Role of the tumor microenvironment in shaping IDH-wildtype glioma plasticity, and potential therapeutic strategies
2023-01-20LingxiangWuRuichaoChaiZhengZhaoQianghuWangTaoJiang
Lingxiang Wu*, Ruichao Chai*, Zheng Zhao, Qianghu Wang, Tao Jiang,5
1Department of Bioinformatics, Nanjing Medical University, Nanjing 211166, China; 2Institute for Brain Tumors, Jiangsu Collaborative Innovation Center for Cancer Personalized Medicine, Nanjing Medical University, Nanjing 211166, China;3Department of Molecular Neuropathology, Beijing Neurosurgical Institute, Capital Medical University, Beijing 100070,China; 4Chinese Glioma Genome Atlas Network (CGGA), Beijing 100070, China; 5Department of Neurosurgery, Beijing Tiantan Hospital, Capital Medical University, Beijing 100070, China
Diffuse glioma is the most common primary malignant brain tumor in adults. Currently, the prognosis of glioma remains dismal, and almost all patients with glioma experience recurrence even after comprehensive treatment including maximal surgical resection, radiotherapy, and/or chemotherapy1.Gliomas can be stratified according to their IDH mutation status. As we have previously reported2,3, the genetic characteristics, pathogenesis, and chemotherapy response are distinct between IDH-mutant and IDH-wildtype tumors.For instance, IDH-wildtype tumors are associated with poor prognosis and frequently have genomic alterations, including a gain of chromosome 7 and loss of chromosome 10. IDHmutant tumors often show findings including CpG island hypermethylation and MET alterations. The median overall survival of the most aggressive type IDH-wildtype glioblastoma (GBM), which accounts for more than 60% of gliomas,is less than 20 months1, mainly because GBM’s distinctive anatomic location, biological characteristics, genetic alterations,and microenvironmental features often render it resistant to conventional treatments. Therefore, alternative therapeutic strategies are urgently needed to improve patient prognosis4,5.In recent years, many efforts have attempted to target the classical hallmarks of tumors that were first summarized by Hanahan and Weinberg in 20006and further extended in 20117. However, heterogeneity within the tumor microenvironment (TME) poses a challenge to treatment efficacy, and the detailed mechanisms underlying this dynamic diversity during treatment remain elusive.
Notably, the advent of single-cell sequencing technology has enabled in-depth characterization of the cellular states of tumor cells and their TME (Table 1), and may fundamentally deepen understanding of the mechanisms underlying treatment resistance and rapid relapse. As recently discussed by Hanahan inCancer Discovery15, tumor hallmarks now include 4 new dimensions, including phenotypic plasticity, senescent cells, and non-mutational epigenetic reprogramming. These features are associated with tumor heterogeneity in response to the changing TME, and have been partially validated in IDHwildtype glioma by several large-scale single-cell studies. For instance, our previous study4has classified GBMs into 3 subtypes and demonstrated that the mesenchymal and proneural subtypes tend to exhibit high accumulation in recurrent GBMs, thus suggesting that each subtype may endow glioma with a specific ability to contribute to treatment resistance.This finding was further extended by Neftel et al.9, through an approach integrating single-cell RNA-sequencing, single-cell lineage tracing, and other methods. The authors found that tumor cells’ plasticity enables dynamic switching to 4 cellular states in response to genetic, microenvironmental and therapeutic stimuli. Moreover, Ravi et al.16have shown that GBM has segregated niches characterized by distinct immunological and metabolic stress factors, thereby indicating the effects of the microenvironment on the development of tumor cells.Importantly, hypoxia induces defined transcriptional andgenomic responses, including copy number alterations, thus suggesting that the TME affects not only transcript plasticity but also genetic alterations in gliomas.

Table 1 Current studies on tumor plasticity and the TME based on single-cell sequencing technologies
IDH-wildtype gliomas appear to have higher plasticity than IDH-mutant tumors. Johnson et al.10have shown that tumor plasticity is accompanied by non-mutational epigenetic reprogramming, as confirmed through integrated analysis including single-cell DNA methylomes and single-cell transcriptomes. In both hypoxia and irradiation experiments, IDH-wildtype tumor cells have a tendency toward exhibiting perturbed DNA methylation, which is associated with cellular state shifts. Moreover, these findings are more likely to occur in IDH-wildtype tumors, which exhibit greater genomic instability than IDH-mutant tumors. A similar finding by Chaligne et al.11has indicated that the heritability of malignant cell states shows key differences in hierarchal and plastic cell state architectures between IDH-mutant glioma and IDH-wildtype tumors, and IDH-wildtype tumors have higher cellular plasticity than IDH-mutant cells. Moreover,the Glioma Longitudinal AnalySiS (GLASS) Consortium has demonstrated that glioma progression is shaped by both genetic evolution and microenvironment interactions,viaanalysis of transcriptional profile alterations in more than 300 longitudinal cases17. Recurrent IDH-wildtype tumors show up-regulation of neuronal signaling programs or epithelial-mesenchymal transition pathwaysviainteraction with myeloid cells, whereas recurrent IDH-mutant tumors often show a proliferative phenotype accompanied by CDKN2A deletion or hypermutation. Overall, tumor plasticity is an emerging hallmark of gliomas for maintaining the distribution of various cellular states, and is highly associated with genetic features and microenvironment interactions.
Among the cellular states, the senescent-like state is now receiving attention for its versatile roles in preventing apoptosis, inducing angiogenesis, promoting invasion, and suppressing immunity15. Senescent cells are induced by pressures such as hypoxia, but the fully viable tumor cell phenotype recovers after the TME changes, thus resulting in rapid disease progression.By activating the senescence-associated secretory phenotype(SASP), senescent cells release cytokines (e.g., CCL2), which recruit immune cells, such as macrophages, that infiltrate into tumor tissue18. Macrophages are a major component of the TME (30%–50%) and are widely involved in treatment resistance4and immunosuppression19. Furthermore, macrophages have recently been reported by Hara et al.12to secrete OSM,which in turn interacts with OSMR or LIFR in complex with GP130 on GBM cells, thus activating the STAT3 signaling pathway and inducing tumor cells to switch to a mesenchymal-like state—a phenotype associated with macrophage recruitment4.Therefore, a forward feedback loop of interactions between tumor cells and macrophages may sustainably promote tumor plasticity and immune cell enrichment (Figure 1).
Perspectives

Figure 1 Forward feedback of the interplay between tumor cells and macrophages in promoting tumor progression. The loop includes 4 steps: (1) tumor cells promote macrophage recruitment through factors including CSF1 and CLL2; (2) macrophages activate polarization;(3) the polarized macrophages induce tumor plasticity by releasing factors including OSM; (4) and tumor cells switch cellular states(e.g., mesenchymal state).
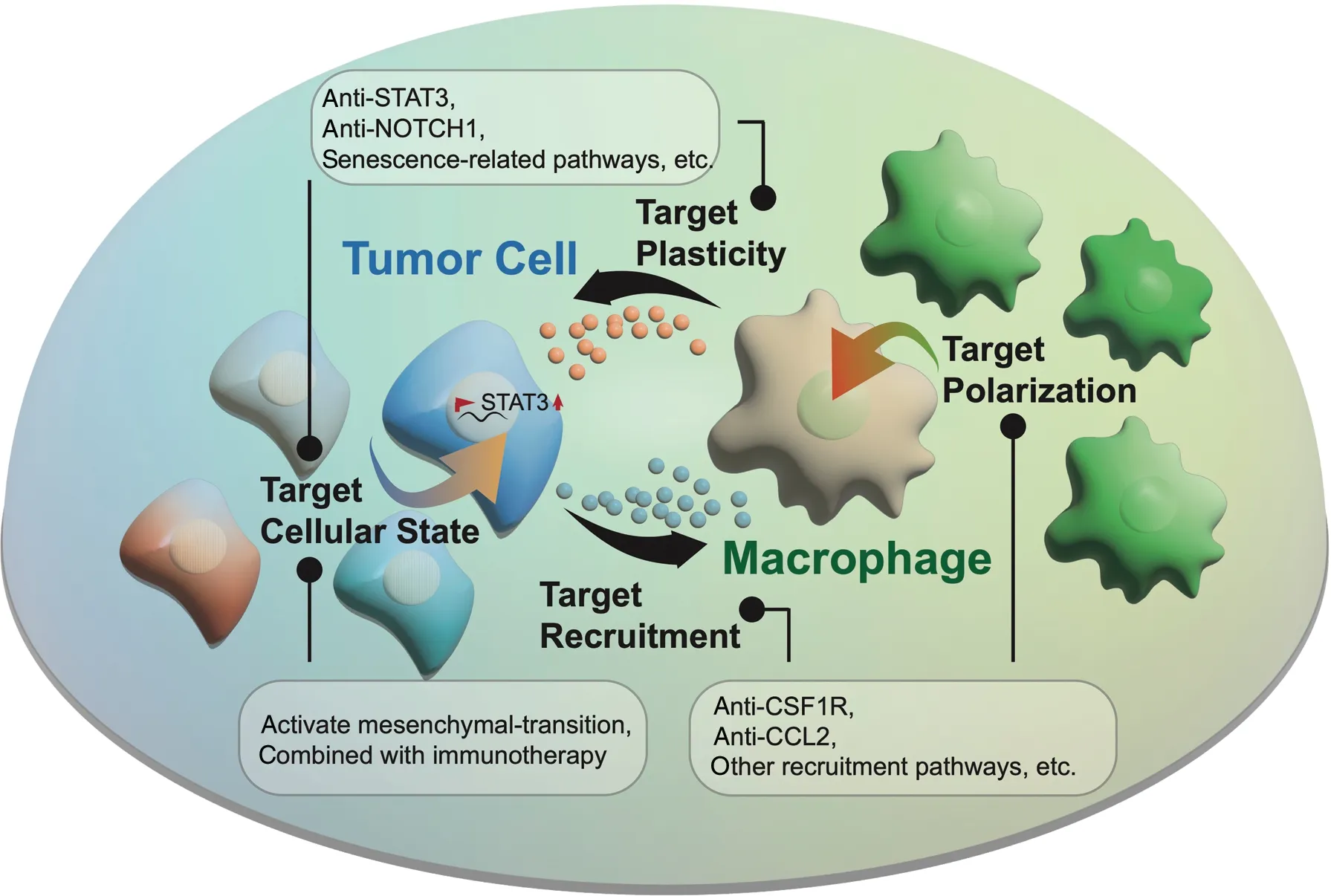
Figure 2 Potential treatment strategies: (1) blocking macrophage recruitment and polarization, such as with anti-CSF1R, anti-CCL2, and other potential recruitment/polarization pathways; (2) inhibiting tumor plasticity and intervening in tumor cellular state switching, such as with anti-STAT3 and anti-senescent cells. An alternative approach involves combining activating mesenchymal transition and immunotherapies.
Currently, a growing body of convincing published evidence20,21indicates that the intense interplay between tumor cells and the TME has a pivotal role in maintaining angiogenesis, an immunosuppressive microenvironment, and tumor cell plasticity.Stromal cells such as cancer-associated fibroblasts remodel the surrounding extracellular matrix—a process increasing GBM invasion. However, the detailed molecule mechanism underlying this influence on tumors remains to be further investigated22.Endothelial cells are an essential component of the vasculature.The VEGF-driven signaling pathway is a well-known classical regulatory axis, which has an essential role in promoting angiogenesis and tumor growth. Substantial efforts have focused on targeting VEGF, but little progress has been made23. The development of immune therapy has spawned a series of new targeted therapies for reshaping the TME. For example, several studies24,25have applied immune checkpoint inhibitors (e.g.,anti-PD1) to reactivate immunity and kill tumor cells. Other efforts have focused on developing chimeric antigen receptor T cell immunotherapy or vaccines based on classical mutations(e.g., EGFRvIII) in tumor cells26,27. However, most of these studies have shown only limited improvements in prognosis.Given the versatile roles of macrophages in the TME, targeting macrophages has also been considered a promising strategy to prevent progression (Figure 2). Preclinical studies have used emactuzumab to target CSF-1R, a macrophage colony-stimulating factor receptor, thus successfully suppressing macrophage recruitment and the M2-like polarization phenotype in mice28.However, the recruitment pathways between tumor cells and macrophages are complex and diverse. Except for cytokines such as CCL2 attracts macrophages18,29, our previous study has indicated that NF1 deficiency may activate an alternative pathway inducing macrophage infiltration4. Therefore, systematic clarification of the mechanism of macrophage recruitment may provide more opportunities to identify potential strategies for improving prognosis. Other alternative strategies include direct inhibition of tumor plasticity. For example, regulators (e.g.,STAT3 or NOTCH1)12associated with the tumor state transition might be potential targets for treatment. Furthermore, systematic characterization of the molecular traits of senescent-like tumor cells with single-cell sequencing technology will also be essential to provide potential novel targets for remodeling the TME and preventing tumor relapse.
Grant support
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 81761168038,81903078, 81972358, and 91959113), and the Beijing Nova Program (Grant No. Z201100006820118).
Conflict of interest statement
No potential conflicts of interest are disclosed.
杂志排行

Cancer Biology & Medicine的其它文章
- Targeting PP2A for cancer therapeutic modulation
- Epidemiological characteristics and genetic alterations in adult diffuse glioma in East Asian populations
- Clinical management and survival outcomes of patients with different molecular subtypes of diffuse gliomas in China(2011–2017): a multicenter retrospective study from CGGA
- Radiotherapy delays malignant transformation and prolongs survival in patients with IDH-mutant gliomas
- Insights into the transition of ductal carcinoma in situ to invasive ductal carcinoma: morphology, molecular portraits, and the tumor microenvironment
- Preoperative ultrasound combined with routine blood tests in predicting the malignant risk of pancreatic cystic neoplasms