DAH-20 d and DAH-40 d represent 0,20 and 40 days after the heading stage (DAH),respectively.HD-MA,from heading to maturity stage.PPR at HD-MA represents the average values of PPR at DAH-0 d,DAH-20 d and DAH-40 d.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
3.7.Source and sink
The grain-leaf ratio decreased as seedling density increased,but there were no significant differences among B1,B2 and B3.The grain-leaf ratio decreased significantly at higher seedling density levels such as B4 and B5.The grain-leaf ratio in B1 was 7.9-9.4% higher than in B5 (Fig.11).
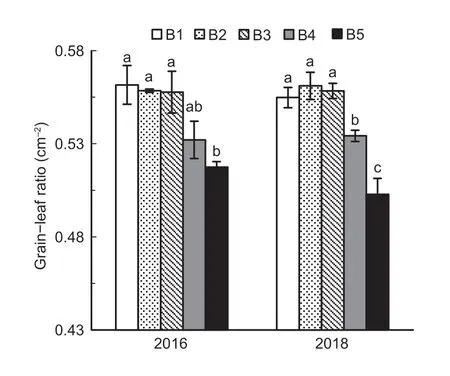
Fig.11 Grain-leaf ratio at the heading stage under the five seedling density levels in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
As shown in Fig.12,the export ratio of biomass per stem-sheath (ERBS) and the transport ratio of biomass per stem-sheath (TRBS) differed among seedling densities.There were parabolic relationships between seedling density and both ERBS and TRBS from heading to maturity: ERBS and TRBS first decreased and then increased with increasing seedling density.In both years,ERBS in B3 was 12.4-33.0% and 22.4-25.3% lower than in B1 and B5.Similarly,TRBS in B3 was 8.9-28.0% and 30.7-31.1% lower than in B1 and B5.
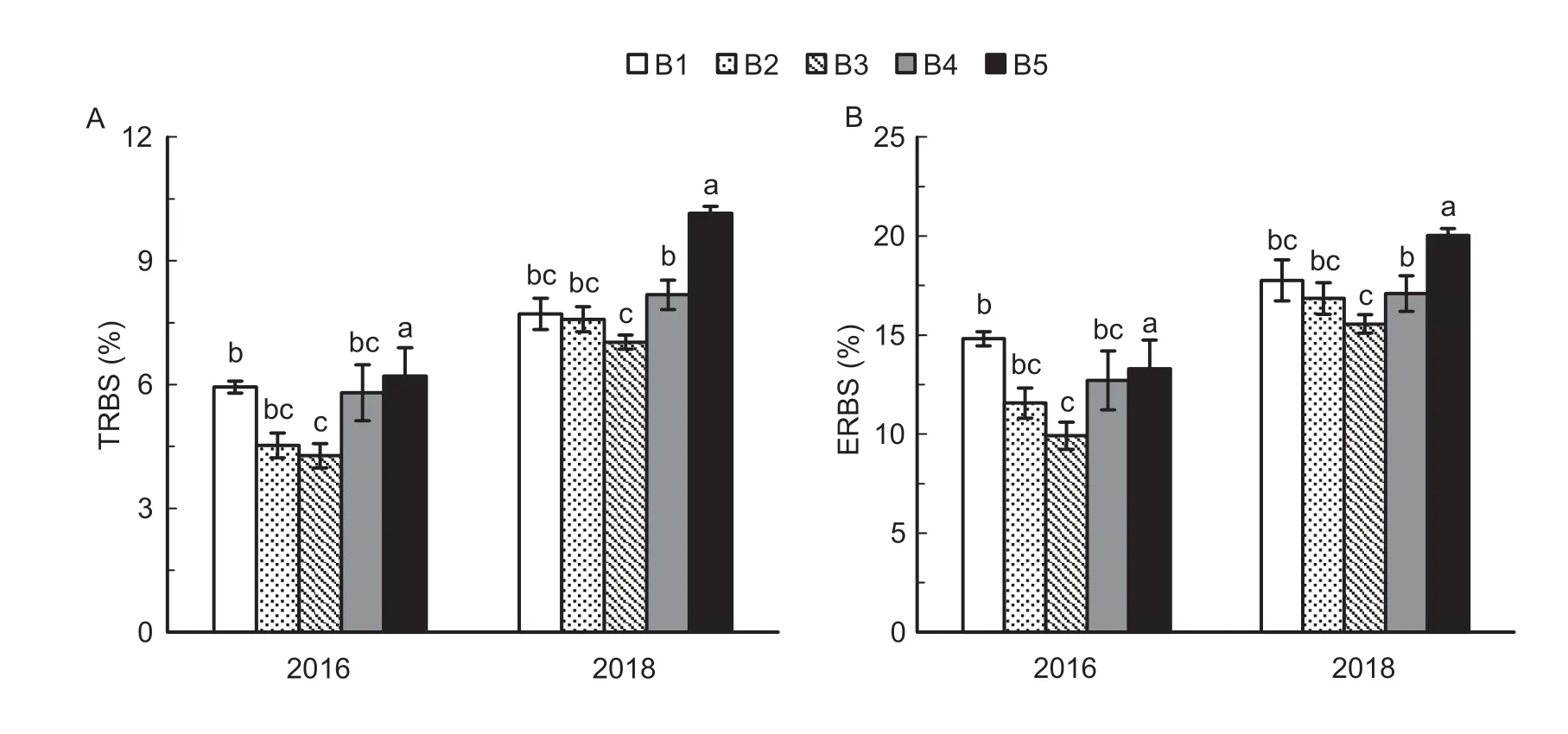
Fig.12 Transport ratio of biomass per stem-sheath (TRBS) (A) and export ratio of biomass per stem-sheath (ERBS) (B) from heading to the maturity stage under the five seedling density levels in 2016 and 2018.B1,B2,B3,B4,and B5 represent target seedling density levels of 100,190,280,370,and 460 seedlings m-2,respectively (the actual seedling densities are shown in Table 1).Error bars show standard error of replicates (n=3).Values followed by different lowercase letters are significantly different at the 0.05 probability level among the different seedling density treatments.
4.Discussion
4.1.Yield responses to seedling density
Seedling density is a critical factor for establishing an appropriate crop population,thereby ensuring high quality and yield (Huanget al.2013;Liet al.2016).Here,we found that yield first increased and then decreased with increasing seedling density,and the maximum yield was obtained at a seedling density level of 280 seedlings m-2(B3).Although some studies have shown that seedling density (sowing rate) has no effect on yield (Ottis and Talbert 2005),sink size,or gross hulled rice yield(Fukushimaet al.2011),they also reported a peak value of yield with increased seedling density.The response curve between yield and seedling density was fitted well by a quadratic function,and the quadratic equation obtained from the two-year experiment predicted that 260-290 seedlings m-2is the optimum seedling density for maximum yield.Previous studies on seedling density of inbredjaponicarice showed that 80-120 seedlings m-2for transplanted rice (Huet al.2020) and 160-220 seedlings m-2for wet direct-seeded rice (Chenet al.2015;Wuet al.2019) were suitable densities in the ricewheat rotation system.However,in this experiment,these seedling density levels (B1 and B2) did not produce the maximum yield.One possible reason is that the poor seedling establishment caused by less-than-ideal soil conditions leads to insufficient tiller numbers and biomass accumulation.Ottis and Talbert (2005) recommended that growers plant twice as many seeds as needed for optimum rice density when planting in a clay soil or under imperfect seedbed conditions.Previously recommended seedling densities would have caused yield loss for CDDR under the conditions of this experiment,and previous seedling density studies are not directly applicable to CDDR in a rice-wheat rotation system.The results of this study preliminary suggest that higher yields for CDDR in the rice-wheat rotation system can be obtained by properly increasing the seedling density.To verify the feasibility of this cultivation model for CDDR,on-farm demonstration experiments were performed in rice-wheat rotation fields of Jiangyan and Yancheng cities,Jiangsu Province in 2020 with another widely used rice variety,Nanjing 5718.The yields reached 10.4 t ha-1in Jiangyan and 10.8 t ha-1in Yancheng.Therefore,appropriately increasing the seedling density is one of the approaches for achieving higher yields of CDDR in the rice-wheat rotation system.
This study showed that a type of homeostasis was maintained between PN and SP (Table 1).This was consistent with previous studies that reported a compensation effect between PN and SP: if one increased,the other declined (Yinget al.1998;Huanget al.2011).This phenomenon may be linked to plant density.On the one hand,plant density directly determines panicle density (PN) based solely on plant numbers.On the other hand,plant density can determine panicle size (SP) by affecting panicle development and spikelet formation.Differences in planting density change the environment around individual plants,including light quantity (Liet al.2019),canopy temperature,humidity,and carbon dioxide concentration,and these changes can affect gene expression (Lewiset al.2008;Endo-Higashi and Izawa 2011),hormone synthesis (Li Yet al.2018;Wanget al.2020) and growth (Clergetet al.2016)with respect to panicle and spikelet formation.There is also interplant competition for resources such as light and soil nitrogen,and increased competition at higher plant densities will lead to poorer development of individual plants.Thus,previous results have consistently shown positive correlations between SP and biomass (Shiratsuchiet al.2007),nitrogen status (Kamijiet al.2011),panicle neck diameter (Liuet al.2008),stem diameter,and vascular bundle (Xuet al.1998) per plant during panicle development (from stem elongation to heading).Sheehyet al.(2001) reported that source strength was particularly important for the differentiation and development of the sink organ,and Kobayashi and Horie (1994) believed that an increase in plant N significantly increased the number of differentiated spikelets at the panicle development stage by promoting the differentiation of secondary branches on primary branches.Here,we concluded that the smaller panicle size at higher seedling densities was caused by lower biomass (Fig.7-B),smaller leaf area(Fig.5-A),and lower nitrogen uptake (Fig.5-B)per plant from stem elongation to heading.
The coordination of panicle density and panicle size is necessary to optimize the homeostasis between PN and SP,and thereby achieve higher yields.The degree of coordination between panicle density and panicle size is reflected in TSP: the larger the TSP,the better the coordination and the higher the yield.We concluded that the smaller TSP at lower seedling density levels arose mainly from insufficient panicle numbers caused by insufficient tillers in CDDR in the rice-wheat rotation system (Fig.4).Although increasing the seedling density reduced the tiller number per plant,it increased the panicle density,thereby compensating for the yield loss caused by insufficient panicle numbers.The proportions of panicles formed by the main stem increased with increasing seedling density,at levels of 37.4,56.0,70.4,84.0,and 96.4% in B1,B2,B3,B4,and B5,respectively.Therefore,we preliminarily concluded that an effective means for increasing yield in CDDR is to appropriately increase the seedling density,thereby increasing the proportion of panicles formed by the main stem.In this way,a coordinated rice population with higher TSP and yield was produced through sufficient main stem panicle numbers,simultaneously ensuring a sufficient filled-grain percentage and 1 000-grain weight.
Grain weight is one of the yield components.The results of previous studies have varied regarding the effect of seedling density on grain weight.Yuet al.(1995)believed that reducing seedling density appropriately and establishing an appropriate population structure could effectively improve the plant physiological activity after flowering,increase grain storage capacity and enrich capacity,and increase grain weight.However,some studies have shown that the 1 000-grain weight increases with an increase in seedling density (Xuet al.2008;Huet al.2015).The results of this study showed that increasing seedling density was conducive to increasing 1 000-grain weight (Table 1).The grain weight of rice is closely related to grain filling.The grain filling process depends on two main carbon resources: photosynthetic assimilates and carbohydrates stored during pre-flowering and transported to the grain from the stem-sheath(Ambavaramet al.2014;Xuet al.2016).This study showed that although the ERBS and TRBS decreased(Fig.12),the increase of biomass accumulation after heading (Fig.7) could be conducive to the increase of grain weight at the seedling density levels from B1 to B3 (Table 1).Biomass accumulation after heading decreased at the seedling density levels from B3 to B5(Fig.7),however,the increases in ERBS and TRBS(Fig.12) also promoted the increase in grain weight(Table 1).Previous studies have shown that the grain weight of the main stem panicle is greater than that of the tiller panicle (Quet al.2009).Therefore,the increase in the proportion of main stem panicles could also be one of the reasons for the increase in the 1 000-grain weight with the increase in seedling density (Table 1).Furthermore,correlation analysis showed that the 1 000-grain weight was negatively correlated with grain density and spikelet number per panicle (Fig.3).Therefore,lower grain density and spikelet number per panicle could provide more growth space and material supply for single grain filling,which was also beneficial to the increase in grain weight.
4.2.Photosynthetic biomass productivity
Yield is the product of harvest index and biomass accumulation at the maturity stage.Given the current high yield levels,it is difficult to improve the harvest index of rice.Therefore,increasing the harvest index may no longer be the main approach for increasing rice yield,and improving biomass accumulation at the maturity stage may be the key to further improvements in rice yield potential based on a high and stable harvest index(Penget al.1999;Liet al.2009).In particular,increasing biomass accumulation from heading to maturity is critical for high biomass accumulation at maturity (Linget al.1993).However,a higher seedling density does not mean higher biomass accumulation.This study showed that biomass accumulation at maturity was the highest at density B3.Lower biomass accumulation at the seedling density levels recommended (B1 and B2) in previous studies (for transplanting or wet direct-seeded rice)was due to lower plant density (panicle density),which is caused by fewer tillers (Fig.4).The poor seedling establishment and the insufficient growth of the new tiller and main stem were partly responsible for the lack of tillers in B1 and B2.The main reason for the poor seedling establishment compared with wet direct-seeded rice may be the microbial consumption of soil available nutrients and the less-than-ideal soil conditions caused by wheat-straw return and minimum tillage.The main reason for the insufficient growth of the new tiller and main stem compared with transplanting rice may be that the seedling growth time in the vegetative growth period (from sowing to stem elongation) was seriously compressed (i.e.,shorter vegetative growth).For example,the reduced number of days from sowing to the stem elongation stage accounted for more than 60% of the shortened duration of the whole growth period (Xinget al.2017).Therefore,adequate main stems are needed to compensate for the insufficient new tillers for CDDR in the rice-wheat rotation system.Biomass accumulation at maturity decreased with increasing seedling density from B3 to B5,mainly owing to the decrease in biomass accumulation from heading to maturity.Although higher seedling densities improved biomass accumulation before heading,they were not conducive to biomass accumulation after heading because the individual rice plants were thin and weak.Hence,a suitable seedling density is necessary to improve biomass accumulation from heading to maturity.Further analysis showed that the crop growth rate and the net assimilation rate were the highest in B3,perhaps because of its higher LAI and lower LAR from heading to maturity (Fig.6).Therefore,the results of this study suggest that appropriately increasing the seedling density,and thereby increasing the proportion of panicles formed by the main stem,is beneficial for increasing the biomass accumulation of CDDR,especially the biomass accumulation after heading,by increasing the number of sufficient main stem panicles.
Higher biomass accumulation could be attributed to higher photosynthesis in B3,because the leaf photosynthetic rate plays a crucial role in determining biomass accumulation and yield.Previous research has demonstrated that the leaf photosynthetic rate is based on leaf N content and stomatal conductance (Kanemuraet al.2007;Ohsumiet al.2007).Rice populations with lower plant densities may have higher single-leaf photosynthetic rates owing to higher leaf N content and uptake (Fig.5).However,lower plant densities did not always equate to higher leaf photosynthetic rate m-2of land,because this parameter encompasses the single-leaf photosynthetic rate,single-leaf area,and number of leaves m-2of land(eq.(8)).The top three leaves of rice control about 75%of the yield,and this point was further confirmed through rice simulation models (Hiroeet al.2008;Hiroe and Takeshi 2009).Adachiet al.(2017) also reported that the top three leaves are the main photosynthetic organs in the later growth period.We therefore analyzed the leaf photosynthetic rate m-2of land based on the cumulative photosynthetic rate of the top three leaves m-2of land(PPR) (eq.(8)).There was a positive correlation between PPR and biomass accumulation from heading to maturity(Fig.10-B).The lower PPR at low seedling density levels,such as B1 and B2,was caused by a lower number of top three leaves m-2of land (plant density),although the low seedling densities were characterized by larger single-leaf photosynthetic rates and leaf areas per plant.The lower PPR at high seedling density levels,such as B4 and B5,was caused by lower photosynthetic rates of the top three leaves and smaller leaf areas of the top three leaves per plant,indicating that the increased plant densities could not compensate for the decreases in leaf area and single-leaf photosynthetic rate.The rice population of B3 exhibited a balance between plant density on one hand and single-leaf photosynthetic rate and leaf area on the other,ultimately improving PPR and achieving higher biomass accumulation and yield for CDDR.Therefore,we preliminarily concluded that appropriately increasing the seedling density,and thereby increasing the proportion of panicles formed by the main stem,is beneficial for improving the photosynthetic capacity of the population,which is beneficial for increasing the biomass accumulation after heading in CDDR.
In this study,the leaf photosynthetic rate m-2of land was determined by measurement and calculation.Although this approach differs from previous methods(Garrityet al.1984;Zhaoet al.2001),it can describe the relationships of both yield and biomass accumulation with leaf photosynthetic rate m-2of land,and it therefore has the value for the study of population photosynthetic productivity.
The formation of crop yield is a process of sourceflow-sink interaction and is related to the production,transport,and accumulation of photosynthate.Previous studies have shown that the sink organ determines the leaf photosynthetic rate,as reductions in the carbon demand of sink tissues eventually limit the photosynthetic rate (Paul and Pellny 2003;McCormicket al.2006;Ainsworth and Bush 2011).However,focusing only on high sink capacity without improving the source supply capacity can lead to insufficient grain filling and declines in filled-grain percentage and grain weight,as has been shown in hybrid rice,and comparable results have been shown in previous studies (Yinget al.1998;Zhanget al.2009).For example,Huanget al.(2019) showed that the higher yield of machine-transplanted doubleseason rice planted after oilseed rape was attributable to improvements in both sink size (spikelet number m-2) and source capacity (total biomass).The grain-leaf ratio has been used as a comprehensive index for measuring the relationship between source and sink (Linget al.1993).Furthermore,the production of photosynthetic substances occurs during photosynthesis in the chloroplasts,and photosynthate can be exported only when the production of carbon compounds exceeds the local demand(Ludewig and Flügge 2013).Similarly,the sink extracts carbon compounds from the stem-sheath when the sink demand cannot be satisfied by photosynthesis (source).The transport of greater amounts of storage material from the stem to the grain is characteristic of low-yield populations (Yanget al.2008).Wuet al.(2010) also reported that the export ratio and the corresponding transport ratio of reserve substances per stem-sheath decrease significantly with increasing rice yield.Hence,the export ratio of biomass per stem-sheath (ERBS)and the transport ratio of biomass per stem-sheath(TRBS) can also reflect the coordination between source and sink.This study showed that the rice population of 260-290 seedlings m-2had lower values for ERBS and TRBS (Fig.12) and higher grain-leaf ratio (Fig.11),source supply capacity (PPR) (Fig.10),and sink capacity(TSP) (Table 1) compared with the other seedling density levels tested.Therefore,the rice population formed at the optimum seedling density showed a coordinated enhancement of sink size and source capacity.
5.Conclusion
Dry direct-seeded rice sown by a multifunctional seeder that performs synchronous rotary tillage and sowing in a rice-wheat rotation system tends to experience poor seedling establishment and yield loss because of the less-than-ideal soil conditions caused by wheat-straw return and minimum tillage.The seedling density,a fundamental consideration for high-yield rice cultivation,can regulate panicle density and panicle size.Based on the results presented here,we can preliminarily conclude that appropriately increasing the seedling density,and thereby increasing the proportion of panicles formed by the main stem,is one way to obtain a higher yield for constrained direct-seeded rice in a rice-wheat rotation system.Because,in this system,a coordinated rice population with higher total spikelet number m-2,biomass accumulation,and photosynthetic capacity was produced through sufficient main stem panicle numbers.
Acknowledgements
We are grateful for grants from the Jiangsu Agriculture Science and Technology Innovation Fund,China(CX(20)1012),the Jiangsu Demonstration Project of Modern Agricultural Machinery Equipment and Technology,China (NJ2020-58),the Jiangsu Technical System of Rice Industry,China (JATS[2020]432),the National Key Research and Development Program of China (2016YFD0300503),the earmarked fund for China Agriculture Research System (CARS-01-27),and the Yangzhou University Scientific Research and Innovation Program,China (XKYCX20_022).
Declaration of competing interest
The authors declare that they have no conflict of interest.
杂志排行

Journal of Integrative Agriculture的其它文章
- A 314-bp SlNE insertion in the ZNF2 promoter region may act as a repressor related to regulation of fat deposition in pigs
- An optimized protocol using Steedman’s wax for high-sensitivity RNA in situ hybridization in shoot apical meristems and flower buds of cucumber
- Characterization of subunits encoded by SnRK1 and dissection of combinations among these subunits in sorghum (Sorghum bicolor L.)
- The role of time preferences in contract breach: Evidence from Chinese poultry farmers participating in contract farming
- Optimal design of culling compensation policy under the African swine fever — Based on simulations of typical pig farms in China
- Drip fertigation and plant hedgerows significantly reduce nitrogen and phosphorus losses and maintain high fruit yields in intensive orchards