水杨酸对铁观音茶树幼苗生理特性及茶氨酸代谢的影响
2023-01-18陈育才
陈育才
福建省南安市丰州镇乡村振兴服务中心,福建 泉州362333
茶氨酸是茶树的特征性氨基酸,是茶树体内最重要的次级代谢产物之一,具有抗氧化、增强机体免疫、松弛神经紧张等生理活性作用[1-2],在食品和医药领域有着广泛应用[3-4]。
水杨酸是一种普遍存在于植物体内的小分子酚类化合物,可调节植物的各种代谢途径[5-6],外源水杨酸提高植物抗性的研究已经取得一定进展[7-9]。董磊等[10]研究表明,外源水杨酸能通过提高叶绿素含量和过氧化物酶(POD)活性等方式有效缓解高温胁迫对金花菜的生理影响。在淹水胁迫下,外源水杨酸可提高活性氧清除系统中的超氧化物歧化酶(SOD)和POD的活性,提高可溶性糖、可溶性蛋白质的含量,降低电解质渗漏率和丙二醛(MDA)含量,从而增强辣椒幼苗耐淹水胁迫的能力[11];在干旱胁迫下,外源水杨酸可有效改善凤仙花的生长发育,保护光合色素,提高抗氧化酶活性,上调P5CS、P5CR基因表达及促进脯氨酸合成,从而降低干旱胁迫导致的脂质过氧化和氧化损伤[12];外源水杨酸能显著缓解干旱胁迫下大豆净光合速率、气孔导度、实际光合效率、用于代谢的能量、氮代谢关键酶活性和抗氧化酶活性的下降,降低干旱胁迫下用于米勒反应和热耗散的能量,减少叶片过氧化氢含量,缓解干旱胁迫下的光抑制,提高大豆对干旱胁迫的抗性[13];水杨酸浸种对盐胁迫下醉马草种子萌发和幼苗生长均有促进作用[14];水稻在受到水杨酸诱导后,激活了水稻体内的多种防卫反应、促进了水稻的病程相关基因的表达,组蛋白乙酰化修饰水平提高,从而使水稻幼苗稻瘟病抗性提高,即产生了诱导抗性[15]。
目前,很多学者主要集中研究茶树栽培方式、外源氮肥、微量元素等方面的调节对茶氨酸代谢的影响[16-17],而使用外源水杨酸调节茶氨酸代谢的研究尚未见报道。本研究以适制乌龙茶品种铁观音幼苗为试材,采用不同浓度的水杨酸对2年生盆栽幼苗进行叶面喷施,分析水杨酸对茶树光合性能、抗氧化特性、茶氨酸代谢途径相关酶基因表达的影响,从而为茶树高质量栽培提供理论依据。
1 材料与方法
1.1 试验材料
以2年生乌龙茶品种铁观音幼苗为试验材料,试验在安溪县茶叶科学研究所生产基地内进行。茶树幼苗种植于花盆中,生长健壮,地径1.1 cm左右,植株高85 cm左右,每盆种植2株,每盆装土4.5 kg;预培养1个月后,选择长势相似的幼苗作为试验材料。
1.2 试验设计
于2019年4—5月对茶苗叶面喷施水杨酸,设置4个喷施浓度:1.0 mmol/L(处理1)、2.0 mmol/L(处理2)、3.0 mmol/L(处理3)和4.0 mmol/L(处理4),以喷施蒸馏水为对照(CK),共5个处理,随机区组排列,3次重复,各处理间距离30 cm。叶面喷施于上午9:00前进行,喷施适度为所有叶片滴液,同时喷施等量蒸馏水作为对照处理;每隔1周喷施1次,共喷施4次;试验期间为防止自然降水影响,设置适当避雨设施。试验期满,分别取样各处理与对照植株的同方向、同节位枝条上的根颈部以上第三片成熟叶片,进行相关指标测定。
1.3 试验方法
1.3.1 生化检测
参照《茶游离氨基酸总量的测定》(GB/T 8314—2013)测定茶氨酸含量,比色法测定MDA含量,考马斯亮蓝法测定可溶性蛋白质含量,比色法测定POD活性,分光光度法测定叶绿素含量,水浴浸提法测定脯氨酸含量,蒽酮比色法测定可溶性糖含量[18]。
1.3.2 茶树茶氨酸代谢途径相关酶基因的表达
采用RNA提取试剂盒提取RNA;以茶树叶片总RNA为模板,采用Perfect Real Time合成cDNA;以cDNA为模板,荧光定量分析采用SYBR Premix Ex Taq TMII试剂盒,引物由上海生工公司合成,引物系列设计见表1。
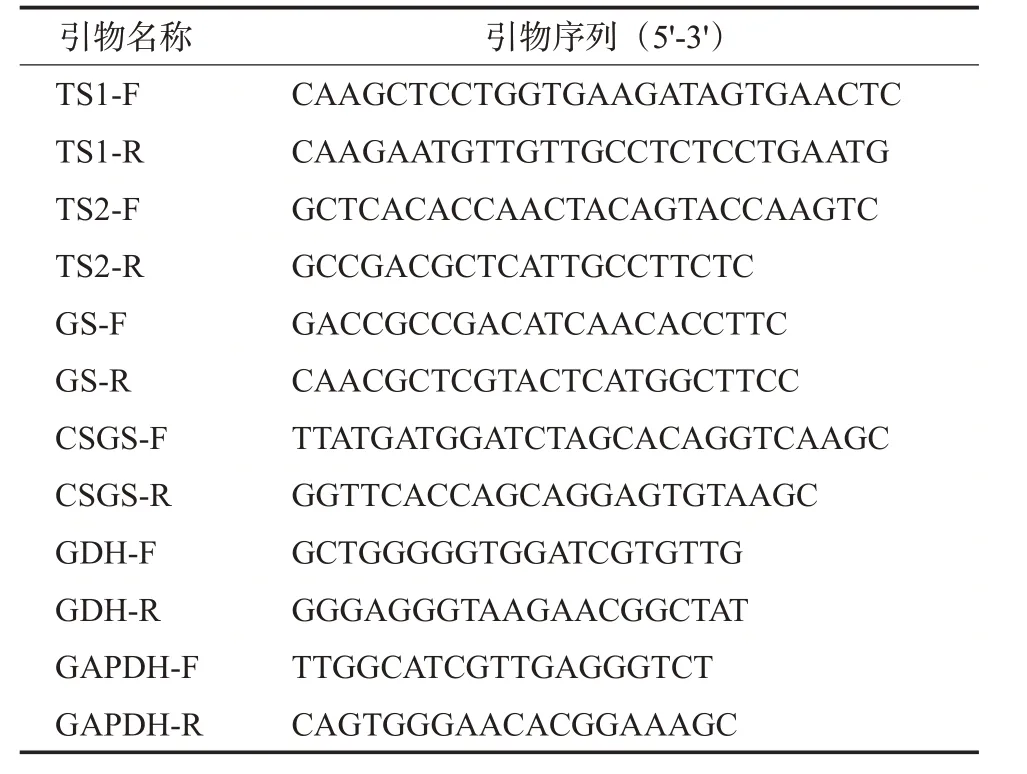
表1 相关酶基因引物序列
选择茶树GADPH作为内参基因,每个对照基因样品和内参基因样品都设置3个重复及1个对照(阴性),观察溶解曲线及扩增曲线,相对表达量计算采用2-ΔΔCT法。
1.4 数据分析
进行多组样本间差异显著性分析。
2 结果与分析
2.1 外源水杨酸处理对铁观音幼苗光合性能的影响
不同浓度的外源水杨酸处理对铁观音幼苗叶绿素、可溶性糖和蛋白质含量均有不同程度影响。由表2可见,铁观音幼苗的叶绿素含量随着外源水杨酸浓度的提高呈现先增后降的趋势,在水杨酸浓度为3.0 mmol/L时达到最大值,且与对照差异显著,比对照增加30.28%;铁观音幼苗的蛋白质含量在喷施外源水杨酸后均有提高,其中水杨酸浓度1.0~3.0 mmol/L处理的蛋白质含量显著高于对照,但处理间差异不显著;可溶性糖含量当水杨酸

表2 外源水杨酸处理对铁观音幼苗光合性能的影响mg/g
浓度3.0 mmol/L时达到最大值,且与对照差异显著,比对照增加49.74%,水杨酸浓度4.0 mmol/L时可溶性糖含量略微下降,但仍显著高于对照。
2.2 外源水杨酸处理对铁观音幼苗抗氧化特性的影响
POD、MDA和脯氨酸为植物抗氧化特性的主要指标。由表3可见,随着外源水杨酸浓度的升高,铁观音幼苗POD活性表现出先升高后降低的趋势,水杨酸浓度为3.0 mmol/L时POD活性达到最大值,与对照差异显著,比对照增加69.29%;MDA含量在水杨酸浓度为2.0~4.0 mmol/L时显著低于对照,并随着水杨酸浓度的提高而下降;脯氨酸含量在喷施外源水杨酸后均有不同程度下降,但只有处理3与对照差异显著。
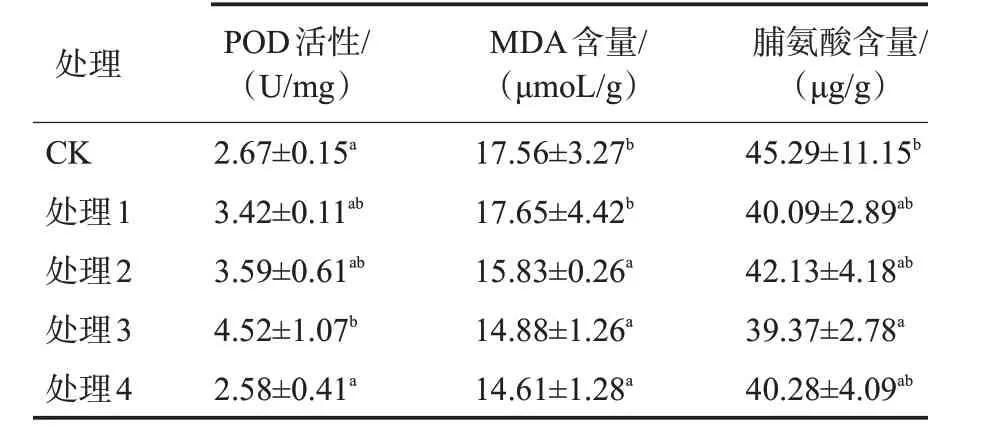
表3 外源水杨酸处理对铁观音幼苗抗氧化特性的影响
2.3 外源水杨酸处理对铁观音幼苗茶氨酸含量的影响
茶氨酸是茶树体内特有的氨基酸,不仅是茶的主要呈味物质,也是其主要功能成分,可以改善人体免疫功能,提高抵抗力等。经外源水杨酸处理后,茶氨酸含量随着水杨酸浓度的提高出现先升高后降低的现象(图1);水杨酸浓度为2.0 mmol/L和3.0 mmol/L时,茶氨酸含量显著高于对照;水杨酸浓度为4.0 mmol/L时茶氨酸含量降低,且显著低于对照。由此可见,水杨酸对茶氨酸的生成具有浓度效应,低于3.0 mmol/L时促进茶氨酸生成,高于3.0 mmol/L时则抑制茶氨酸生成。
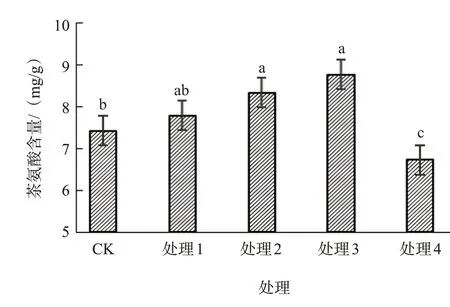
图1 水杨酸浓度对铁观音幼苗茶氨酸含量的影响
2.4 外源水杨酸处理对铁观音幼苗茶氨酸代谢途径相关酶基因表达的影响
茶氨酸合成代谢途径基因包括直接参与合成的茶氨酸合成酶基因和茶氨酸水解酶基因,还包括谷氨酸合成相关的基因及茶氨酸转运相关基因。谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)、谷氨酸合成酶(CSGS)和茶氨酸合成酶(TS)是茶氨酸代谢途径中的关键酶。由图2可见,水杨酸处理的GDH和TS1的基因表达量均高于对照,且随着水杨酸浓度增加表现出先升高后降低的趋势;当水杨酸浓度为3.0 mmol/L时GDH和TS1的基因表达量均达到最大值。CSGS基因表达量随着水杨酸浓度的增加而逐渐降低,当水杨酸浓度为1.0 mmol/L时基因表达量显著高于对照,当水杨酸浓度为2.0~3.0 mmol/L时基因表达量与对照差异不显著,当水杨酸浓度为4.0 mmol/L时基因表达量显著低于对照。
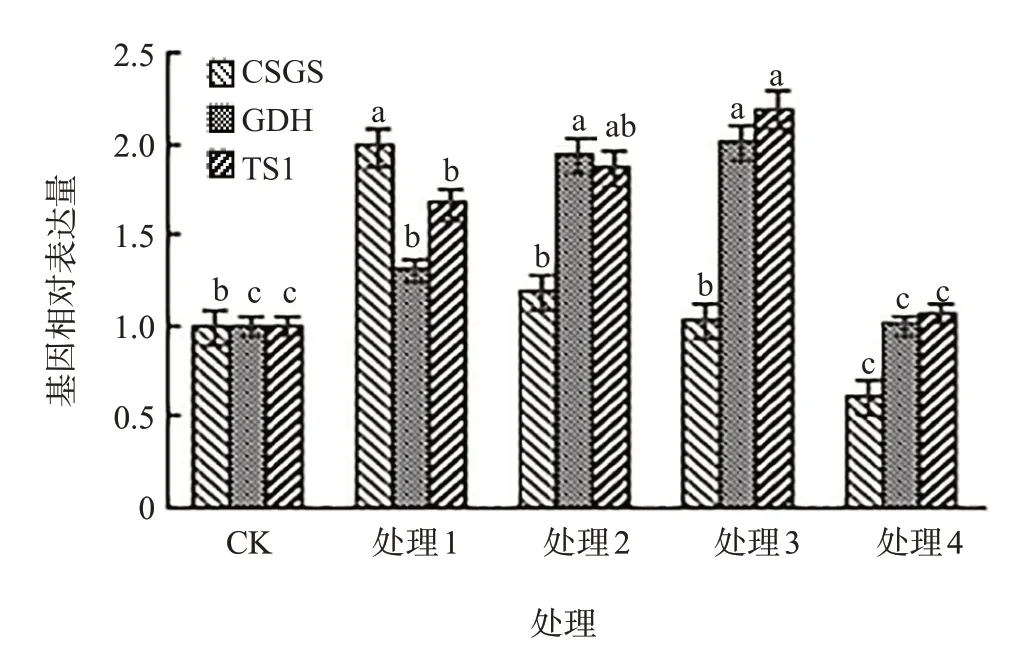
图2 不同水杨酸浓度处理对CSGS、GDH、TS1表达量的影响
水杨酸处理的GS和TS2的基因表达量均高于对照,且随着水杨酸浓度增加表现出先升高后降低的趋势;当水杨酸浓度为2.0 mmol/L、3.0 mmol/L时GS和TS2的基因表达量均显著高于对照,且当水杨酸浓度为3.0 mmol/L时GS和TS2的基因表达量均达到最高值(图3)。
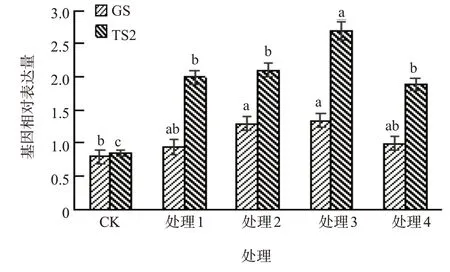
图3 不同水杨酸浓度处理对GS、TS2表达量的影响
3 小结与讨论
水杨酸在植物代谢生理中起着重要的作用,能够促进植物细胞分裂和伸长,降低膜脂过氧化作用,提高植物体内抗氧化酶活性,以缓解环境胁迫对植物造成的伤害[19]。水杨酸可以通过提高植物的抗氧化活性对植物体内的信号通道产生影响[20]。前人研究表明,半夏植株喷施0.5 mmol/L水杨酸后,叶绿素含量和块茎鲜质量显著增加,植株MDA含量降低,SOD保护酶活性提高[21]。甜樱桃采收期喷施0.5 mmol/L水杨酸,果实的色泽、硬度、可溶性固形物、总酚酸含量,以及保护酶活性显著提高[22]。水杨酸虽能促进细胞的生长,但高浓度的水杨酸处理或内源水杨酸的积累会对细胞生长产生氧化胁迫伤害,表现出抑制作用[23]。本试验中喷施3.0 mmol/L水杨酸处理的铁观音幼苗叶绿素、可溶性糖和蛋白质含量均显著增加,脯氨酸和MDA含量显著降低,POD活性显著提高。而较高浓度的水杨酸明显抑制铁观音幼苗的生长,表现出轻微的毒害作用。这可能是由于叶面喷施水杨酸后,植物体内防御机制被启动,导致参与防御反应的相关酶活性升高。
水杨酸是植物抗逆的内源信号物,可诱导植物体内某些次生代谢产物的生物合成[24-26]。本研究中喷施3.0 mmol/L水杨酸对茶树茶氨酸含量的增加具有促进作用,而较高浓度的水杨酸明显抑制茶氨酸生物生成。
茶氨酸的生物合成也受到茶氨酸合成代谢途径基因表达的调控[27]。林郑和等[28]比较茶氨酸含量不同的茶树种质,发现在高茶氨酸种质中GDH基因表达显著增强。Wang等[29]发现干旱导致茶氨酸含量降低,其中GS、CSGS等基因表达下降,同时茶氨酸水解酶基因表达水平升高。本研究中,GDH和TS1对茶氨酸合成代谢起到主要作用,适宜浓度的外源水杨酸有利于提高茶氨酸的积累。目前,虽然部分参与茶氨酸合成代谢的关键酶基因已被克隆,但对于该代谢途径的转录调控网络还不清楚;外源信号物对茶氨酸代谢调控机理有待进一步研究。
