粪肠球菌脑膜炎小鼠脑组织差异表达miRNAs筛查与鉴定
2023-01-31陈明杰李益涛曹梦园王晨豫闫雪琪齐亚银
陈明杰,李益涛,曹梦园,王晨豫,闫雪琪,陈 杰,齐亚银
(石河子大学 动物科技学院,新疆 石河子 832000)
粪肠球菌(E.faecalis)为革兰阳性细菌,通常居住在健康人和动物的肠道中,但如果破坏了脆弱的微生物群平衡,则有可能引起侵袭性感染[1-2]。近些年关于粪肠球菌致病的报道越来越多,甚至可以通过包括侵入性医疗设备在内的多种交叉污染途径轻松传播[3-4]。而粪肠球菌又是医院环境中最常见的分离菌之一,通常与医院心内膜炎、尿路感染、菌血症和牙周炎等相关[5-6]。目前已有研究表明致病性粪肠球菌不仅给卫生系统造成了巨大的经济损失,而且还使脆弱的患者容易遭受致命的感染[3,7-8]。
2005年齐亚银等[9]在患脑炎的羔羊中分离到粪肠球菌,随后周霞等[10]通过致病性试验证实该粪肠球菌是导致羔羊发生脑膜炎的致病菌,目前已经有很多研究表明粪肠球菌可以导致血脑屏障的紧密连接受到破坏[11]。由于目前对于粪肠球菌导致血脑屏障损伤的机制尚不明确,本试验选用实验室保存的粪肠球菌感染小鼠构建体内血脑屏障损伤模型。通过人工感染粪肠球菌后,利用深度高通量测序技术研究小鼠在感染粪肠球菌后,脑组织microRNA的表达情况,分析差异表达microRNA的种类及其调控的靶基因[12],并进行相关功能分析,以期为研究细菌性脑膜炎感染机制提供理论参考。
1 材料与方法
1.1 仪器与试剂凝胶成像仪(美国UVP公司);37℃恒温摇床(购迅仪器公司);CM1950 冷冻切片机(上海徕卡);Roche LightCycler®96 (瑞士Roche);伊文思蓝(麦克林公司);2×PCR MIX、DNA Marker(天根公司);1×PBS(索莱宝公司);小鼠降钙素原(PCT)ELISA检测试剂盒和小鼠C-反应蛋白(CRP)ELISA检测试剂盒(语纯公司);OTC组织包埋剂(上海樱花);PrimeScript@RT reagent Kit 反转试剂盒 (日本TaKaRa);LightCycler 480 SYBR GreenⅠ Master (北京华美生物)。
1.2 实验动物与试验菌株8周龄清洁级昆明小鼠45只,体质量22~25 g,由新疆医科大学实验动物中心提供。致羔羊脑膜炎粪肠球菌 (命名为XJ1)由石河子大学动物科技学院传染病实验室保存。
1.3 方法
1.3.1菌株复苏与模型的建立 将实验室保存的粪肠球菌XJ1进行复苏,PCR验证后采用XJ1的2/3 LD50[13]细菌数对小鼠进行腹腔注射。
1.3.2伊文思蓝(EB)法观测血脑屏障通透性 采用EB法进行检测[14],选择对照组及攻毒后12,24,60 h组,提前2 h利用尾静脉注射方法注射2%的EB。各时点将小鼠用0.9%生理盐水迅速经心脏灌注,脱颈处死,剪断鼠头,取出脑组织,制作脑组织冰冻切片后激光共聚焦显微镜观察 EB 在脑组织中的分布情况,每个时间点选用3个视野,将图片用 Image J 图像分析,并将光信号转化为数字信号,得出红色荧光密度。
1.3.3脑组织中细菌的鉴定 试验组小鼠在无菌条件下取脑组织,一部分组织做触片用于瑞氏染色;另一部分接种于BHI培养基中,在37℃、200 r/min振荡12~14 h后进行镜检,并提取其DNA用于16S RNA的鉴定。16S rRNA引物F:TGGCATAAGAGTGAAAGGCGC;R:GGGGACGTTCAGTTACTAACGT,退火温度56℃。
1.3.4血中PCT和CRP的测定 采取攻菌后12,24,48,60,72 h时各3只小鼠的全血,放置于37℃的培养箱中1~2 h待其自然析出血清后收集其血清,再分别按照小鼠降钙素原(PCT)ELISA检测试剂盒和小鼠C-反应蛋白(CRP)ELISA检测试剂盒对血中PCT和CRP进行测定并制作标准曲线。
1.3.5miRNA文库的构建 (1)分别选取对照组与攻菌后60 h小鼠脑组织样品提取总RNA,采用1%的琼脂糖凝胶电泳检测总RNA的完整性,再使用Nano Drop 2000测定其浓度及纯度是否合格。
(2)样品检测合格后,首先构建含有3′及5′磷酸基团的Small RNA样品,通过PCR将含有特殊结构的Small RNA样品反转录形成cDNA,随后以cDNA为起始模板,利用Small RNA Sample Pre Kit对其扩增得到DNA片段,AGE电泳凝胶分离,进行胶回收得到CDNA文库。测序由北京诺禾致源公司利用 Illumina Hi SeqTM2500/Mi Seq 测序平台完成。
(3)经过测序得到原始数据测Raw reads,为了保证信息分析的质量,对Raw reads进行处理,得到Clean reads。对各样品的Clean reads进行筛选,选取一定长度范围内的sRNA用于后续分析。
将sRNA比对到mRNA的外显子和内含子,找出来自mRNA降解片段的sRNA,miRNA前体的标志性发夹结构,能够用来预测新的miRNA。通过整合miREvo[15]和mirdeep2[16]这些miRNA预测软件来进行新miRNA的分析。
(4)以padj<0.05 & | log2(foldchange)|>1为条件筛选差异miRNA,通过miRanda和RNAhybrid预测差异miRNA的靶基因得到差异表达miRNA后,根据miRNA与其靶基因间的对应关系,对每组差异表达miRNA的靶基因的集合分别进行Gene Ontology和KEGG富集分析。
(5)确保测序分析结果的准确性,采用实时荧光定量PCR对测序获得的差异表达miRNA进行进一步验证。将提取的总RNA反转录合成cDNA,随机选取8个miRNAs,以U6作为内参基因,每个样品重复3次,miRNA定量上游引物为已知成熟miRNA序列(表1),下游引物为反转录试剂盒提供的通用引物,进行实时荧光定量PCR。

表1 差异miRNA引物信息
2 结果
2.1 EB测定结果在激光共聚焦显微镜下激发观察EB进入小鼠血脑屏障的变化,再通过Image J软件计算视野中平均荧光强度,如图1所示,攻菌12,24 h 脑组织平均荧光强度缓慢增加,在60 h达到最大。

图1 小鼠脑组织EB含量变化图
2.2 HE染色观察结果如图2所示,图2A为PBS处理组,可观察到小鼠脑组织的脑膜完整,界限清晰,细胞状态较好,细胞的轮廓清晰,细胞和间质无水肿现象。用XJ1号毒株的2/3 LD50进行攻毒后12 h(图2 B)少数神经细胞周围出现空白间隙,表现为脑组织轻微水肿。在24 h(图2 C)出现血管充血、血管周围的间隙明显增大。在60 h时(图2 D)脑组织的病变较明显,可观察到脑组织出现明显损伤,出现微血栓。

A.PBS处理组;B.12 h处理组;C.24 h处理组;D.60 h处理组
2.3 脑组织中细菌的鉴定如图3A所示,可在脑组织的触片中观察到双球状的细菌,该细菌的形态及大小与粪肠球菌极为相似。并且将脑组织接种于BHI培养基中的菌液进行镜检可观察到形态大小与脑组织触片一致的革兰阳性球菌(图3 B)。经PCR验证后可得知其脑中的细菌确实为粪肠球菌。可扩增出大小为296 bp大小的片段(图3 C)。

A.小鼠脑组织瑞士染色结果 (×1 000);B.脑组织增菌镜检结果 (×1 000);C.脑组织增菌细菌PCR验证结果(M.DL2000 DNA MarkerⅠ;1~6.分离菌株)
2.4 PCT和CRP的标准曲线和含量测定检测0,50,100,200,400,800 ng/L的PCT含量时的D值,制定PCT的标准曲线,结果显示其具有线性关系,其R2=9 961,方程为:y=460.920 0x-30.661 0。分别检测 0,15,30,60,120,240 mg/L含量时CRP的D值制定CRP的标准曲线,结果显示其具有线性关系R2=9 988,方程为:y=148.540 0x-4.916 6。
根据其曲线方程计算出被检血清中PCT含量,结果显示在攻菌后24,36,48 h时具有极显著性差异(P<0.01);在6,12 h时差异不显著,不存在统计学意义;在60,72 h时具有显著性差异(P<0.05)(图4)。

**.P<0.01;*.P<0.05。下同
根据其曲线方程计算出被检血清中CRP含量,结果显示在攻菌后24,36,48 h时具有极显著性差异(P<0.01);在6,72 h时差异不显著,不存在统计学意义; 在12,60 h具有显著性差异(P<0.05)(图5)。

图5 小鼠血清中CRP含量分析图
2.5 miRNA测序分析采用Illumina Hi SeqTM-2500/Mi Seq 测序平台完成测序,测序结果如表2所示。由表2可知,每个文库获得原始测序数据(Raw reads)均超过9 000 000条,对照组共获得3 164 097条Raw reads,60 h试验组获得35 378 676条Raw reads。去除受污染和低质量序列,每个测序文库获得Clean reads均超过9 000 000条,Clean reads占Raw reads的百分比>96%。选取长度18~35 nt的序列(sRNA)进行后续分析。对sRNA进行分类统计后发现,21~23 nt的sRNA数量占总sRNA的百分比>70%,其中22 nt长度的sRNA所占比例最高。将长度筛选后的sRNA定位到参考序列上,每个测序文库比对到参考序列上的sRNA总数>90%。6个文库测序结果分析得出平均GC碱基含量为49.27%,AT和GC含量基本持平。测序结果表明各文库的Q20、Q30均在90%以上,表明本测序结果质量高,所得数据可靠,可用于下一步分析。

表2 对照组和试验组miRNA测序结果
2.6 Small RNA 分类注释将所有测序所得结果进行Small RNA与各类RNA的比对、注释。由于在比对、注释过程中存在一对多的情况,按照已知miRNA>rRNA>tRNA>snRNA>snoRNA>repeat>gene>新miRNA检测的优先级顺序对各类RNA进行比对与注释。此方法将RNA作为质控标准,在质量较好的动物样品中rRNA总量占比低于40%,说明样品质量较好(表3)。
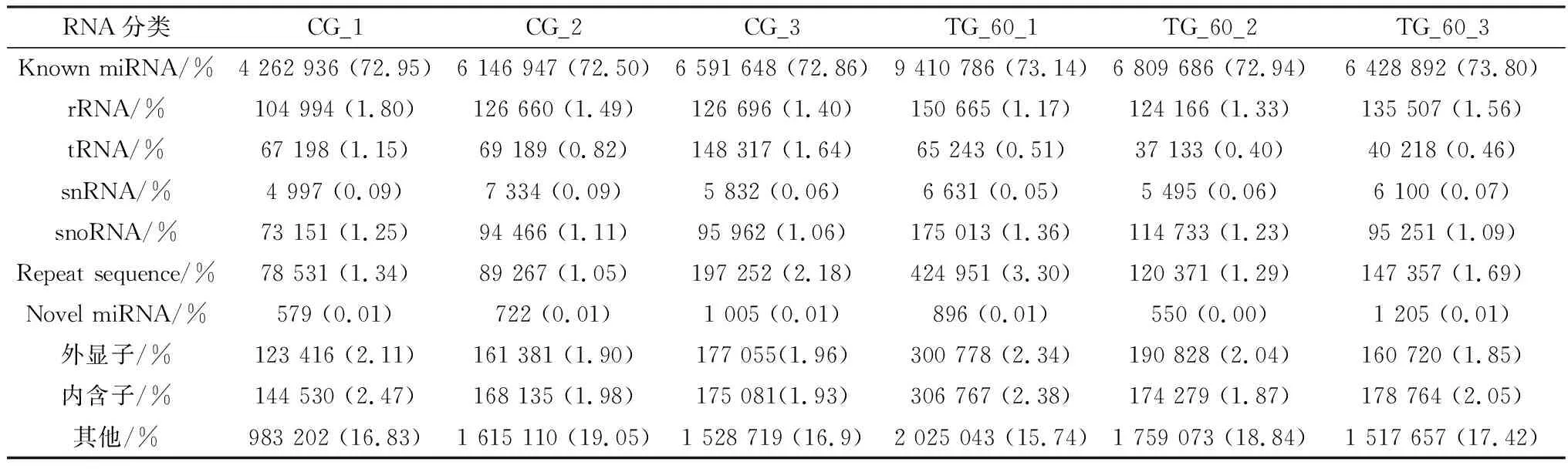
表3 sRNA的注释与统计
2.7 已知 miRNA 的分析及新 miRNA 预测通过与miRBase中指定范围序列进行比对,鉴定小鼠脑组织的miRNA,最终6个测序文库共鉴定已知mi RNAs成熟体1 376个,已知miRNA前体965个。选用miREvo和mirdeep2这些miRNA预测软件来进行新miRNA的分析,共预测到miRNAs成熟体78个,预测到前体78个。
采用TPM对miRNA表达量进行归一化处理后,使用 DEGseq 软件进行差异表达 miRNAs 的筛选,一共鉴定了22对差异表达的miRNAs,其中上调表达的 miRNAs有8个,下调表达的 miRNAs有14个(表4)。

表4 试验组和对照组中差异表达miRNAs
2.8 差异miRNA靶基因富集分析得到各组比较间的差异表达miRNA后,根据miRNA与其靶基因间的对应关系,对每组差异表达miRNA的靶基因的集合分别进行Gene Ontology和KEGG富集分析。GO分析结果表明一共进行了60个显著富集的GO条目,包括20个细胞成分、20个生物学过程和20分子功能(图6A),分子功能富集在蛋白质丝氨酸/苏氨酸激酶活性分子功能以及细胞骨架蛋白结合分子功能等。KEGG富集分析表明,差异表达miRNA的靶基因显著富集于调节干细胞多能性的信号通路、RAS信号通路、MAPK信号通路、神经胶质瘤、FcεRI信号通路(图6B)。

图6 差异表达miRNAs靶基因的GO(A)和KEGG(B)富集分析
2.9 差异表达miRNA的验证结果表明,60 h试验组挑选的8个差异表达miRNAs中6个是上调,分别为miR-34b-5p、miR-21a-5p、miR-219b-3p、miR-881-3p、miR-135a-5p及miR-34c-5p;下调的有2个,分别为miR-323-5p和miR-183-3。验证结果显示miR-34b-5p、miR-21a-5p、miR-219b-3p、miR-881-3p和miR-183-3p与对照组相比差异极显著(P<0.01),miR-34c-5p和miR-323-5p与对照组相比有显著性差异(P<0.05),miR-135a-5p无显著性差异,miRNAs的表达趋势与Small RNA测序结果一致,由此证实了Small RNA测序的可靠性(图7)。

图7 对照组和试验组中8个差异表达miRNAs的表达验证
3 讨论
研究表明miRNA在维持血脑屏障的完整性中发挥重要作用[17-20]。ARIE等[17]研究发现,将人脑微血管内皮细胞分别暴露于正常或炎症条件下培养,炎症刺激显著改变了内皮细胞中miRNA的表达水平,其中有98种miRNA经肿瘤坏死因子α和γ干扰素处理后表达下调,且伴随血脑屏障损伤,差异表达的miRNA与血脑屏障功能密切相关。有学者发现,在多发性硬化患者尸检分离的新鲜脑毛细血管中,血脑屏障相关miRNA表达水平显著下降,其中miR-125a-5p表达减少,而过表达miR-125a-5p可显著改善脑内皮细胞屏障功能。ALEJANDRO等[18]发现,miR-155在多发性硬化患者脑组织中表达增加可降低神经炎症动物模型的血脑屏障通透性。
本研究通过粪肠球菌XJ1株构建小鼠血脑屏障模型,通过检测EB、组织切片的观察以及致病菌的分离鉴定,证明该粪肠球菌确实可以穿越血脑屏障,导致小鼠产生脑膜炎症状,脑组织产生病变,并且改变血脑屏障的通透性。
同时通过探究感染后小鼠血清中CRP和PCT的含量得出,在感染致脑膜炎粪肠球菌后,小鼠体内的CRP和PCT会有所上升,而在医学上CRP和PCT已经被证明可以通过含量的变化判定脑膜炎[21]。本研究结果和贺巧峰等[22]对儿童化脓性脑膜炎和病毒性脑膜炎血清中CRP和PCT的研究结果类似,都是呈现出上升趋势,证明本试验成功构建了感染小鼠脑部模型。
选择感染症状最严重的60 h试验组小鼠和对照组小鼠脑组织样品测序,通过比对分析共鉴定了22个差异表达的miRNAs,其中上调表达的miRNAs有8个,下调表达的miRNAs有14个。本研究中鉴定mmu-miR-34b-5p和mmu-miR-21a-5p都与血脑屏障的通透性密切相关。
miR-34家族(miR34s)包含3种亚型,即miR-34a、miR-34b及miR-34c。近年来的研究证实miR-34家族能直接作用于p53,抑制细胞周期及DNA损伤应答所涉及的mRNA转录水平[23]。WELCH等[24]研究发现在神经母细胞瘤中异位表达miR-34a可以加速细胞的凋亡。更有研究证明miR-34b/c可以通过阻碍细胞周期G,抑制细胞的增殖及生长。因此miR-34家族在细胞的增殖和凋亡中起着重要的角色。刘利群等[25]的研究中发现miR-34b-5p可能在发育期惊厥性脑损伤后早期神经细胞凋亡中发挥重要作用,并且通过构建复发性癫痫大鼠模型证明miR-34b-5p介导星形胶质细胞的凋亡继而导致血脑屏障的损伤,这与本研究结果一致。
miR-21家族目前已经在30多个物种体内的组织和细胞中被发现,是在哺乳动物体中最早被发现的miRNAs分子之一。miR-21是在哺乳动物上最早被发现和确认的miRNAs分子中的一种,目前已在31个物种体内的多种组织及细胞中发现有miR-21的存在,但还没在植物上发现[26]。大量试验表明,miR-21对多种肿瘤和心血管疾病的形成和发展都具有重要影响。RAIKWAR等[27]研究发现当出现神经外伤尤其是脑外伤(TBI)时,miR-21a-5p表达明显上调,可以用来做TBI特定的生物标记。YINING等[28]证明miR-21a-5p 通过抑制 CNTF/STAT3/Nkrf 通路上调神经毒性反应性星形胶质细胞 (A1) 极化来促进创伤性脊髓损伤。更有试验表明miR-21a-5p可调节新生小鼠的小胶质细胞/巨噬细胞极化,并保护大脑免受缺氧缺血性损伤后的炎症。多种试验证明miR-21a-5p可能参与星形胶质细胞的调节,与本次测序所得到的结果相符。
miRNA靶基因预测分析对探索miRNA分子功能具有重要意义,靶基因功能注释将为基因调控机制的阐释提供研究方向。本研究通过差异表达miRNA靶基因预测及功能注释发现靶基因显著富集于Ras信号通路、癌症中的蛋白多糖、FcεRI信号通路和神经胶质瘤通路。RAS通路作为调节细胞增殖、存活、生长、迁移、分化或细胞骨架动态的信号通路;FcεRI信号通路则调节许多细胞因子,其中最重要的是TNF-α、IL-4和IL-5。这些介质和细胞因子有助于炎症反应,这些通路都会对血脑屏障的通透性产生影响,这与本研究的目的一致。综上所述,差异表达miRNA靶基因预测及基因功能注释分析,进一步揭示相关miRNA及其靶基因对血脑屏障通透性的改变发挥着重要的调控作用。
