食果鸟类对秋枫果实的取食和传播研究
2023-01-19汪国海李福燕涂文馨黄秋婵唐创斌周岐海
汪国海,李福燕,涂文馨,黄秋婵,唐创斌*,周岐海
(1.广西民族师范学院 化学与生物工程学院,广西 崇左 532200;2.珍稀濒危动植物生态和环境保护教育部重点实验室(广西师范大学),广西 桂林 541006;3.广西珍稀濒危动物生态学重点实验室(广西师范大学), 广西 桂林 541006)
周岐海(1976—),男,广西贵港人,广西师范大学教授,博士。E-mail:zhouqh@ioz.ac.cn
种子传播是决定植物种群空间结构、动态和组成的一个重要过程[1]。许多研究表明,70%~90%的热带雨林和亚热带雨林植物,其种子都是依靠脊椎动物进行传播[2-3],尤其是鸟类。这是因为鸟类种类和数量众多且不同鸟类间的食性、行为和体型特征存在明显差异,同种鸟类能同时取食多种不同尺寸的植物种子,并对其进行有效传播[4-5];而且鸟类具有较强的飞行能力,消化道中的植物种子能伴随其日移动距离的增加而被传播到适宜萌发的微生境中,并占据新的生态位,扩大植物种群的空间分布[6],从而降低子代与亲代间对光照、水分和营养等资源的竞争,减少后代因密度制约效应造成的死亡率[7]。
鸟类对植物果实的取食和传播是一个复杂的动态过程,受母树的结实率、果实大小、母树高度、动物体型特征(体长、体质量和喙长)及次生物质浓度等多种因素共同影响[8-9]。研究发现,访问南方红豆杉Taxuschinensis目标母树的鸟类数量和种数均与母树结实量呈显著正相关,而鸟类数量与母树郁闭度呈正相关[10];鸟类对香樟Cinnamomumcamphora果实的取食量均与其体长、体质量和喙长呈显著正相关[11];同等条件下,大型鸟类比小型鸟类取食更多的种子,加之其较强的飞翔能力和活动范围,能将植物种子传播到不同类型的生境中[12]。
秋枫Bischofiajavanica又称红桐、乌杨,隶属于大戟科Euphorbiaceae秋枫属Bischofia,是热带和亚热带常绿季雨林中的主要树种[13],广泛分布于越南、印度、日本、澳大利亚及我国云南、四川、贵州、广西、广东及台湾等省区[14]。秋枫4—5月开花,果实9—10月开始成熟,果实呈浆果状(长度: 10.08±0.06 mm、宽度: 12.40±0.07 mm、质量: 0.99±0.01 g,n=100)。秋枫果实成熟后会一直悬挂至翌年2月,可能是通过延长种子悬挂时间来吸引更多的鸟类对其种子进行取食和传播,但目前还未见有关鸟类在秋枫果实传播中作用的相关报道。本文以分布在广西民族师范学院内的秋枫为研究对象,收集鸟类的取食行为数据,并分析鸟类体型特征、秋枫母树特征对食果鸟类取食行为的影响,拟为进一步研究鸟类与植物间的种子传播网络提供基础参考数据。
1 研究区域与方法
1.1 研究区域概况
研究区域位于广西崇左市广西民族师范学院校园内(22°23′N, 107°23′E)。该地属于亚热带季风区,气候温和、雨量充沛,常年光热充足;年平均温度22 ℃,7月最高温度36 ℃,1月最低温度1 ℃;年降雨量1 200 mm,有明显的旱季和雨季之分(雨季为4—9月,旱季为10月至翌年3月),其中80%的降雨量都集中在雨季;年无霜期达340 d,年日照时数高达1 600 h。校园内种植有大量的果实植物,包括海南蒲桃Syzygiumcumini、秋枫Bischofiajavanica、桂花Osmanthusfragrans、萍婆Sterculianobilis、假萍婆Sterculialanceolata、香樟Cinnamomumcamphora、雅榕Ficusconcinna和海桐Pittosporumtobira等。
1.2 研究方法
秋枫作为行道树被种植在校园道路两侧,数量多,属于优势树种。2020年11月至2021年1月底,随机选取9株相对集中且易于观察的秋枫母树作为目标观察树,每天7:00—10:00、16:00—18:00借助Safari 10×26变焦双筒望远镜对访问目标母树的鸟类取食行为进行连续观察,并详细记录鸟类种类、数量、访问频次、取食时间、取食数量、取食基质和取食方式等,直至其离开观察树为止[15]。若一群同种鸟类同时访问目标母树,又无法对所有鸟类的取食行为进行同步观察时,则选择最适宜观察的一只个体进行记录,并将以整吞方式取食秋枫果实的鸟类定义为种子传播者[16]。所有观察都在晴朗的天气中进行。
同时,测量目标母树的特征(结实量、郁闭度和高度),具体方法:1)郁闭度用林冠的投影面积与林地面积之比来表示[17];2)结实量则选取母树一个小枝的果实进行计数,再计算母树的所有小枝数以统计果实量[10];树高的测量采用激光测距仪(深达威, SW-M70)。
1.3 数据处理
采用Shannon-Wiener指数、优势度Simpson指数和均匀度Pielou指数对访问目标母树的鸟类多样性进行分析:
(1)
(2)
J=H/Hmax。
(3)
式中:S为鸟类的数目;H为Shannon-Wiener指数;C为Simpson指数;Pi为第i种鸟的个体数量占全部鸟种个体数量的比例[18];J为均匀度Pielou指数;Hmax为理论上最大的多样性指数[19]。
采用 One-SampleT-test 对鸟类的访问频次、平均取食时间、平均取食数量差异进行分析;以不同鸟类对秋枫果实取食方式(啄食或整吞)的频次为基础数据,采用bipartite函数绘制取食网络图;对秋枫母树特征与访问鸟类种类、数量及多样性指数间的关系、鸟类形态特征与取食量间的关系进行相关性分析。鸟类形态特征(体质量、体长和喙长)的所有参数来自《中国鸟类野外手册》[20]和《中国鸟类志》[21]。所有数据的分析均在SPSS 20.00上完成,并将显著水平设定为P<0.05。采用R语言(3.6.1)作图[22]。
2 结果
2.1 不同鸟类取食行为差异
在42 d的观察中,共记录到5种鸟类(1目2科)取食秋枫的果实行为1 002次,具体见表1。虽然不同鸟类对秋枫果实的访问频次不存在显著差异(t=1.488,df=4,P=0.211),但红耳鹎Pycnonotusjocosus和白头鹎Pycnonotussinensis的访问频次最高(表1、图1),分别占总频次的74.04%和16.65%。不同鸟类的平均访问只数、取食时间和取食数量间均存在极显著差异(P<0.01),其中红耳鹎的取食数量(2.52±0.05颗/次)和持续时间(27.38±0.38 s/次)均最大。取食时,鸟类多集群而来,其中暗绿绣眼鸟Zosteropsjaponicus群最大为2.67±0.31只/次,黄臀鹎Pycnonotusxanthorrhous最少为1.64±0.14只/次。从取食方式上看,有4种鹎科鸟类同时以啄食和整吞的方式取食秋枫果实,对秋枫种子具有潜在的传播作用;而暗绿绣眼鸟只啄食秋枫果实的果肉部分,故不起传播作用(表1、图1)。

表1 取食秋枫果实的鸟种类Tab. 1 Species of frugivorous birds feed on the fruits of Bischofia javanica
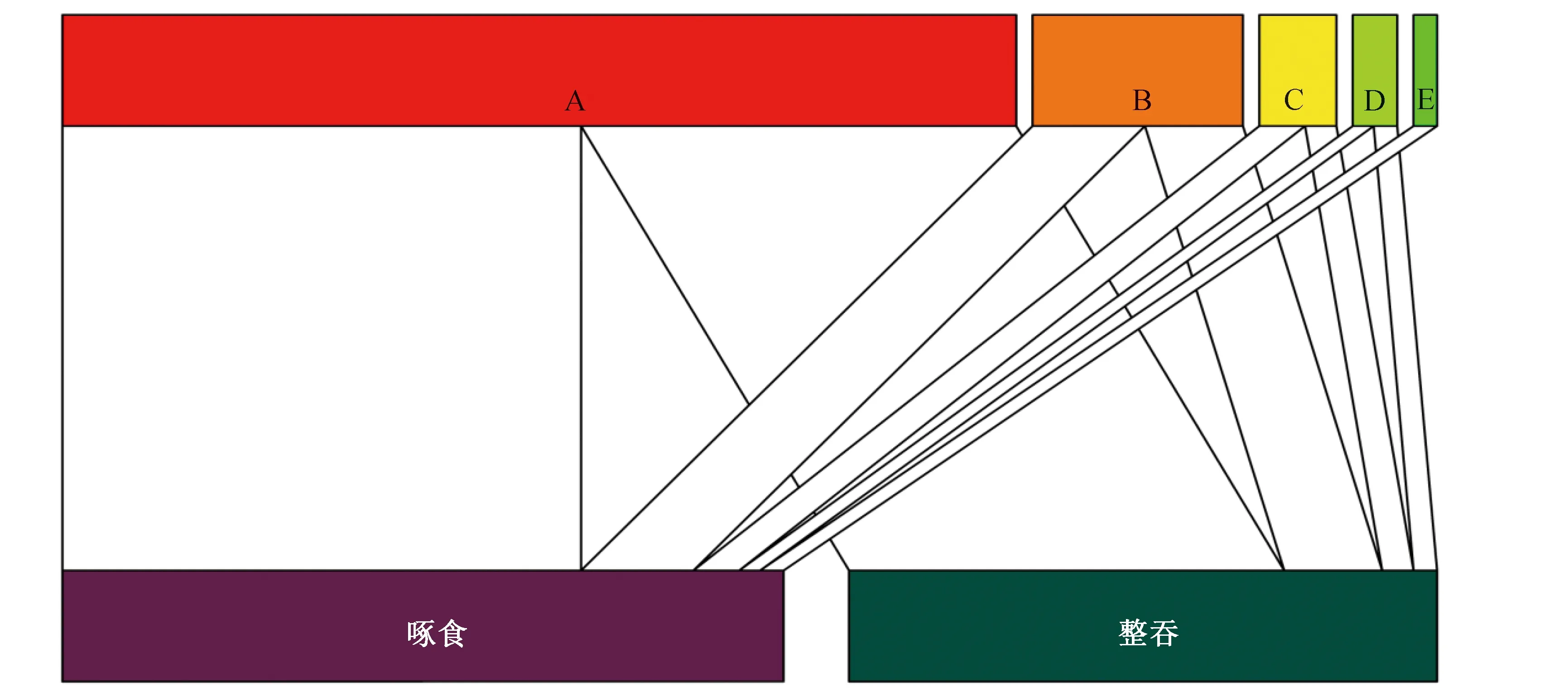
A.红耳鹎;B.白头鹎;C.黄臀鹎;D.白喉红臀鹎;E.暗绿绣眼鸟先统计出每种鸟类以啄食或整吞方式取食秋枫果实的频次,然后以此为基础绘制取食网络图,连线的宽度越大,说明鸟类以该种方式取食秋枫果实的频次越多图1 鸟类与秋枫果实间的取食网络Fig. 1 Foraging network between birds and fruits of Bischofia javanica
2.2 动植物特征对鸟类取食行为的影响
访问鸟类多样性与秋枫母树特征的相关性分析结果见表2。分析数据表明,秋枫母树结实量与访问母树的鸟类数量(P<0.001)、种数(P=0.001)、Shannon-Wiener指数(P=0.033)及Simpson指数(P<0.001)间均有显著正相关性,说明随着母树结实量的增加,鸟类访问数量、种数和多样性指数也增加。但母树结实量并不影响均匀度指数(P=0.681)。母树郁闭度与访问母树的鸟类数量(P=0.039)间具有显著正相关性,说明访问母树的鸟类数量随着郁闭度的增加而增加,但与访问鸟类种数(P=0.150)、Shannon-Wiener指数(P=0.567)、Simpson指数(P=0.807)和均匀度指数(P=0.422)间均无显著相关性。母树高度与访问母树的鸟类数量(P<0.001)和种数(P=0.002)间均具有显著正相关性,但与Shannon-Wiener指数(P=0.076)、Simpson指数(P=0.214)和均匀度指数(P=0.505)间均无显著相关性。
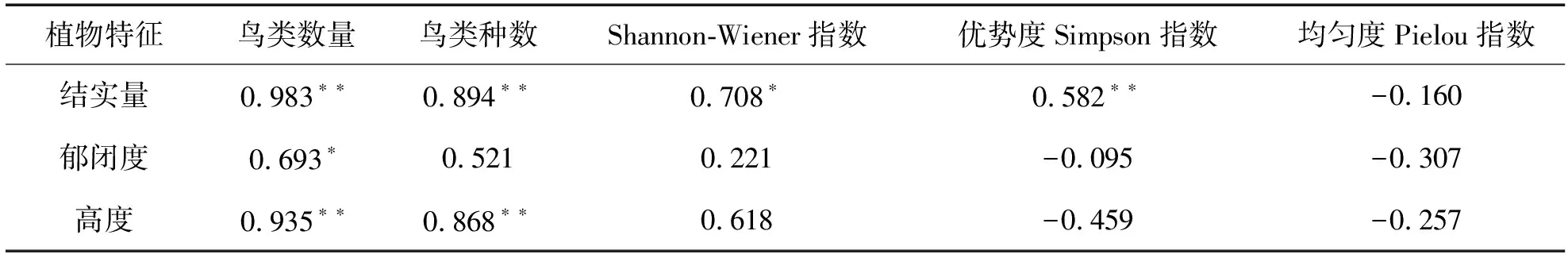
表2 访问鸟类多样性与秋枫母树特征的相关性Tab. 2 Correlation between visiting bird diversity and characteristics of Bischofia javanica
平均取食量与鸟类形态特征的相关性分析结果见图2。由图可见,鸟类的体长(r=0.885,P=0.046)、体质量(r=0.950,P=0.013)和喙长(r=0.894,P=0.041)都与平均取食量间呈正相关,说明鸟类体型越大,其对秋枫植物果实的平均取食量越多。
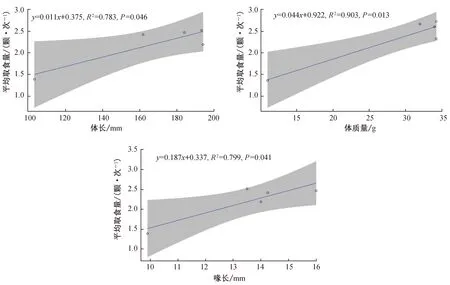
图2 平均取食量与鸟类形态特征的相关性Fig. 2 Relationship between average foraging quantity and morphological characteristics of birds
3 讨论
植物为了尽可能地吸引更多的食果鸟类对其果实或种子进行取食和传播,在长期协同进化过程中分化出一系列的特征,例如颜色、大小、形状、营养组成和成熟时间等[23],其中颜色(红色和黑色)是植物最常用的视觉吸引剂[24-25]。秋枫果实成熟后由青色转变为棕褐色,与周围植物的绿色背景形成强烈的视觉反差,从而对空中飞行鸟类产生强烈的视觉吸引作用,但研究期间仅发现5种鸟类取食秋枫的果实(表1)。较少的取食鸟类可能是由于同时期校园中的海桐、香樟等肉质果植物也正处于成熟期,鸟类对食物资源有更多的选择,从而降低了访问秋枫母树的鸟种类。
鸟类对植物果实或种子的取食方式常会随果实或种子尺寸的大小而发生改变[26]。例如:整吞小型种子后,鸟类会以粪便的形式将其排出体外,而尺寸较大的种子常以呕吐的方式传播,2种方式都有利于植物种子到达适宜萌发的生境中,以实现其种群的自然更新[27]。本文研究中,秋枫果实的宽度明显大于暗绿绣眼鸟喙的长度,同时受自身的体质量和消化道容量限制,故暗绿绣眼鸟仅能以啄食果肉的方式取食秋枫果实,而4种鹎科鸟类的喙长较大,选择比较随意,既可整吞又可啄食,属于秋枫果实的潜在传播者(表1、图1)。其他研究也发现,秋冬季节鹎科鸟类食物组成中含有大量的植物果实,且能以多种方式取食不同大小的植物果实[28-29]。
访问母树的鸟类多样性决定了种子或果实的潜在传播有效性[10]。本文研究中,秋枫结实量与访问母树的鸟类数量、种数、Shannon-Wiener指数及Simpson指数间均有显著相关性;母树郁闭度与访问母树的鸟类数量间具有显著相关性(表2),这可能是由于较高的母树结实率意味着食物资源的可获得性偏高,故能吸引大量的鸟类取食其果实,同时,郁闭度高的母树能为鸟类提供一个较为安全的取食和栖息场所。秋枫母树的高度也会影响访问的鸟类数量和种类(表2),这与其他研究[10]存在一定的差异。这种差异可能与本研究在校园中进行且人为干扰较大有关,故鸟类倾向于选择较高的母树取食,而鸟类的警戒行为是其一种自我保护的本能。
鸟类的形态特征(体质量、体长和喙长)会影响其对植物果实或种子的取食数量[30]。许多研究表明,鸟类对种子的取食数量与其形态特征间呈显著正相关[31-32],这是因为鸟类的体型越大意味着其具有更大的消化道容量,能一次性取食更多的植物种子,加之大型鸟类具有较强的飞行能力和活动范围,能将植物种子传播到不同的生境中,从而提高植物种群的空间分布[33]。本文研究结果同样显示,鸟类的体质量、体长和喙长与鸟类的取食数量间均呈正相关关系(图2),即体型特征的数据越大,鸟类的取食数量越多。
观察期间,大多数鸟类取食完秋枫果实后,会直接停歇在母树周边的柚木Tectonagrandis和降香黄檀Dalbergiaodorifera树上,待消化完果肉部分后再以呕吐的方式将种子排出体外;同时,在鸟类停歇的树木下方地面上,散落着大量裸露的秋枫种子,且偶尔看见鸟类衔着秋枫果实藏于灌木丛中,可见鸟类的取食行为能促进秋枫果实的传播,二者间能形成稳定的互利互惠关系。
4 结语
本文研究表明,鸟类有助于城市绿地中的植物种子传播,对维持城市生态系统结构和功能的稳定性具有重要意义。因此在城市化建设过程中,应考虑适当搭配诸如种植不同类型鸟类偏好的果实植物及植被结构等栖息地植物景观因素,借助鸟类的取食和传播行为来提高不同斑块间的连通性,进而提高城市生态系统中的物种多样性。
