寄主植物对不同种群密度樟叶蜂取食后的生理防御响应
2023-01-31程柯方程远东陈伏生
程柯方,刘 俏,程远东,梁 宽,陈伏生,2*
(1.江西农业大学 林学院/江西省森林培育重点实验室,江西 南昌 330045;2.江西农业大学 鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室,江西 南昌 330045)
【研究意义】森林病虫害是中国林业经营管理中的“三害”之一,会造成林木生理、形态以及组织上的不正常变化,严重影响林木的生长、发育,甚至会导致死亡[1]。植物在遭受昆虫取食和干扰时不能像动物进行逃避避险,而是在相互作用中进化出有效的防御机制[2]。叶片作为植物感受外界环境变化的敏感性器官之一[3],在自然界中被害虫啃食后影响其光合作用,继而影响植株的生长状态和生理功能[4]。为此,植株叶片对虫害的生理防御响应研究一直受到关注,对预防和指导虫害防治具有重要的科学价值。【前人研究进展】一般认为植物是通过释放次生代谢物、植物激素以及改变养分含量等防御策略应对害虫危害[5]。次生代谢物的释放可以延缓植食性昆虫的生长、降低繁殖能力甚至造成死亡[6]。当植食性昆虫取食植物的叶片时,植物通过影响生物合成基因表达产生有毒的次生代谢物质(譬如酚类等)。而酚类物质摄入过多会限制昆虫的生长发育,提升幼虫的死亡率,其中单宁摄入会影响昆虫对营养物质的取食和消化,出现拒食现象[6]。研究表明,乌桕(Sapium sebiferum)叶片受到红胸律点跳甲(Bikasha collari)侵害时,叶片的单宁含量显著增加[7]。次生代谢物质的传递、调控和诱导主要通过激素等信号分子实现对灾害的抵御,其中,茉莉酸和茉莉酸甲酯最具代表性,在平衡植物生长发育和响应胁迫起重要作用[8]。当植物叶片受到虫害时,茉莉酸和茉莉酸甲酯进行防卫基因的表达,产生特异的诱导蛋白,有效提高植物对昆虫的防御[9-10]。同时,昆虫的取食作为植物防御的诱导因子,能引发植物的营养成分发生变化,影响氨基酸和糖类等其他营养物质组成,进而影响植食性昆虫的取食和植物的防御响应[11]。目前的研究多集中在植物对虫害产生防御物质成分的评估,但对不同密度虫害和受虫害后植物在时间动态上的响应机制及不同防御物质的协同与权衡等方面知之甚少,有待进一步研究。樟树(Cinnamomum camphora)是我国南方重要的乡土树种,集木材、观赏和固碳等多功能为一体,种植广泛,具有较高的经济和生态价值[12]。虫害防治是樟树经营管理首要任务,其典型虫害之一是樟叶蜂(Mesoneura rufonota),通过幼虫食害幼树或林木的嫩叶及嫩梢[13],严重影响林木生长。【本研究切入点】目前,樟叶蜂防治以化学防治(杀虫剂)为主,对环境危害较大,如何诱导植物自身抗虫性抵御虫害是虫害防治的关键措施。本文以樟树及食叶害虫樟叶蜂为研究体系,采用盆栽模拟控制试验,观测樟叶蜂不同取食密度后樟树叶片次生代谢物、植物激素和养分含量的动态变化,旨在了解樟树对樟叶蜂取食胁迫的响应及生理机制。【拟解决的关键问题】(1)樟树叶片防御性物质含量受不同密度虫害而有别,种群密度40 头/株虫害诱导叶片防御强度显著高于低度虫害,即前者防御性物质含量高于后者;(2)樟树叶片受虫害取食后,次生代谢物和植物激素响应快于养分含量,且前两者的时间滞后效应大于后者,即虫害停止后,次生代谢物和植物激素响应的持续时间长于养分含量。研究结果有助于理解植物与食叶昆虫的互作关系,为林业生态防治害虫提供科学依据。
1 材料与方法
1.1 试验材料
樟叶蜂幼虫来自江西农业大学森林保护昆虫实验室,该幼虫原始种群来自江西省金溪县双塘镇乌石村和对桥镇旸田村的香樟林基地。试验所用樟叶蜂虫态均为3~4 龄幼虫,为人工饲养培育的稳定种群。
2018 年4 月从江西省南昌市苗圃基地选取健康无病虫害、长势均匀、健壮的一年生樟树幼苗27 株,平均地径为0.90 cm,平均株高46.00 cm,平均分枝数为5,叶片量为40~55 片,种植于上口径为30.00 cm、下口径为25.00 cm、高为30.00 cm 的花盆中。培养土壤选取当地土壤,混合均匀,土壤pH 为4.62,全氮(TN)为1.66 g/kg、全磷(TP)为0.51 g/kg;铵态氮(NH4+-N)和硝态氮(NO3-N)分别为3.29 mg/kg和0.77 mg/kg、有效磷(AP)为0.54 mg/kg。
1.2 试验设计
盆栽培养1年后,采用单因素三水平完全随机试验设计,将樟树幼苗分为9组,每组3株重复,且每组间隔5 m 以上,并用5 mm 孔径纱网围住,防止樟叶蜂幼虫逃逸及其他昆虫进入危害。2019 年4 月,设置PD20(种群密度20 头/株)、PD40(种群密度40 头/株)和CK(种群密度0 头/株)等3 种虫害密度进行控制试验。将樟叶蜂幼虫转移到对应的樟树幼苗中,在自然光温条件下任其自由取食。由于樟叶蜂幼虫期较短,在取食5 d之后收回,更换另一批相同日龄的幼虫重复以上处理继续取食5 d,取食时间合计10 d,至此,目测害虫危害差异显著。两批樟叶蜂幼虫培养时间相同,虫龄相近,樟树幼苗种植期间维持常规且相同水分等管理措施。
1.3 取样与测定方法
在樟叶蜂取食10 d 后1 d(D1)和10 d(D10),分别在各组所有樟树幼苗上随机选取被樟叶蜂取食过的等量叶片,每个处理混合采样3 份,作为3 个重复,叶片采下保存在-80 ℃超低温冰箱,用于指标测定;D10 破坏性取植株全株叶片、枝杆以及根,将所有樟树幼苗的根、茎、叶分成3 部分,分别装进信封中,之后放进烘箱进行烘干处理,用于计算各器官生物量,3 部分器官生物量相加可得全株的生物量干质量数据。次生代谢物总酚采用Folin-酚法测定[14],单宁采用盐酸-香草醛法测定[14];植物激素茉莉酸及茉莉酸甲酯采用试剂盒法,用液相色谱仪测定[15];营养物质游离氨基酸采用茚三酮显色法[16]测定,总糖采用DNS法[16]测定。
1.4 数据处理
采用SPSS 17.0 软件对数据进行处理和统计分析。双因素方差分析虫害密度、取样时间及交互作用对叶片防御性物质含量的影响。单因素方差分析及多重比较检验不同虫害密度的影响及显著性差异,t检验分析两次取样时间的异同。相关分析剖析不同取样时间不同虫害密度驱动下叶片各观测指标的内在关联。显著性P<0.05,Origin 8.5软件作图。
2 结果与分析
2.1 叶片次生代谢物对樟叶蜂取食的响应
樟叶蜂虫害密度对取样时间D1 和D10 时叶片总酚含量均有显著影响,取样时间D1 表现为PD20 处理比CK 低19.3%,两者均与PD40 处理无显著差异,取样时间D10,PD40 处理比CK 和PD20 处理分别高20.1%和28.2%(P<0.05,图1a)。3 种虫害密度叶片总酚含量在取样时间D1 和D10 之间均无显著差异(图1a)。虫害密度未显著改变樟树叶片单宁含量;与取样时间D1相比,D10时PD20和PD40处理叶片单宁含量分别显著增加17.8%和19.5%,而CK处理两次取样时间无显著变化(图1b)。
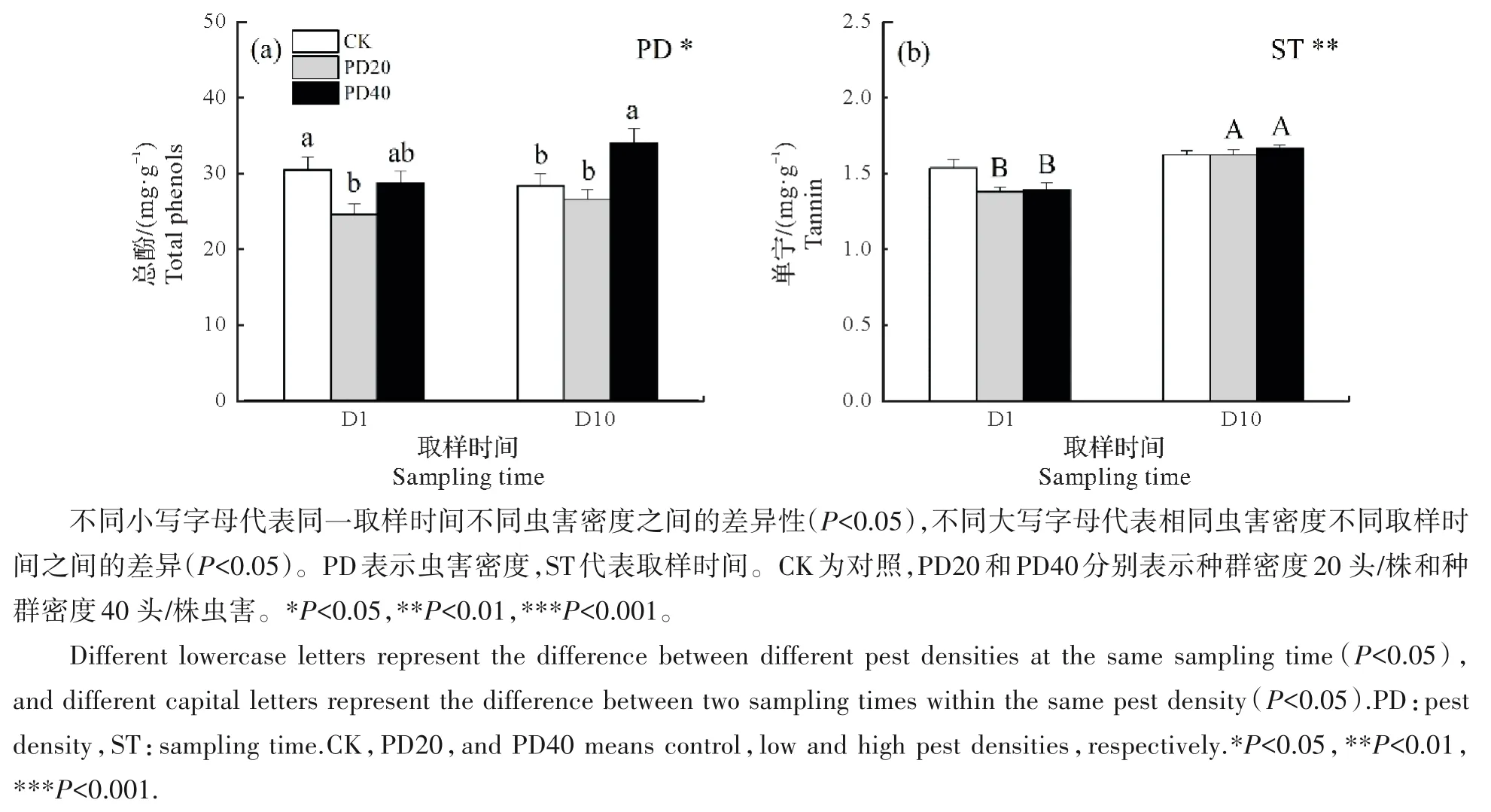
图1 叶片次生代谢物总酚(a)和单宁(b)对不同虫害密度及取样时间的响应Fig.1 Response of leaf secondary metabolites including total phenols(a)and tannin(b)to different pest densities and sampling times
2.2 叶片植物激素对樟叶蜂取食的响应
樟叶蜂虫害密度和取食后采样时间对叶片茉莉酸含量影响显著(P<0.05,图2)。与CK 相比,PD20 显著增加D1 和D10 时茉莉酸含量,分别为64.1%和127.6%,PD40 处理仅显著增加D10 时茉莉酸含量18.3%;与D1 相比,D10 时CK 和PD40 处理叶片茉莉酸分别显著减少32%和34.1%,而PD20 处理两次取样时间无显著差异(图2a)。虫害密度对D1 和D10 时茉莉酸甲酯含量均有显著影响,与CK相比,D1 时PD20 和PD40 处理分别显著降低5.3%和16%,D10 时PD20 和PD40 处理分别显著降低16.2%和25.1%(图2b)。与茉莉酸含量不同,相同虫害密度叶片茉莉酸甲酯含量在D1 和D10 之间均无显著差异(图2b)。

图2 叶片植物激素茉莉酸(a)和茉莉酸甲酯(b)对虫害密度和取样时间的响应Fig.2 Response of leaf phytohormones including jasmonic acid(a)and methyl jasmonate(b)to different pest densities and sampling times
2.3 叶片营养成分对樟叶蜂取食的响应
樟叶蜂虫害密度和取样时间对叶片总糖和氨基酸含量均有显著影响(P<0.05)。与CK 相比,叶片总糖在D1 时PD20 处理显著降低27.3%,而PD40 处理显著增加26.8%,在D10 时,仅PD40 处理显著增加25.4%。PD20 处理在D10 时叶片总糖含量比D1 增加32.6%,而CK 和PD40 处理两次取样时间无显著差异(图3a)。与CK 相比,叶片氨基酸含量在D1 和D10 时PD20 处理分别显著增加45%和47.7%(图3b)。
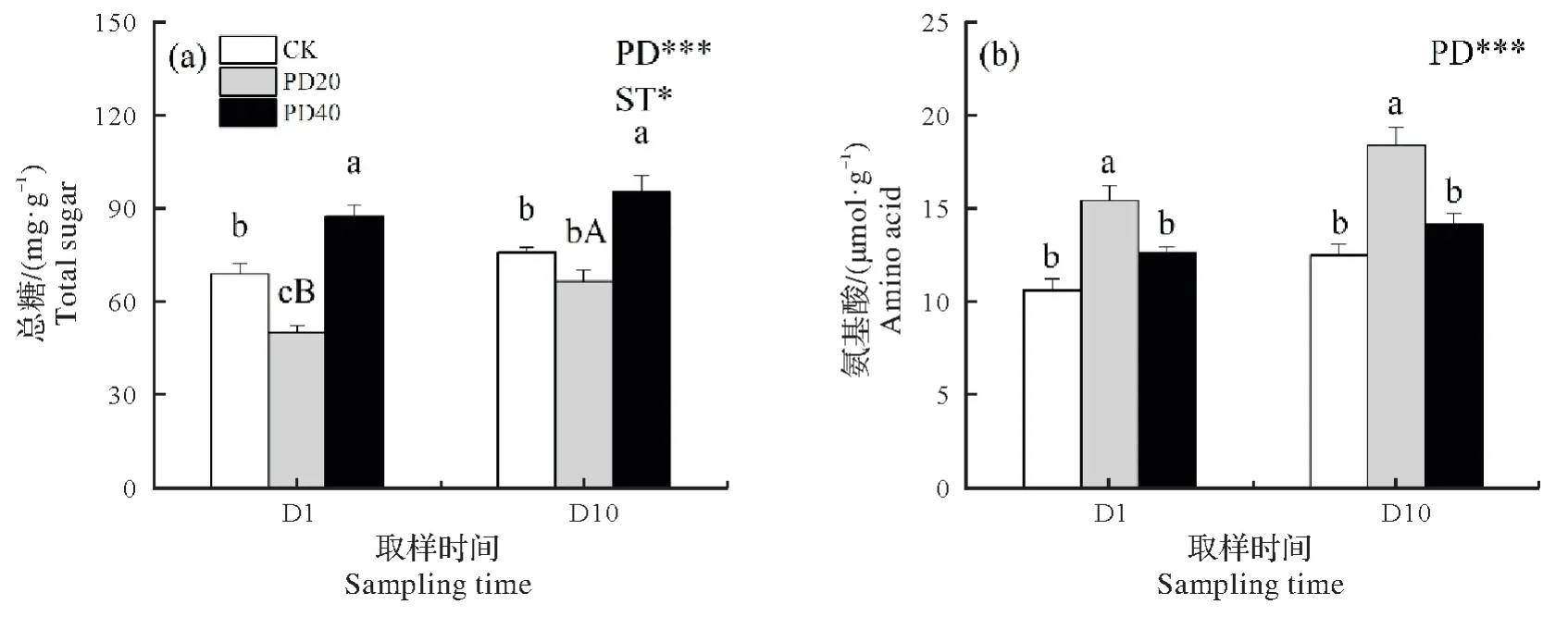
图3 叶片营养成分总糖(a)和氨基酸(b)对虫害密度和取样时间的响应Fig.3 Response of leaf nutrients including total sugar(a)and amino acid(b)to different pest densities and sampling times
2.4 幼苗地生物量对樟叶蜂虫害密度的响应及与叶片防御性物质的关联
樟叶蜂取食虫害密度对樟树幼苗叶片和总生物量影响显著(P<0.05),但未显著改变其枝杆和根生物量(P>0.05,图4)。与CK 相比,PD20和PD40处理分别显著减少叶片生物量30.4%和46.8%,显著减少幼苗总生物量的10.6%和14.2%。相关分析表明,D1 时茉莉酸与氨基酸显著正相关,总酚和单宁均与氨基酸显著负相关,茉莉酸甲酯与叶片和总生物量显著正相关,叶生物量和总生物量显著正相关。D10 时,总酚与总糖、茉莉酸与氨基酸显著正相关,茉莉酸甲酯与叶片生物量和总生物量显著正相关,叶生物量和总生物量显著正相关(图5)。
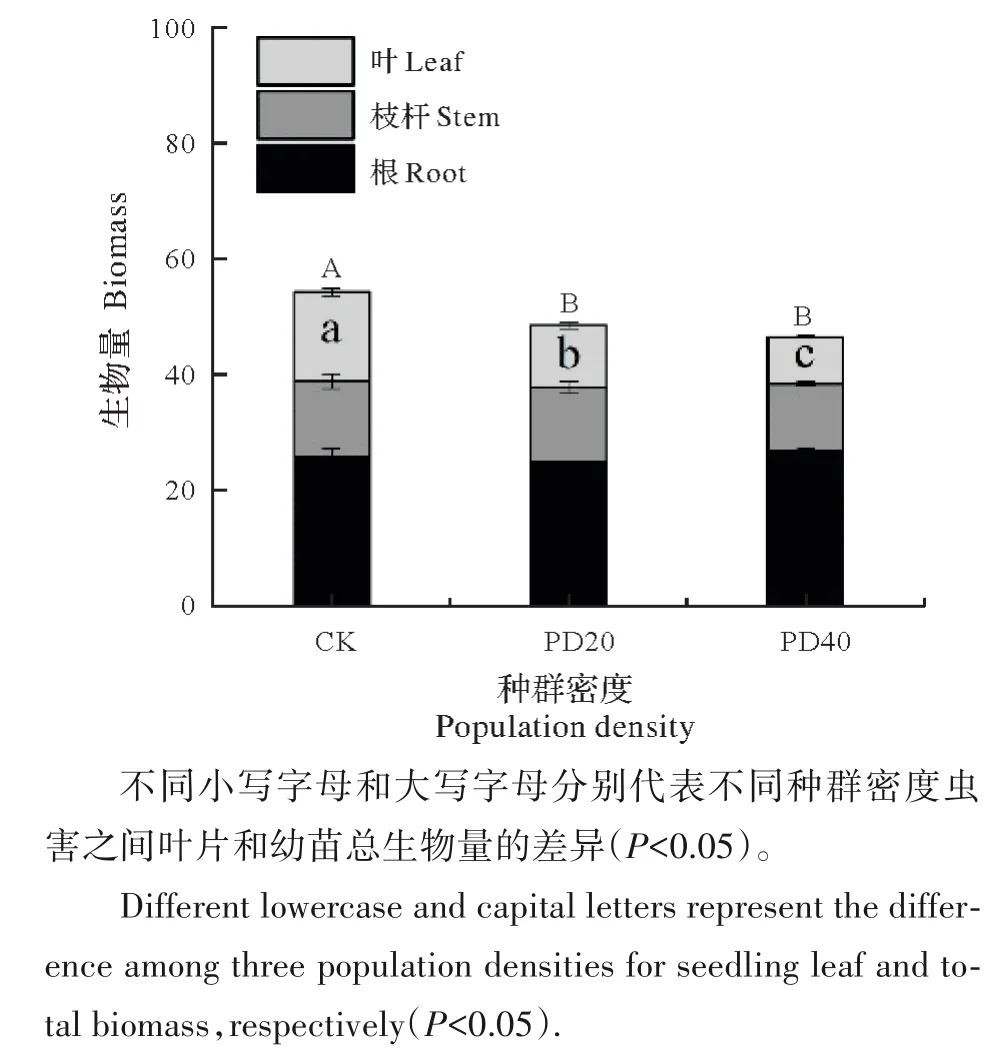
图4 不同虫害密度对幼苗叶片、枝杆、根和总生物量的影响Fig.4 Effects of different pest densities on leaf,stem,root and total biomass of seedlings

图5 虫害之后D1(左上方)和D10(右下方)时叶片次生代谢物、植物激素与营养成分等防御性物质与叶和幼苗总生物量的相关性Fig.5 Correlations of defensive substances including secondary metabolites,phytohormones and nutrients,and seedling leaf and total biomass at the sampling time of D1(upper left)and D10(lower right),respectively
3 讨论
3.1 不同虫害密度对叶片防御性物质含量的影响
植物为逃避虫害往往快速启动生理响应,生成防御性物质,但其与虫害密度的关联尚不明确[17]。次生代谢物质在植物防御中起重要作用,会导致昆虫拒食、产生排斥性,甚至中毒、直接死亡[18],研究发现种群密度20 头/株虫害导致总酚含量快速下降,但种群密度40 头/株虫害引发叶片总酚含量随时间累计而显著增加,表明樟树会通过调控叶片总酚代谢应对昆虫取食,但其响应模式与虫害密度有关。贾彦霞等[19]研究发现,西花蓟马取食诱导辣椒叶片总酚含量在短时间内不同程度的下降;高佳敏[18]研究赤霞珠葡萄应对诱导刺激时发现,除第1天外,在第3~5天昆虫取食显著增加总酚含量;这些研究结果支持了本研究的发现。但王海涛[20]研究发现,当黑松受到松褐天牛危害后,随着受害程度的增加,总酚含量先增加后减少,与本研究结果不一致,估计与松褐天牛危害茎干,而樟叶蜂直接取食叶片有关。
茉莉酸和茉莉酸甲酯等植物激素是植物应对虫害关键激素和信号分子[21],可通过调控植物次生代谢物合成,诱导抗性基因表达,产生相应化学防御物质,增强植物抗逆性[22]。本研究发现,叶片在受到樟树蜂种群密度20 头/株虫害时,其茉莉酸释放增加。而种群密度40 头/株虫害茉莉酸含量下降。一般来讲,植物防御功能具有一定的度,适宜的诱导条件下植物易产生防御性物质,而一旦超过临界值,植物防御反而会下降甚至丧失[23],这可能是造成种群密度40 头/株虫害下叶片茉莉酸含量显著低于种群密度20 头/株虫害的主因。同时,本研究发现茉莉酸甲酯随樟树蜂虫害加重而显著下降,其原因是茉莉酸甲酯作为诱导植物生长的信号分子[21],在诱导次生代谢物释放过程中被大量消耗;或者在茉莉酸甲酯合成途径中,某些基因表达水平降低所致[24]。
植物遭受虫害后,通过改变自身营养成分来适应生存状态[22]。本研究发现,叶片总糖含量因种群密度20 头/株虫害显著下降,而种群密度40 头/株虫害后显著增加;氨基酸仅种群密度20 头/株虫害显著增加,种群密度40 头/株虫害无显著变化。已有的研究也报道了叶片总糖和氨基酸因虫害降低或增加的多样化结果。如,段文昌等[25]发现枸杞瘿螨(Aceri macrodonis)取食导致枸杞叶多糖含量显著降低,何菁等[26]研究表明,烟粉虱种群趋势指数与辣椒叶片糖含量呈显著负相关,王丽丽等[27]发现,葡萄对虫害的抗性与叶片可溶性糖含量显著负相关;以上结果表明,可通过减少昆虫能量来源从而达到抵御虫害的目的。蒲恒浒等[28]发现番茄在遭受虫害和机械损伤引发叶片糖类含量显著增加,估计是植物通过加快碳水化合物积累而维持正常生长有关。与此同时,高佳敏[18]发现,虫害造成赤霞珠葡萄叶片氨基酸含量的显著增加,这与本研究结果类似。
可见,樟树叶片防御性物质含量因受不同密度虫害而响应有别,次生代谢物质、植物激素和营养物质无统一的响应模式,除叶片茉莉酸甲酯含量随虫害密度增加呈下降趋势外,其他观测指标均表现为非线性变化,本研究不支撑科学假设1 的后半部分,即种群密度40 头/株虫害处理叶片防御性物质含量并非一定高于低度虫害处理。
3.2 两次取样时间叶片防御性物质含量的变化
植物受虫害后除顺势启动防御响应外,还往往有类似“记忆免疫”的功能,即防御性物质含量并不会因虫害消除马上回复至正常水平[29]。本研究发现,虫害消除后,与D1 相比,D10 时樟树叶片单宁含量显著增加,即叶片单宁表现为继续积累。袁红娥等[30]对昆虫取食落叶松(Larix gmelinii)研究发现,在针叶遭受取食后相当长一段时间,其单宁含量不断增加至峰值;贾彦霞等[19]在辣椒叶片抗虫害研究中也发现,随着昆虫取食诱导时间的持续单宁含量先持续增加,直至1个月后恢复正常。一般认为,受虫害诱导单宁的积累是一种经济型的防御方式,植株避免防御过程中消耗自身过多资源和营养物质,故生成单宁进行防御,从而与生长形成动态平衡[29]。
茉莉酸在叶片代谢调控和防御反应发挥重要作用[8]。本研究发现对照和种群密度40 头/株虫害处理叶片茉莉酸含量均随时间延长而显著降低,而种群密度20 头/株虫害处理叶片茉莉酸含量维持相对稳定。Schittko 等[31]研究发现烟草被烟草天蛾取食后,叶片中茉莉酸含量显著增加2~10 倍,这与本研究结果相反,主要原因可能为叶片受到种群密度40 头/株虫害后,植株间传递伤害信息的途径被破坏,叶片调节代谢方式,导致茉莉酸信号分子的诱导传递受到阻碍,其含量相应减少[32]。
糖类作为植物重要的营养物质和能量来源,在抵抗虫害方面起到重要作用[17]。本研究发现,种群密度20 头/株虫害导致叶片总糖含量随时间延长显著增加,这与王丽艳等[33]发现水稻叶片受负泥虫(Lilioceris impressa)种群密度20 头/株虫害其可溶性糖显著增加的研究结果相似。
从以上分析可知,樟树叶片受虫害取食后,次生代谢物、植物激素和养分含量均表现为一定时间滞后效应,且3 类物质之间时间滞后效应无统一的模式,本研究也不支持科学假设2。但研究结果支持樟树对虫害具有自我调节能力,且对种群密度20 头/株虫害的生理防御强于种群密度40 头/株虫害。
3.3 幼苗生物量积累与叶片防御性物质的关联
虫害驱动植物生长与防御性物质合成的权衡关系一直受到关注[17]。本研究表明,叶片和总生物量与2个取样时间叶片茉莉酸甲酯均呈显著正相关,而与叶片其他防御性物质无显著相关。从叶片和幼苗总生物量的变化来看,主要源于樟叶蜂幼虫啃食樟树幼苗叶片所致。从茉莉酸甲酯的变化来看,茉莉酸甲酯转录生成主要发生于植物营养组织,叶片受虫害越重,茉莉酸甲酯催化途径受到破坏越严重,茉莉酸甲酯含量也就越少[32]。正是因为叶片和幼苗总生物量和叶片茉莉酸甲酯含量受虫害密度的影响模式相似,故其呈现出正相关。由于盆栽时间较短,并没有发现植物生长和防御性物质合成的权衡关系,有待后续试验验证。
与此同时,虫害诱导防御性物质生成,不同防御性物质也可能存在协同与权衡的关系[17]。本研究发现,D1和D10两次采样时间茉莉酸与氨基酸均呈现正相关,估计与茉莉酸是叶片防御虫害的信号分子,叶片受虫害后信号分子增多,诱导叶片合成更多特殊氨基酸防御虫害[32]。同时,还发现在D1时叶片总酚和单宁均与氨基酸呈现负相关,这与次生代谢物酚类的生物合成以莽草酸途径为主,合成过程中转变为苯丙氨酸,进而形成酚类化合物有关[6]。此外,还发现D10时叶片总酚与总糖显著正相关,表明两者对虫害响应的时间滞后效应类似。可见,受到虫害后,叶片次生代谢物、植物激素和营养物质的合成与积累确实存在协同和权衡的关系,但因虫害密度设置水平较少,相关的生理防御机制未能明确,有待进一步研究。
4 结论
樟树幼苗叶片对樟叶蜂虫害表现出生理防御响应,其次生代谢物质、植物激素和营养物质等防御性物质对两种不同虫害密度的响应模式有别。尽管不同防御性物质对叶片受虫害取食的时间滞后效应不一致,但樟树叶片对种群密度20 头/株虫害生理自我调节强于种群密度40 头/株虫害。樟树幼苗叶片和总生物量随虫害密度增加而下降,且与叶片部分防御性物质呈现出显著相关性。同时,叶片不同防御性物质也表现出较好地内在关联,生理防御性物质的协同和权衡因虫害密度而不同。研究结果对理解樟树与樟叶蜂的相互作用,指导虫害防治具有重要的参考价值,但鉴于虫害密度设置梯度有限、试验处理时间偏短等局限,樟树叶片对樟叶蜂虫害的生理防御机制还有待深入研究。
