平潭岛台湾相思群落优势种群生态位研究
2023-01-15马晓迪姜德刚刘子琳温瑞龙邓传远
马晓迪,姜德刚,刘子琳,王 芳,温瑞龙,邓传远*
平潭岛台湾相思群落优势种群生态位研究
马晓迪1,姜德刚2,刘子琳1,王 芳1,温瑞龙1,邓传远1*
1. 福建农林大学园林学院,福建福州 350002;2. 自然资源部海岛研究中心,福建福州 350400
台湾相思()是平潭海岛相思林的主要建群种,是海岛优良的造林阔叶树种,了解平潭岛台湾相思群落主要木本植物和草本植物种群的生态位特征有助于深入了解海岛群落物种共存与竞争机制。在对其内岛和附属岛屿进行群落调查的基础上,利用Levins和Shannon生态位宽度和Pianka生态位重叠指数分析平潭岛台湾相思群落各层片主要种群生态位。共调查到维管束植物144种隶属63科123属,以禾本科和菊科为主,与大陆植被存在显著差异;发现入侵风险植物14种,草本入侵现象较严重,其中1级恶性入侵的假臭草()、鬼针草()和2级严重入侵的香丝草()在群落中占据较大的生态位,可能对共生种群造成一定的生态干扰,应当引起足够重视;乔木层中的台湾相思和木麻黄()、灌木层中台湾相思幼树和雀梅藤()、草本层中芒()和假臭草生态位宽度较大,生态位宽度小于0.5的物种数占总数的96.67%,重要值较大的物种生态位宽度也较大,但并非完全一致;优势种群生态位重叠表现为草本层(0.253)>灌木层(0.231)>乔木层(0.149),均小于0.3,生态位重叠值普遍偏低,种群通过生态位分化来减弱种间竞争。海岛地区较强的环境过滤性导致种群产生不同程度的生境分化,同时,种群通过压缩生态位宽度,聚集在局部生境中,从而促进海岛群落物种共存,平潭岛台湾相思群落属于地带性顶级群落,种间关系稳定,种群更新缓慢。
平潭岛;台湾相思;生态位宽度;生态位重叠
生态位概念最早是由GRINNELL[1]提出,用于描述群落中不同种群对资源的利用程度及其与相关种群之间的功能关系,从而体现群落中各个种群的重要程度及其功能和地位。生态位研究一直以来都是种群生态学研究的核心问题,主要包括生态位宽度和生态位重叠度,量化物种群落对环境资源的支配能力和重叠利用度,广泛应用于种间关系[2-3]、群落特征[4-5]、群落演替[6-7]、物种多样性保护[8-9]等方面的量化研究,是用于科学解释群落间物种共存与竞争机制的理论之一。海岛地理环境特殊、规模有限,使得海岛生态系统具有明显的独立性与资源短缺性,稳定性较低,自我调节能力和恢复能力弱。植被是海岛生态系统的结构基础,对于维持生态系统稳定具有重要作用,因此,量化海岛物种之间、物种与环境之间的相互关系,有助于更清晰直观地了解海岛群落内物种对资源的利用状况,对种群的保护具有重要意义。
台湾相思隶属于豆科(Leguminosae)金合欢属()常绿乔木,因其耐干旱瘠薄,广泛分布于我国热带及亚热带海岛冲刷严重地区,是海岛地区水土保持和海岛防护林重要树种[10]。台湾相思群落是平潭岛分布面积最广的群落类型之一,主要分布于山地低海拔地区,在调节气候、固碳释氧、涵养水源以及沿海防护方面具有重要作用。目前关于相思树的研究主要集中在群落演替及物种多样性[10]、生物学特性[11]、功能性状[12]方面。关于平潭岛台湾相思群落的生态学研究尚处在探索阶段,其群落发展状况及其与其他相关种群间的关系尚不明晰。本研究运用生态位理论研究台湾相思群落,对于认识其群落物种共存规律对该地区的生态维持和受损生态系统恢复具有重要意义,以期揭示台湾相思群落结构与动态,从而优化海岛植物群落配置、提高群落稳定性,为海岛防护林植被恢复与生物多样性保护提供科学理论依据。
1 材料与方法
1.1 调查区域自然概况
平潭岛作为全国第五大岛,福建第一大岛,其面积为267.13 km2。地处中国东南部沿海地区(25°15′~25°45′N,119°32′~120°10′E),位于平潭综合实验区内,东临台湾岛。以半湿润海洋性季风气候为主,年平均风速为5.0 m/s,大风极值出现在夏季,夏季盛行西南风,冬季盛行东北风,受台风影响巨大[13]。平潭岛地形为丘陵台地平原岛屿区,主要土壤类型包括红壤、滨海风沙土和盐土,年平均气温在19~19.9℃,全年气候温和[14]。岛上森林类型主要分为针叶林、针阔混交林和阔叶林。其中常绿阔叶林以台湾相思林为主,植被结构较为单一,优势种明显,乔木层建群种包括台湾相思、木麻黄,灌木层包括算盘子()、福建胡颓子()和茅莓(),草本层主要有弓果黍()、假臭草和芒。
1.2 野外群落调查
分别于2017年9月至2018年11月期间对平潭岛主岛及附属岛屿进行调查,根据平潭岛实地走访调查,选取平潭主岛的白犬山、瑞玲山、东屿;附属海岛调查包括大嵩岛、北香炉屿、黄门岛共包括21个20 m×20 m的样方,样方布设遵循均匀性,可达性,合理性(图1)。其中,北香炉屿和东屿分别为4、5个,其余均为3个,详情见表1。采用标准样方法进行样方设置,森林样方取样单元为20 m×20 m,对样方内树高≥3.0 m或者树胸径≥2.5 cm的乔木进行记录;每个样方四角设置4个10 m×10 m灌木小样方(包括胸径<2.5 cm的乔木幼树),记录乔灌样方内所有个体的种名、数量、高度和胸径(基径),于大样方内四角及中心位置设置5个1 m×1 m小样方进行草本调查,记录其高度、冠幅、株数、盖度等数据[6]。样方地地形因子使用GPS记录经纬度及海拔,森林罗盘仪记录坡度、坡向、坡位。样方内无法鉴定的植物采集标本,拍摄并做好标记带回实验室,邀请海岛专家及参考《福建植物志》[15]、《中国植物志》[16]资料鉴定。

图1 研究区位置及样方布设
1.3 数据处理
1.3.1 重要值 各层物种的重要值(IV)计算方法如下[17]:
乔木层、灌木层IV=(RA+RH+RD)/3
草本层IV=(RD+RH)/2
式中,RA为相对多度,RH为相对高度,RD为相对优势度。
1.3.2 生态位 分别选取平潭岛台湾相思群落乔木层、灌木层和草本层中重要值靠前且频度较大的8种、15种和11种主要物种进行生态位分析(乔灌层部分重叠),利用Levins指数(B)和Shannon-Wiener(B)[18]指数测定种群的生态位宽度,Pianka指数(O)[19]计算种群的生态位重叠。


表1 平潭岛台湾相思群落21个样方的基本情况
注:E. 东坡;S. 南坡;W. 西坡;N. 北坡;ES. 东南坡;WS. 西南坡;WN. 西北坡;Us. 上坡;Ms. 中坡;Ds. 下坡;Ws. 迎风坡;Ls. 背风坡。
Note: E. East Slope; S. South slope; W. West slope; N. North slope; ES. Southeast slope; WS. Southwest slope; WN. Northwest slope; Us. Uphill; Ms. Medium slope; Ds. Downhill; Ws. Windward slope; Ls. Leeward slope.
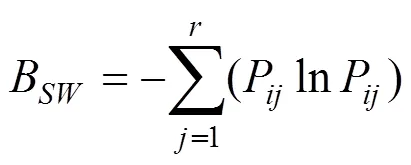
式中,P=n/N,P为物种在样方中的重要值占所有样方重要值总和的比例,为样方数。B具有域值[0,1],当值为0时,生态位宽度最窄,当值为1时,生态位最宽。
式中,O为种群和的生态位重叠值,P和P分别表示第和第物种在第个资源水平下的重要值占该种在所有资源水平上重要值总和的比例,为样方数。
2 结果与分析
2.1 台湾相思群落物种组成
在所调查的21个样方中,共发现维管束植物144种隶属63科123属,其中乔木共有植物16科17属18种,灌木层有植物28科40属47种(乔灌有重复),草本层有植物32科65属73种,层间植物22种。其中层间植物出现频度较大的包括鸡矢藤(, 81.82%)、千金藤(, 77.27%)、海金沙(, 63.64%)和忍冬(, 54.55%)。研究区植物属种系数为84.72%。其中以被子植物为主,共计50科110属127种,蕨类植物12科12属15种,裸子植物仅1科1属2种。依据国家环境保护总局发布的入侵物种名单、《中国入侵植物名录》等确定外来入侵植物和入侵等级[20-21]。研究区共发现入侵植物14种,其中1级恶性入侵植物5种,包括鬼针草、假臭草、马缨丹()、土荆芥()、小蓬草(),2级严重入侵植物5种包括铺地黍()、牵牛()、仙人掌()、香丝草、银合欢()。科组成上,以禾本科()和菊科()为主,仅含1种的科有34个,占总科数的53.97%,如樟科()、胡颓子科()、忍冬科()等;属组成上,优势属不明显,以仅含1种的属(105个)优势显著,占总属数的85.37%,主要有梵天花属()、菝葜属()、荛花属()等;含2~3种的属共18个,占总属数14.63%,如酢浆草属()、榕属()、南蛇藤属()等;由此可见,研究区的科属组成复杂多样,且草本层入侵植物较多。
2.2 台湾相思群落主要种群重要值特征
在所调查的21个样方中,乔木层有植物18种,平均重要值排序前8位(表2)的是台湾相思(74.06%)>木麻黄(10.07%)>朴树(, 2.47%)>银合欢(2.07%)>海桐(, 1.41%)>楝(, 1.06%)>天仙果(., 1.01%)>滨柃(, 0.95%),其中台湾相思最大值为100,样方中含有台湾相思纯林。灌木层有植物47种,平均重要值排序前15位(表3)为台湾相思(27.09%)>雀梅藤(7.50%)>滨柃(6.01%)>车桑子(, 5.63%)>木麻黄(5.23%)>福建胡颓子(4.03%)>茅莓(3.83%)>九里香(, 3.55%)>算盘子(3.31%)>栀子(, 2.98%)>铁包金(, 2.13%)=球花脚骨脆(, 2.13%)>构棘(, 1.96%)>海桐(1.95%)>黑面神(, 1.81%),其中台湾相思平均重要值占一定优势,其余灌木较为均衡。草本层有植物73种,挑选平均重要值前11位(表4)分别为芒(12.32%)>假臭草(8.47%)>狗尾草(, 2.58%)>求米草(, 6.45%)>积雪草(, 4.87%)>弓果黍(9.29%)>火炭母(, 3.56%)>香丝草(3.00%)>鬼针草(3.87%)>山麦冬(, 2.19%)>酢浆草(, 2.82%)。
2.3 生态位宽度
由表2可知,平潭岛台湾相思群落乔木层种群的生态位宽度差异明显,乔木层中台湾相思和木麻黄的重要值分别为74.06%和10.07%,生态位宽度也相对较高,其B和B分别为0.93、3.00和0.30、1.95,对台湾相思群落起着决定性作用,环境适应能力也较强。台湾相思的生态位宽度显著最大,银合欢、朴树和海桐虽然重要值排序相对靠前,但分布样地少,导致生态位宽度较小。由表3可知,灌木层中生态位宽度最大的是台湾相思幼树,其B和B分别为0.59和2.70,其次为雀梅藤,其B和B值分别0.49和2.50,生态位宽度也存在一定差异。生态位宽度最小的是构棘,其B和B值分别为0.11和1.15,多数物种B和B分别处于0.1~0.3和1~2之间。由表4可知,草本层生态位宽度差异较小,生态位宽度最大的是芒,最小的是鬼针草,i和B多位于0.20~0.40和1.5~2.5之间。整体来看,物种的重要值与生态位宽度排序并非完全一致,Levins和Shannon-Wiener生态位宽度公式计算出二者结果的排序基本趋于一致。台湾相思群落乔木层和灌木层存在重复,但是生态位宽度却存在差异,台湾相思在乔木层和灌木层均占据较大生态位宽度,但是灌木层其B和B分别为0.59和2.70,小于乔木层。木麻黄在乔木层生态位宽度位于第二,但是在灌木层中分布宽度不占据优势,位居灌木层生态位宽度的11位。

表2 台湾相思群落乔木层种群的重要值和生态位宽度
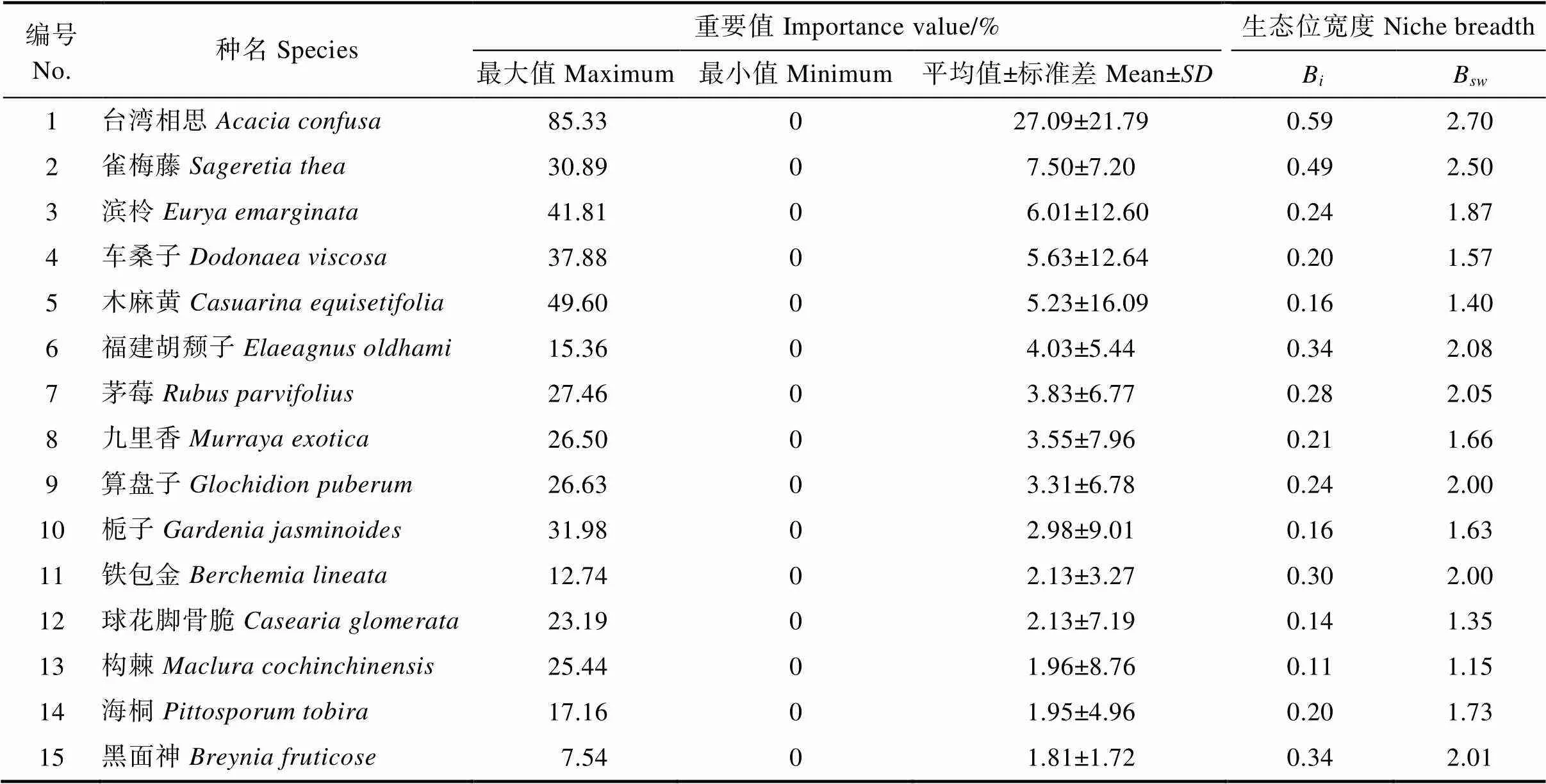
表3 台湾相思群落灌木层种群的重要值和生态位宽度

表4 台湾相思群落草本层种群的重要值和生态位宽度
2.4 生态位重叠
由表5可以看出,平潭岛台湾相思群落乔木层8个物种共组成28个种对,重叠值最大的是楝与滨柃(0.500),其中有24个种对Pianka生态位重叠值小于0.5,占总数的85.71%,生态位重叠值大于等于0.4的种对有4对,占总数的14.29%,不发生重叠的种对有12对,占总数的42.86%。由表6可知,群落灌木层15个优势种共组成105个种对,构棘与球花脚骨脆重叠值最大为0.863,Pianka生态位重叠指数O>0的有97对,占总数的92.38%,其中O≥0.5的种对有8对,占总数的7.62%,O<0.5的97对,占总数的92.38%;无重叠种对有8对,占总数的7.62%。由表7可知,草本层11个物种共组成55个种对,其中弓假臭草重叠值最大(0.690),有6对生态位重叠果黍与值≥0.5,占总数的10.91%。
在资源轴上(图2),台湾相思群落乔木层优势种间有13对种对生态位重叠值在0~0.1之间,占总种对的46%;灌木层生态位重叠值在0~0.3之间有76对,占总对数的72%;草本层位于0.2~0.3之间的种对最多,有18对,占总数的33%。可以看出,生态位重叠值大于0.6的种对明显减少,且没有种对间生态位重叠值位于0.9~1之间。结果表明,生态位宽度较大的种群,与其他种群产生重叠的可能性较大,如乔木层生态位宽度较大的台湾相思与木麻黄,与同层片的种群均产生重叠,且能与生态位较窄的物种也发生重叠,可见这2个种群在海岛上分布较广,环境资源利用较强,充分适应海岛恶劣环境。

表5 台湾相思群落乔木层种群生态位重叠
注:物种编号同表2。
Note: Species number saw Tab. 2.

表6 台湾相思群落灌木层种群生态位重叠
注:物种编号同表3。
Note: Species number saw Tab. 3.

表7 台湾相思群落草本层种群生态位重叠
注:物种编号同表4。
Note: Species number saw Tab. 4.

图2 种间生态位重叠值分布特征
3 讨论
3.1 群落物种组成和入侵植物生态位特征
本研究共发现维管束植物144种隶属63科123属,其中物种的丰富度指数表现为草本层(10.76)>灌木层(10.29)>乔木层(2.52)。禾本科和菊科是该植物区系的优势科,植被优势科属与陆地存在显著差异,樟科和壳斗科等亚热带陆地阔叶林优势树种,在海岛上却少有分布,这与菊科和禾本科繁殖能力及其功能性状适应海岛生境有关,表明平潭岛植物并非以亲缘关系群居,而趋于在适宜生境中群居[22]。受外来活动影响,岛屿具有入侵趋势的外来物种增多,威胁当地生物多样性[23]。本研究区中调查发现有14种植物构成入侵危害,以菊科、禾本科和豆科居多。草本入侵现象最为严重,其中1级恶性入侵的鬼针草、假臭草和2级严重入侵的香丝草在台湾相思群落草本层中占据优势地位。根据生态位计算结果显示,三者的Levins和Shannon-Wiener生态位宽度分别为0.40,2.20、0.23,1.62、0.23,1.70,假臭草重要值和生态位宽度均靠前,在群落中入侵现象最严重。三者均为菊科一年生草本,菊科植物生长周期短、繁殖速度快、植株成熟早都成为成功定居群落且占据平潭岛台湾相思群落草本层主要地位的关键[24],海岛入侵植物对共生种群造成一定的生态干扰,应当引起足够重视。
从生态位角度分析,物种成功入侵主要是由于恶劣的环境条件、低植被覆盖率、频繁的干扰和空白生态位[25]。海岛生态环境恶劣,植物都聚集在局部适宜生境中,导致生态系统处于非饱和状态,具有空缺生态位可以容纳更多种群。假臭草占据较大生态位宽度,但是与其他草本的生态位重叠都较小,其在资源有限的条件下充分利用空生态位,在资源需求上产生分化,进而在某种程度上回避了种群间对资源的竞争,这可能是假臭草能够在海岛上与定居物种共存的主要原因。引入外来植物虽暂时的丰富了当地生物多样性水平,但由于缺乏科学管理,其对多样性水平维持和生态系统结构功能造成严重威胁。假臭草入侵后便零散分布于平潭各种生境,强大的自繁能力使其生长过程中极大消耗土壤肥力,伴随有植物间的化感作用,长久下来会形成单优势种群从而危及当地草本植物生长[26]。鬼针草在入侵过程中虽然没有占据较大的生态位宽度,但与多种草本具有相似的生境偏好,在资源利用上是竞争关系,如芒(0.51)和狗尾草(0.62)。鬼针草种子具有倒刺毛,能靠风力传播和动物传播在短时间内大量繁殖,比原有生物更具竞争优势,如果不及时防治,可能会威胁相似生境下的植物生长。
3.2 群落主要植物生态位宽度特征
重要值和生态位是衡量群落中物种地位和作用的重要指标,重要值侧重于反映物种优势度,是影响生态位宽度的重要因素之一,研究表明,重要值越大,生态位宽度也越大。郑俊鸣等[27]通过回归分析发现东山岛木麻黄和花吊丝竹混交林中物种生态位宽度和重要值具有显著正相关。但也存在不完全一致的情况,魏凯等[7]研究光幼屿典型群落生态位,发现黑松和木麻黄虽然重要值大,但是生态位宽度却偏小,表明重要值和生态位宽度存在不一致的情况。本研究中重要值和生态位宽度也并非完全一致,例如台湾相思群落中灌木层木麻黄重要值位列第五,但生态位宽度却排在较后,而海桐和黑面神重要值较小,生态位宽度位居中等水平。草本层的狗尾草重要值较小,但其生态位宽度却位于前列,相反,鬼针草重要值较大,生态位宽度却最小。出现这种情况可能是因为重要值并不是唯一影响因素,分布频度、种间关系也同样影响着生态位宽度,一般频度越大,生态位宽度就越大。本研究中台湾相思(100%)、雀梅藤(54.76%)、弓果黍(29.52%)、和芒(22.86%)等在资源位中出现的次数多,频度较大,分布范围较广,所以它们的生态位宽度较大,对资源的利用能力和环境适应能力较强。
生态位宽度可以衡量物种对环境资源利用能力和分布状况,生态位宽度越大,其对生境中环境资源利用能力就越强,在群落中分布越广,占据越重要的地位[5]。刘润红等[6]研究枫杨群落发现,建群种在营造群落内部特殊生境和决定群落内物种构成等方面起主导作用,在所有资源位上都有分布且数量多,生态位宽度要比伴生种宽。尼玛曲珍等[28]研究川滇高山栎群落同样发现建群种在群落中占据主导地位,即使与其他物种混生,也在立地中也具有一定优势。台湾相思群落中生态位宽度较大的物种有台湾相思和木麻黄,灌木层中有台湾相思幼树、雀梅藤和福建胡颓子,草本层生态位宽度较大为芒、假臭草和积雪草,说明这些植物在平潭岛中生态幅较宽,具有较强适应能力和资源利用能力,对平潭岛群落构建起到重要作用。灌木层构棘和球花脚骨脆数量较少,且分布相对集中,所以生态位宽度较小,在群落中地位较低,可能由于他们对环境变化非常敏感,分布只能局限在特定的区域。木麻黄在乔木层中的生态位宽度显著大于灌木层,可能因为木麻黄是强阳性树种,在灌木层中无法获得充足阳光,造成林下幼树竞争力弱,分布范围受到限制。本研究发现,台湾相思建群种因其极耐干旱和瘠薄,在乔木层和灌木层中重要值、频度和生态位宽度都较大,广泛分布于资源位上,对平潭岛环境的适应能力和资源利用能力都较强,在发展中逐渐形成单优势种群落,对海岛群落环境发挥着决定性作用。因此,台湾相思可以作为海岛防护林优良的生态树种之一,在平潭岛上充分发挥群落的水土保持和防护的作用。
3.3 群落主要植物生态位重叠特征
生态位重叠反映2个物种利用同等资源的相互重叠情况,看出2个物种间生态学特性相似程度和对资源利用相似程度,从而反映物种间竞争关系[29]。研究区各层片之间生态位重叠值普遍偏低,低于0.4的种对占据总种对的82.45%,表明优势种间对环境利用存在一定程度分化,资源共享充分,种间竞争较弱,整体处于稳定状态。平潭岛植物演替方向为灌草丛—针叶林—针阔混交林—阔叶林[30]。目前,研究区台湾相思群落由针阔混交林阶段过度到较为稳定的阔叶林阶段,所以各物种间生态位重叠程度变小,种间竞争相较之前有所降低,各物种间达到一种相对平衡,在同一环境内稳定共生。这符合孙成等[31]和刘润红等[32]认为群落到演替后期形成地带性顶级群落,生态位分化越合理,群落进入相对稳定状态,说明平潭岛植被演替中也具有相似的规律。整体来看,台湾相思群落发生重叠的种对有168对,占总数的89.36%,重叠率较高。共有20对物种间不发生重叠,说明台湾相思群落生态位分化较明显,这可能与平潭岛土层浅,土壤含水量低等理化性质和植物的生态习性差异导致的生境异质性有关。例如车桑子和球花脚骨脆、福建胡颓子和栀子,从实地调查来看,球花脚骨脆多生长于阔叶疏林中,而车桑子则喜强光并耐干旱,常生长于干旱山坡及海边的沙土中;福建胡颓子和栀子也同样存在生态习性的差异,福建胡颓子常分布于林下阳光充足的疏林中或者受强海风吹袭的迎风坡,而栀子不耐阳光直射且是浅根性植物,因此常生于荫蔽条件好的密林下,且多分布于背风坡。上述二者具有不同的生态习性,在环境利用上存在一定的分化,该分化在一定程度上保护了平潭岛物种多样性。
研究表明,物种的生态位宽度越大,与其他种群发生重叠的几率也随之上升[33]。但是物种由于自身的生态学和生物学特性,即使生态位宽度大的物种对资源的要求也有所不同。本研究发现,生态位宽度较小的种群也存在较大的生态位重叠,例如,球花脚骨脆和海桐、球花脚骨脆和构棘、狗尾草和鬼针草,生态位重叠值分别为0.576、0.863、0.620。相反生态位宽度较大的种群,如木麻黄和朴树、木麻黄幼苗和车桑子、台湾相思幼苗和车桑子,重叠值仅为0.032、0.002、0.192。优势种群生态位重叠表现为草本层(0.253)>灌木层(0.231)>乔木层(0.149),由于灌木层和草本层物种生态习性趋于一致且物种丰富度明显高于乔木层,因此对资源的竞争程度大于乔木层,并且平潭岛受外来影响较大,灌草层更容易受到人为干扰,因此重叠较大。有学者认为生态位重叠并不一定会导致竞争,只能说明二者占据了相似的生态空间,在环境压力和种间相互适应作用下协同进化,物种通过不同途径利用资源,使物种与环境和谐发展[34]。海岛环境封闭,受台风、海浪等因素的影响,各种资源匮乏,使物种之间更加倾向于分享基础生态位,因此某些种群生态位宽度压缩,聚集在局部生境中。本研究结果中生态位宽度小于0.5的物种数占总数的96.67%,验证了物种通过压缩生态位宽度聚集在适宜生境这一结果。
3.4 海岛植物多样性保护及维护措施
面对岛屿生物多样性减少丧失,岛屿生态系统具有一定的自我修复功能,尽管生物多样性的丧失显著减少了植物生物量,但这种影响可以通过剩余物种的补偿反应来缓解,而这种缓解会随着时间的推移而加强[35]。这表明,如果其他物种能够占据新出现的生态位空间,那么生态系统对物种的损失具有恢复能力。从入侵植物角度分析,要充分保护海岛原生植物多样性,可筛选功能性状或生态位特征相近的本地物种进行替代种植。例如假臭草喜光,在海岛路边、裸地及林边光照条件好的地方优势度非常大,可以利用具有经济价值和生态价值的乡土植物,如木豆、车桑子、算盘子和芒等在海岛上快速生长、形成密丛的特点,阻止阳生性草本大量繁殖,也可利用弓果黍和狗牙根进行替代种植,弓果黍为耐阴草本,常与假臭草在同一生境中出现,对光照的不同需求减少了两者间的竞争,可以和谐共生在相思林下,狗牙根与假臭草生存环境相似,在同等资源下竞争强烈,可以有效减少假臭草密集生长;鬼针草入侵对海岛生态系统造成巨大危害,可以利用生存环境相似的芒和狗尾草来增加资源竞争或引入寄生植物南方菟丝子共同种植,防止鬼针草持续形成优势群落;香丝草防控则可选用求米草进行替代种植。
从植物配置角度分析,应优先配置生态位宽度大的灌草植物,将生态位特征相似的物种进行搭配种植,集中保护,确保生物多样性不降低,使资源得到充分利用。茅莓、铁包金和福建胡颓子对生境的要求相似,常生于向阳山坡和阳光较好的路旁空旷地带,可将三者搭配种植作为边坡道路绿化,既富有野趣又可抑制路旁阳性杂草蔓延;海桐和构棘、球花脚骨脆、黑面神重叠较大且均为革质叶,叶片表面光滑,角质层较厚,对海岛环境适应性强,可以种植在林缘形成防风灌丛景观,具有观赏性并保护林内幼苗及下层植被生长;积雪草、弓果黍、求米草、酢浆草和火炭母常于较为阴湿环境中密集存在,在海岛上可作为林下地被植物成片种植保护,不仅可以丰富海岛植物层次还可以抑制入侵植物肆意生长。乔木中的木麻黄、黑松、朴树、楝树和灌木中的九里香、滨柃、车桑子、仙人掌和夹竹桃都适应滨海沙地恶劣环境,可种植在近海岸沙地作为海岸防护林中上层植被,具有海岛特色的同时还能够修复近海岸脆弱的生态系统。
4 结论
本研究分析了平潭岛台湾相思群落的生态位特征,发现台湾相思在群落中地位高,分布范围广,能与大量生态位较窄的物种发生重叠,对海岛环境资源利用充分且具备较强适应能力。海岛地区较强的环境过滤性导致物种生境分化和生态位重叠增加在群落物种共存中发挥重要作用。可以看出群落植被生态位宽度与生态位重叠指数普遍较小,虽整体处于稳定发展状态,但都聚集在局部适宜生境中,群落物种对资源环境的利用程度不大,物种多样性有待提高。因此,在对平潭岛台湾相思群落进行植被修复和生态保护的实践过程中,积极防控入侵植物,避免入侵物种与当地物种产生过多的资源竞争。充分考虑群落内各种群的生态学特征,可以将生态位特征相似的物种进行搭配种植,成片保护,对重叠值较大的物种尽量成对保护,优化植被配置的同时,利用物种在资源位上的差异,筛选适合物种弥补空缺生态位,从而在海岛有限的资源下充分实现资源互补利用,增加海岛物种多样性,保持群落稳定发展。
[1] GRINNELL J. The niche-relationships of the California Thrasher[J]. Auk, 1917, 34(4): 427-433.
[2] HART S P, TURCOTTE M M, LEVINE J M. Effects of rapid evolution on species coexistence.[J]. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(6): 2112.
[3] 陈佳卉, 卜元坤, 苏少峰, 黄 维, 陈周娟, 李卫忠. 油松飞播林灌木层主要物种种间联结及其环境解释[J]. 生态学杂志, 2021, 40(11): 3512-3522.
CHEN J H, BU Y K, SU S F, HUANG W, CHEN Z J, LI W Z. Interspecific association and environmental interpretation of main species in shrub layer ofaerial seeding forest[J]. Chinese Journal of Ecology, 2021, 40(11): 3512-3522. (in Chinese)
[4] LI Y Z, SHIPLEY B, PRICE J N, DANTASS V DE L, TAMME R, WESTOBY M, SIEFERT A, SCHAMP B S, SPASOJEVIC M J, JUNG V, LAUGHLIN D C, RICHARDSON S J, BAGOUSSE-PINGUET Y L, SCHÖB C, GAZOL A, PRENTICE H C, GROSS N, OVERTON J, CIANCIARUSO M V, LOUAULT F, KAMIYAMA C, NAKASHIZUKA T, HIKOSAKA K, SASAKI T, KATABUCHI M, DUSSAULT C F, GAUCHERAND S, CHEN N, VANDEWALLE M, BATALHA M A. Habitat filtering determines the functional niche occupancy of plant communities worldwide[J]. Journal of Ecology, 2018, 106(3): 1001-1009.
[5] 董 雪, 李永华, 张正国, 李思瑶, 包岩峰, 郝玉光, 姚 斌. 甘肃酒泉荒漠戈壁灌木群落优势物种生态位特征[J]. 中国沙漠, 2020, 40(4): 138-145.
DONG X, LI Y H, ZHANG Z G, LI S Y, BAO Y F, HAO Y G, YAO B, Niche of dominant shrub species in desert gobi in Jiuquan, Gansu, China[J]. Journal of Desert Research, 2020, 40(4): 138-145. (in Chinese)
[6] 刘润红, 常 斌, 荣春艳, 姜 勇, 杨瑞岸, 刘星童, 曾惠帆, 傅桂焕. 漓江河岸带枫杨群落主要木本植物种群生态位[J]. 应用生态学报, 2018, 29(12): 3917-3926.
LIU R H, CHANG B, RONG C Y, JIANG Y, YANG R A, LIU X T, ZENG H F, FU J H. Niche of main woody plant populations ofcommunity in riparian zone of Lijiang River, China[J]. Chinese Journal of Applied Ecology, 2018, 29(12): 3917-3926. (in Chinese)
[7] 魏 凯, 郑建忠, 杨 铭, 何雅琴, 史晓洁, 邓传远. 平潭光幼屿典型植物群落优势种生态位特征[J]. 森林与环境学报, 2020, 40(3): 290-297.
WEI K, ZHENG J Z, YANG M, HE Y Q, SHI X J, DENG C Y. Niche characteristics of dominant species in typical plant communities in Guangyou Island, Pingtan[J]. Journal of Forest and Environment, 2020, 40(3): 290-297. (in Chinese)
[8] LUCIE M, FLORIAN F, CYRILLE V, GRÉGOIRE T F. Multiple facets of diversity effects on plant productivity: species richness, functional diversity, species identity and intraspecific competition[J]. Functional Ecology, 2020, 34(1): 287-298.
[9] ITAI G, JONATHAN B. Niche breadth and species richness: correlation strength, scale and mechanisms[J]. Global Ecology and Biogeography, 2020, 29(1): 159-170.
[10] 徐华林, 袁天天, 王 蕾, 关开朗, 廖文波. 广东内伶仃岛台湾相思群落在15年间的演替研究[J]. 生态科学, 2016, 35(4): 12-22.
XU H L, YUAN T T, WANG L, GUAN K L, LIAO W B. Succession ofcommunities during fifteen years in Neilingding Island of Guangdong Province, China[J]. Ecological Science, 2016, 35(4): 12-22. (in Chinese)
[11] SAUD L A R, GAMEEL S S A N, ABDULAZIZ M AJOSÉ M F, BASHARAT M D, MAGDY I E B, AHMED M A E. Infection by Plicosepalus curviflorus mistletoe affects the nutritional elements of Acacia species and soil nutrient recycling in an arid rangeland[J]. Plant Ecology, 2020, 221(11): 1-12.
[12] 张秀芳, 穆振北, 林美娇, 江淼华, 巩嘉欣, 游巍斌. 琅岐岛4种优势植物叶功能性状及其影响因子[J]. 应用与环境生物学报, 2020, 26(3): 667-673.
ZHANG X F, MU Z B, LIN M J, JIANG M B, GONG J X, YOU W B. Functional traits of leaves of four dominant plants on Langqi Island, Fuzhou, and factors influencing these traits[J]. Chinese Journal of Applied and Environmental Biology, 2020, 26(3): 667-673. (in Chinese)
[13] 蔡晓禾, 廖 廓. 福建平潭大风气候特征分析[J]. 闽江学院学报, 2011, 32(5): 130-133.
CAI X H, LIAO G. Analysis of climatic characteristics for gale in Pingtan of Fujian[J]. Journal of MinJiang University, 2011, 32(5): 130-133. (in Chinese)
[14] 赵林洪. 基于GIS和RS的海岛型城市生态脆弱性评价[D]. 福州: 福州大学, 2018.
ZHAO L H, Evaluation of island city’s ecological vulnerability based on GIS and RS: a case study of Pingtan Island[D]., 2018. (in Chinese)
[15] 福建省科学技术委员会. 福建植物志-第1~6卷[M]. 福州: 福建科学技术出版社, 1982.
Editing Committee of Fujian Flora. Annals of Fujian flora[M]. Fuzhou: Science and Technology Press, 1982. (in Chinese)
[16] 编辑委员会中国科学院中国植物志. 中国植物志[M]. 北京: 科学出版社, 2004.
Editorial Board of Flora of China, Chinese Academy of Sciences. Flora of China[M]. Beijing: Science Press, 2004. (in Chinese)
[17] 张坚强. 海坛岛次生植被物种多样性与群落特征[D]. 福州: 福建农林大学, 2019.
ZHANG J Q. Species diversity and community characteristics of secondary vegetation in Haitan Island[D]. Fuzhou: Fujian Agriculture and Forestry University, 2019. (in Chinese)
[18] LEVINS R. Evolution in changing environments: some theoretical explorations. (MPB-2)[M]. Princeton: Princeton University Press, 1968.
[19] PIANKA E R. The structure of lizard communities[J]. Annual Review of Ecology and Systematics, 1973, 4(1): 53-74.
[20] 马金双. 中国入侵植物名录[M]. 北京: 高等教育出版社, 2013.
MA J S, The checklist of Chinese invasive plants[M].Beijing: Higher Education Press, 2013. (in Chinese)
[21] 闫小玲, 刘全儒, 寿海洋, 曾宪锋, 张 勇, 陈 丽, 刘 演, 马海英, 齐淑艳, 马金双. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性, 2014, 22(5): 667-676.
YAN X L, LIU Q R, SHOU H Y, ZENG X F, ZHANG Y, CHEN L, LIU Y, MA H Y, QI S Y, MA J S. The categorization and analysis on the geographic distribution patterns of Chinese alien invasive plants[J]. Biodiversity Science, 2014, 22(5): 667-676. (in Chinese)
[22] 郑维艳, 曾文豪, 唐一思, 石 慰, 曹坤芳. 中国大陆北热带及亚热带地区樟科、壳斗科物种多样性及其生物地理格局分析[J]. 生态学报, 2018, 38(24): 8676-8687.
ZHENG W Y, ZENG W H, TANG Y S, SHI W, CAO K F. Species diversity and biogeographical patterns of Lauraceae and Fagaceae in northern tropical and subtropical regions of China[J]. Acta Ecologica Sinica, 2018, 38(24): 8676-8687. (in Chinese)
[23] IRL S D H, SCHWEIGER A H, STEINBAUER M J, AHPENG C, ARÉVALO J R, BEIERKUHNLEIN C, CHIARUCCI ALESSANDRO, DAEHLER C C, FERNÁNDEZPALACIOS J M, FLORES O, KUEFFER C, MADĚRA P, OTTO R, SCHWEIGER J M I, STRASBERG D, JENTSCH A. Human impact, climate and dispersal strategies determine plant invasion on islands[J]. Journal of Biogeography, 2021, 48(8): 1889-1903.
[24] 杨 博, 央金卓嘎, 潘晓云, 徐海根, 李 博. 中国外来陆生草本植物: 多样性和生态学特性[J]. 生物多样性, 2010, 18(6): 660-673.
YANG B, YANGJIN Z G, PAN X Y, XU H G, LI B. Alien terrestrial herbs in China: diversity and ecological insights[J]. Biodiversity Science, 2010, 18(6): 660-673. (in Chinese)
[25] ANTUNES C, PEREIRA A J, FERNANDES P, RAMOS M, ASCENSÃO L, CORREIA O, MÁGUAS C. Understanding plant drought resistance in a Mediterranean coastal sand dune ecosystem: differences between native and exotic invasive species[J]. Journal of Plant Ecology, 2018, 11(1): 26-38.
[26] INTANON S, WIENGMOON B, MALLORY S C A. Seed morphology and allelopathy of invasiveea[J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2020, 48(1): 261-272.
[27] 郑俊鸣, 李 敏, 张 盟, MUHAMMAD WAQQAS KHAN TARIN, 何天友, 陈凌艳, 陈礼光, 洪小龙, 郑郁善. 滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析[J]. 热带亚热带植物学报, 2021, 29(5): 465-473.
ZHENG J M, LI M, ZHANG M, MUHAMMAD W K T, HE T Y, CHEN L Y, CHEN L G, HONG X L, ZHENG Y S. In-terspecific association and niche of mixed forest commu-nities ofandvar.in the windbreak of sandy coast[J]. Journal of Tropical and Subtropical Botany, 2021, 29(5): 465-473. (in Chinese)
[28] 尼玛曲珍, 张 玺, 谭 坤, 鲁 闯, 王国严, 徐阿生, 罗 建. 西藏色季拉山川滇高山栎群落生态位特征[J]. 生态科学, 2018, 37(3): 51-58.
NIMA Q Z, ZHANG X, TAN K, LU C, WANG G Y, XU A S, LUO J. Niche characteristics ofcommunity on the Sejila mountains, Tibet, China[J]. Ecological Science, 2018, 37(3): 51-58. (in Chinese)
[29] GAUSE G F. Experimental analysis of Vito Volterra’s mathematical theory of the struggle for existence[J]. Science, 1934, 79(2036): 16-17.
[30] 张增可, 吴雅华, 黄柳菁, 刘兴诏. 海岛森林不同演替阶段土壤和植物的碳、氮、磷化学计量特征[J]. 西北植物学报, 2019, 39(5): 925-934.
ZHANG Z K, WU Y H, HUANG L J, LIU X Z. C, N and P stoichiometry of soil and plant in different forest successional stages in Island[J]. Acta Botanica Boreali-Occidentalia Sinica, 2019, 39(5): 925-934. (in Chinese)
[31] 孙 成, 秦富仓, 包雪源, 李 龙, 杨振奇, 董晓宇, 李 艳. 砒砂岩区人工油松林下草本植物的生态位特征[J]. 水土保持通报, 2021, 41(3): 81-86.
SUN C, QIN F C, BAO X Y, LI L, YANG Z Q, DONG X Y, LI Y. Niche characteristics of herbaceous plants under artificialforest in feldspathic sandstone region[J]. Bulletin of Soil and Water Conservation, 2021, 41(3): 81-86. (in Chinese)
[32] 刘润红, 姜 勇, 常 斌, 李娇凤, 荣春艳, 梁士楚, 杨瑞岸, 刘星童, 曾惠帆, 苏秀丽, 袁海莹, 傅桂焕, 吴燕慧. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析[J]. 生态学报, 2018, 38(19): 6881-6893.
LIU R H, JIANG Y, CHANG B, LI J F, RONG C Y, LIANG S C, YANG R A, LIU X T, ZENG H F, SU X L, YUAN H Y, FU G H, WU Y H. Interspecific associations and correlations among the main woody plants in acommunity in a riparian zone of Lijiang River, Guilin, Southwest China[J]. Acta Ecologica Sinica, 2018, 38(19): 6881-6893. (in Chinese)
[33] 陈 林, 辛佳宁, 苏 莹, 李月飞, 宋乃平, 王 磊, 杨新国, 卞莹莹, 田 娜. 异质生境对荒漠草原植物群落组成和种群生态位的影响[J]. 生态学报, 2019, 39(17): 6187-6205.
CHEN L, XIN J N, SU Y, LI Y F, SONG N P, WANG L, YANG X G, BIAN Y Y, TIAN N. Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China[J]. Acta Ecologica Sinica, 2019, 39(17): 6187-6205. (in Chinese)
[34] POLECHOVÁ J, STORCH D. Ecological Niche☆[M]// Fath B. Encyclopedia of Ecology (Second Edition). Oxford: Elsevier, 2019: 72-80.
[35] KARDOL P, FANIN N, WARDLE D A. Long-term effects of species loss on community properties across contrasting ecosystems[J]. Nature, 2018, 557(7707): 710-713.
Niche of Dominant Plant Populations ofCommunity in Pingtan Island
MA Xiaodi1, JIANG Degang2, LIU Zilin1, WANG Fang1, WEN Ruilong1, DENG Chuanyuan1*
1. College of Landscape Architecture, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China; 2. Island Research Center, Ministry of Natural Resources, Fuzhou, Fujian 350400, China
is the main construction species of Acacia forest, it’s an excellent afforestation broad-leaved tree species in Pingtan Island. Understanding the niche characteristics of the main woody and herbaceous plants population is helpful to understand the coexistence and competition mechanism of species in island community. We investigatedcommunities in Pingtan of its inner and subsidiary islands, Levins and Shannon niche breadth and Pianka niche overlap indexes were used to analyze the ecological niche of the main species in the woody layers and herb layers ofcommunities in Pingtan Island. A total of 144 species of vascular plants belonging to 123 genera and 63 families were found, mainly Gramineae and Compositae, significantly different from continental vegetation types. 14 species of invasive plants include Pra,andwere found in the community, which occupied a large niche in the community and may cause some ecological disturbance to the symbiotic population.andin the tree layer,andin the shrub layer, andin the herb layer had higher niche breadth than other species. Species with niche breadth less than 0.5 accounted for 96.67% of the total. The niche breadth of species with higher importance values were generally larger, but not entirely consistent. The niche overlap of dominant population was herb layer (0.253) > shrub layer (0.231) > tree layer (0.149), all smaller than 0.3. The niche overlap between the main species in the community was generally lower, niche differentiation weakened interspecific competition. The strong environmental filterability in the Island led to different degrees of habitat differentiation of the population. The population concentrated in local habitats by narrowing niche breadth, thus promoting the coexistence of species in the island community. Thecommunity on Pingtan Island was the top zonal community with stable interspecific relationship and slow population regeneration.
Pingtan Island;; niche breadth; niche overlap
S794.9
A
10.3969/j.issn.1000-2561.2022.12.024
2022-03-09;
2022-04-09
福建省区域发展科技项目计划(No. 2018Y3006);福建农林大学科技创新专项(No. CXZX2019086);2019年度福建省科技特派员工作经费(No. KTP19186A)。
马晓迪(1998—),女,硕士研究生,研究方向:海岛植物与应用。*通信作者(Corresponding author):邓传远(DENG Chuanyuan),E-mail:dengchuanyuan@163.com。
