气候变化对青藏高原植被生态系统的影响研究进展
2023-01-14曹晓云周秉荣周华坤颜玉倩赵慧芳于红妍
曹晓云,周秉荣,周华坤,乔 斌,颜玉倩,赵 彤,陈 奇,赵慧芳,于红妍
(1.青海省气象科学研究所,青海 西宁 810001;2.青海省防灾减灾重点实验室,青海 西宁 810001;3.青海省气象局科技与预报处,青海 西宁 810001;4.中国科学院西北高原生物研究所,青海省高寒区恢复生态学重点实验室,青海 西宁 810001;5.祁连山国家公园青海服务保障中心,青海 西宁 810001)
引 言
青藏高原平均海拔约4500 m,是世界上最高的独立地貌单元,有着“世界屋脊”、“第三极”和“亚洲水塔”之称,其热力和动力作用对区域与全球气候有重要调节[1],对气候系统稳定、水资源供应、生物多样性保护等具有重要的生态安全屏障作用[2-3]。高原植被复杂多样,水平自然带自东南向西北依次为森林、草甸、草原和荒漠,垂直自然带由海洋性湿润型递变为大陆性干旱型,从喜马拉雅山南侧的热带雨林和常绿阔叶林向北相继更替为针叶混交林、暗针叶林、灌丛、草甸以及高山永久冰雪带,是中国最完整的山地自然景观垂直带谱[4-5],也是生物物种基因库和生物多样性保护地[6]。
作为气候变化的“敏感区”和“启动区”、生态环境变化的“指示器”和“调节器”、全球变化的“驱动机”和“放大器”[3,7],气候的微小波动就会强烈影响高原生态系统,从而改变高原生态系统的格局、过程与功能[6]。近60 a 来,全球变暖问题在青藏高原尤为突出[8-11]。《中国气候变化蓝皮书(2020)》[12]指出:1961—2019年,高原气温平均每10 a 升高约0.37 ℃,升温速率为全球的近2倍,而年降水量平均每10 a 增加约10.40 mm,这势必影响高原生态系统。作为陆地生态系统最活跃的组分,植被能够很好地响应气候变化。因此,气候变化对植被生态系统的影响引起了广泛关注,目前已通过野外调查、台站观测、遥感监测、控制试验与模型模拟等方式获取了大量有价值的研究成果[13-16]。但是,由于各研究采用的数据源不同、选取的时空范围不一,加之对植被生态系统与气候变化的相互作用研究不够深入,仍然存在一些需亟待解决的问题。
本文梳理总结了高原气候变化状况,归纳了气候变化对高原植被生长状况、物候、净初级生产力、生物量碳库、多样性等植被生态系统参量影响的现有研究成果,并进行了讨论与展望,以期进一步厘清青藏高原植被生态系统对气候变化的响应,为开展青藏高原生态安全屏障建设、保证青藏高原生态安全提供科学依据。
1 高原气候变化特征
1.1 气 温
青藏高原多年平均气温在-6.50~12.90 ℃之间,且自东南向西北逐渐降低,东南部气温较高,西北部和中部气温较低。在气候变暖背景下,1961—2019年青藏高原地表气温平均每10 a 升高0.37 ℃[12],其中1979—2018年高原整体升温率为0.46 ℃·(10 a)-1,高于中国、同纬度及全球平均水平,存在“高原变暖放大效应”(图1)[17-19]。高原北部变暖最为显著,尤其柴达木盆地增温率最快[13,20]。此外,高原增温率存在明显的季节差异,冬季增温最显著,尤其是西南部和中部江河源区(念青唐古拉山—唐古拉山—巴颜喀拉山—阿尼玛卿山)[21];高原增温存在显著的年代际变化特征,1982—2000年为缓慢增温时段,2001—2010年为显著增温时段,而2010年之后增温有所减缓,增温率低于北半球平均[22]。
海拔是影响气温变化的重要地形因子,高原变暖存在“海拔依赖现象”(图1)[23-25],高海拔比低海拔地区增温更剧烈[25-26],高原海拔依赖型变暖可能在未来气候变化情景下将持续增强[27],但高原海拔依赖型变暖的主要物理机制还未达成共识[24]。
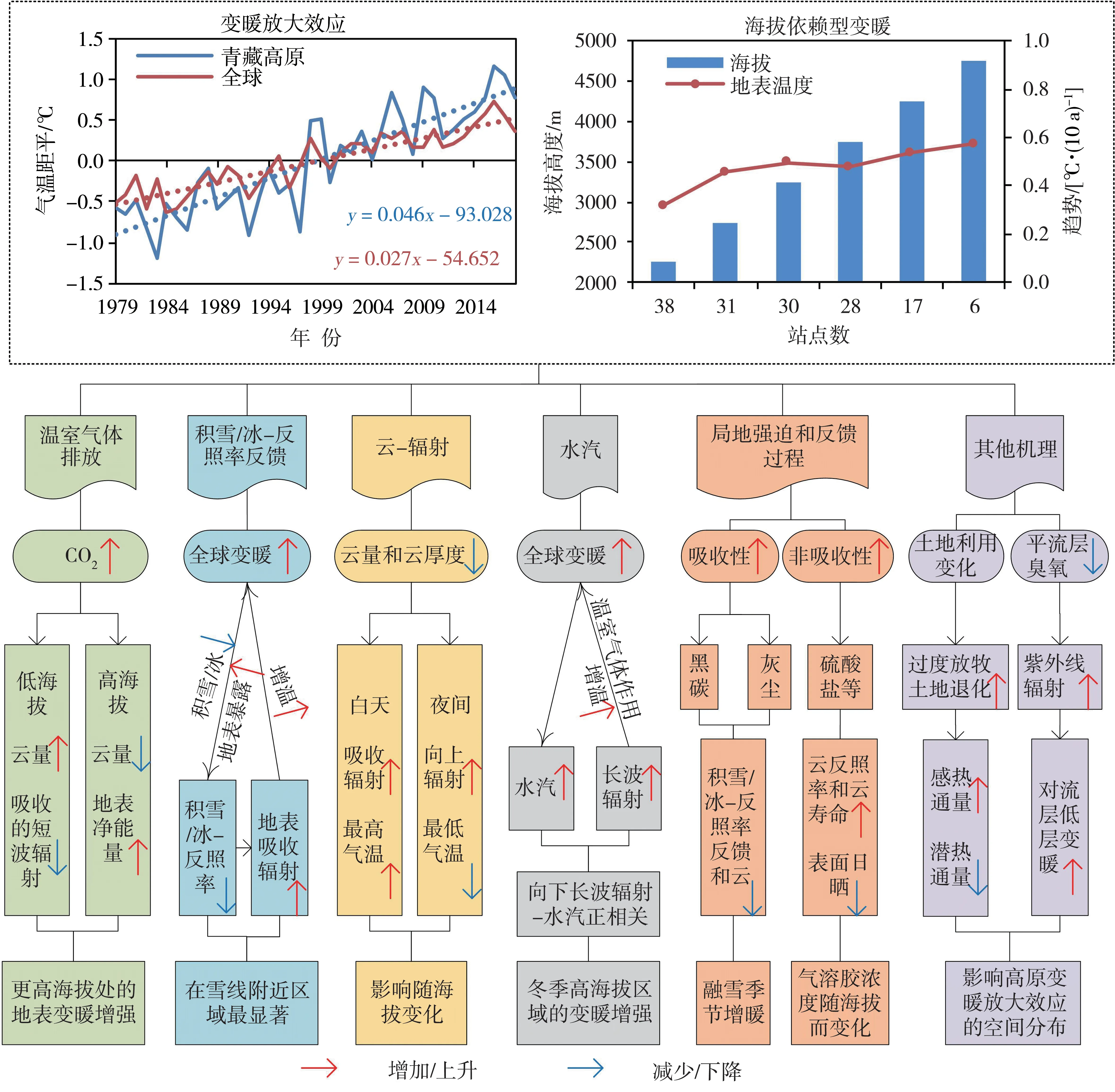
图1 青藏高原变暖放大效应和海拔依赖型变暖的物理机制示意图(引自文献[17])Fig.1 Schematic diagram of the physical mechanisms controlling the profile of warming amplification and elevation dependent warming in the Tibetan Plateau (cited from literature [17])
1.2 降 水
受季风和地形影响,青藏高原降水空间差异显著,多年平均降水量为50.0~2000.0 mm(整体平均约472.4 mm),且自东南向西北递减[28-29]。近几十年来,高原降水量总体呈增多趋势,气候倾向率为10.40 mm·(10 a)-1(1961—2019年),高原整体趋于湿润[12,30],但20世纪80年代后降水量总体增加不显著[31-32]。降水量变化的空间格局差异较大,高原中部和北部干旱半干旱地区降水量增加显著,而东南部和南部湿润区则呈减少趋势(图2)[31]。高原四季降水量均表现出增多趋势,但春季降水量增幅最大[33]。另外,从雨季起讫期来看,1961—2017年高原雨季开始期平均每 10 a 提前1.4 d,结束期推迟0.8 d,雨季延长2.2 d,降水量增加6.5 mm,且雨季降水极端性增强,2007年以后表现尤为明显,2007年前后中雨日数、1日最大降水量、强降水量和降水强度分别增加1.0 d、9.3%、16.9%和0.4 mm·d-1[34]。预测结果表明,未来高原东部的祁连山区、柴达木盆地东部、青海湖流域与长江源区极端降水事件将明显增加,高原中西部地区发生强降水的可能性亦增大,而高原东南边缘的干旱事件将增多[31,35]。

图2 1961—2015年青藏高原年降水量变化空间分区(引自文献[31])(a)变化趋势;(b)波动特征Fig.2 The spatial division of annual precipitation change in the Tibetan Plateau from 1961 to 2015 (cited from literature [31])(a) variation trend,(b) fluctuation characteristics
总体而言,高原东南部呈暖干化、西北部呈暖湿化的态势。然而,由于高原西部尤其是藏北高原气象台站稀少,长时间序列的5000 m 以上台站观测资料十分有限,该地区的气候变化特征还需进一步论证。
2 高原气候变化对植被的影响
2.1 植被覆盖度与NDVI
归一化植被指数(normalized difference vegetation index,NDVI)是有效反映绿叶密度、光合有效辐射及累积生物量等参数的重要指标,广泛应用于大尺度地表植被覆盖和生长状况监测研究[36-38]。研究表明,1982年以来青藏高原植被在整体改善背景下存在局部退化态势,改善面积占比67.7%~75.0%,显著改善面积占比30.0%左右,主要分布在高原中东部,而高原西部、南部和东北部的部分地区呈退化趋势[39-41]。
高原气温、降水及高寒植被生长限制因子的空间差异导致植被变化趋势存在显著的时空异质性,且不同植被类型在不同阶段的表现不同[41-42]。其中,1982—2006年,低覆盖度的植被改善,高覆盖度的植被退化;针叶林、阔叶林的覆盖度下降,其他类型植被的覆盖度上升[43];湿润半湿润及部分半干旱地区植被改善,半干旱和干旱地区植被退化[44]。基于变化点检测分析发现,1982—2012年高原58.9%的区域植被状态趋于稳定,32.3%的区域植被变化显著,植被改善面积大于退化面积,中东部植被呈改善趋势,西部植被略有退化趋势,而南部和东北部植被退化趋势明显[45-46]。1982—2020年,高原植被NDVI 整体呈显著波动增加趋势,每10 a 增加0.006[41](图3);在季节尺度上,四季NDVI均呈增加趋势,春、秋季增加尤为显著;荒漠、草原、草甸和森林植被NDVI 也表现出波动增加趋势,但不同类型植被的阶段性变化规律有所不同[47]。2000年以来,高原植被NDVI 变化存在明显的空间异质性,大部分地区植被生长良好、NDVI 增加,局部地区有所退化、NDVI 减小,NDVI 整体呈微弱增加趋势[48]。其中,高寒草原、高寒荒漠NDVI 呈增加趋势,而高寒草甸和针叶林则呈减小趋势[49];干旱和半干旱区植被NDVI 增加面积占比明显大于湿润和半湿润区[50]。在植被改善大环境下,原有退化植被的退化程度存在进一步加重的现象[51]。
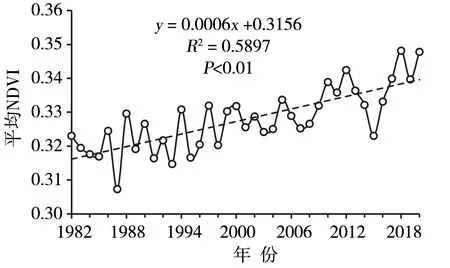
图3 1982—2020年青藏高原植被生长季 NDVI 变化(引自文献[41])Fig.3 The change of vegetation NDVI in growth season in the Tibetan Plateau from 1982 to 2020(cited from literature [41])
气候变化是高原生态系统变化的主控因子,青藏高原植被生长受水热因子驱动的区域占总面积的56%[41],气候暖湿化对高原植被覆盖度、NDVI 为正影响,且存在时空不均衡性。其中,湿润区生长季植被NDVI变化主要受气温影响,气温升高促进植被生长发育;干旱和半干旱区降水对植被NDVI的影响明显高于湿润和半湿润区,降水减少抑制植被生长发育[50,52],且在干旱年份过度放牧、垦殖、樵采等人类活动会促使西部干旱和半干旱区植被严重退化、NDVI 显著降低。然而,气候因子对植被NDVI 的影响存在一定的时滞性,且响应时间存在空间差异,从当月到滞后6 个旬不等[53]。据统计,高原有超过43%的区域植被受气候时间滞后影响[54],联合考虑气候滞后与累加效应十分重要。此外,极端气候事件也会给植被乃至整个地表环境造成严重后果[55]。
2.2 植被物候
物候指植物受环境影响出现以年为周期的自然生长发育节律,是气候变化最敏感的指标之一[53]。1982年以来,高原植被物候整体呈返青期提前、枯黄期推迟、生育期延长的趋势[29,53,56-60],植被返青期提前和生长期延长的区域主要集中在高原东部[61],但2000年后高原植被返青期是否有提前趋势存在较大争议。
1982年到1990年代中期高原植被返青期显著提前,平均每年提前0.31~0.88 d[62-65]。考虑非生长季对物候提取的影响,高原植被返青期在2006年前呈提前趋势,2006年后返青期开始推迟,而1982—2012年植被枯黄期无明显变化[62]。关于2000年后高原植被返青期是否提前?常清等[66]认为,2003—2012年高原高寒草地植被返青期呈提前趋势。宋春桥等[67]发现,2001—2010年藏北高原约60%的草地植被返青期呈提前趋势,枯黄期变化不明显,生育期主要受返青期影响呈延长趋势。SHEN 等[15,68]认为,2000—2011年高原东北部气温上升、降水增加引起返青期提前,而西南部降水减少引起返青期推迟,返青期整体无明显变化趋势。孔东东等[69]通过偏最小二乘法回归分析发现,高原植被返青期转折发生在1997—2000年,枯黄期转折发生在2004—2007年,生育期转折发生在2005年左右,转折前每10 a 返青期平均提前2~3 d、枯黄期平均延迟1~2 d、生育期平均延长1~2 d;转折之后返青期推迟趋势的显著性水平仅为0.1,而枯黄期、生育期的变化趋势不显著(图4)。整合NDVI 的3 种长序列数据集(GIMMS、SPOT-VGT 和MODIS)发现,结论争议较大的原因是2000年后的GIMMS NDVI 数据存在严重的质量问题[70]。可见,遥感数据的时空尺度差异导致高原植被物候分析结果产生偏差,2000—2006年GIMMS NDVI 数据质量问题影响了高原长序列物候期监测结果的准确性[61]。近年来,高原植被返青期提前及枯黄期推迟现象有所减缓甚至出现逆转[71-73],2001—2017年植被返青期、枯黄期整体分别以3、1 d·(10 a)-1的速率呈推迟趋势[73]。树木年轮数据结果显示,1960—2014年高原植被返青期和枯黄期每10 a 分别提前2.8 d 和推迟3.3 d[74],证实了气候变化影响下高原植被返青期提前、枯黄期推迟、生育期延长的事实,但高原植被物候分布和变化趋势在不同海拔和坡向上的差异十分显著,高海拔植被的变化趋势比低海拔更为复杂[75-77]。

图4 青藏高原物候指标突变前后趋势及全部年份整体趋势(引自文献[69])Fig.4 The phenological metrics’ trend before and after turning points and trends of the whole research period in the Tibetan Plateau (cited from literature [69])
由于植被物候对气候变化的响应并非线性相关,加之地形复杂的高原各地区气候条件与变化程度不同[75-77],气候变化对植被物候的影响机制众说纷纭。SHEN 等[68]认为,21世纪初高原西南部植被返青期推迟与印度季风减弱导致的该地区降水量减少密切相关。而YU 等[62]则认为,返青期推迟可能是冬季增温减弱了春化作用,进而导致2000年后春季返青期推迟。此外,不同于北半球高纬度大部分地区植被返青期主要受白天温度变化的影响,高原植被返青期与夜间温度的关系更为显著[15];极端天气也会影响植被物候期,霜冻对高原植被返青期和枯黄期的影响程度最大、范围最广,霜冻日数增加有利于高原植被提前结束生长以躲避不利环境的损害[47,78];冬季积雪增多会导致次年春季土壤水分增加,有利于高原植被返青期提前[79-80]。
2.3 植被净初级生产力
植被净初级生产力(net primary productivity,NPP)定义为单位面积及时间内绿色植物通过光合作用积累的有机物数量,是衡量植被群落在自然条件下生产力、陆表生态系统健康状况以及判定碳汇和碳源的核心指标[81-82]。青藏高原植被NPP 多年平均约0.21 PgC,占中国总量的10%左右[81]。1982年以来,高原植被NPP 变化存在较大的空间差异,显著增加区主要分布在祁连山南部和念青唐古拉山北部高寒草甸区[14,82],而藏北高原、西藏“一江两河”和三江源中西部地区则呈下降趋势[14],但整体呈显著增加趋势[14,83-85],且2000年后增速减缓[13]。研究发现,1990—2013年潜在的NPP 整体呈平稳上升趋势,人类活动影响下1993年以前NPP 为负值,之后均为正值,即1994年后实际的NPP 持续大于潜在的NPP(图5),尤其在2000—2002年[13],表明恢复措施对高原植被的改善已初见成效。
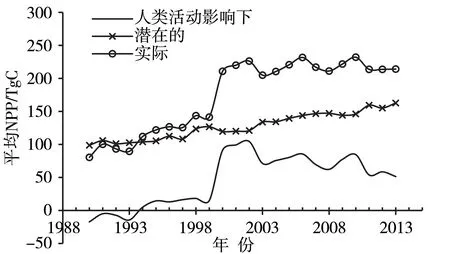
图5 1990—2013年青藏高原植被净初级生产力年际变化(引自文献[13])Fig.5 Inter-annual variation of vegetation net primary productivity in the Tibetan Plateau from 1990 to 2013 (cited from literature [13])
此外,1980年以来高原不同植被类型NPP 增加幅度不同,荒漠增幅最大,其次是草甸、草原、森林和灌丛[86]。2000年以来高原植被NPP 整体表现高位波动增加态势,植被NPP 重心总体向西南方向移动,表明西南部植被NPP 在增量和增速上大于东北部[13,87-89]。单点和典型区域的植被NPP 表现出一致态势。1961年以来,三江源区植被NPP 整体呈显著上升趋势,气温升高导致该区域植被NPP 增加[90],其中五道梁地区植被NPP 整体也呈显著上升趋势,这主要是气温升高引起的生育期延长及生物量快速增长所致[91-92]。
总体上,气候变化主导着高原大部地区植被NPP 的变化[79]。然而,植被生长变化是多因子共同作用的结果,其定量化归因研究仍是难点,关键影响因素一直存在争议,但大部分研究认为气温、降水和干旱的影响较深[93-94]。积温是影响植被生长发育的主要气象因子,是决定高原植被NPP 的基础[95]。相对于夏季,春、秋季增温对高原植被生产力的促进作用更强[63]。在降水匮乏的高原干旱半干旱区,植被生产力变化的主导因子是降水,而在降水充沛的高原湿润和半湿润区,主导因子则为气温[96]。极端干旱对植被生长有明显抑制作用,严重影响了植被生理和土壤微生物活性,进而降低植被生产力[97],甚至导致植被死亡[47]。
2.4 植被生物量碳库
植被生物量是地表碳循环的重要组成部分和土壤碳库的主要输入源[98-99]。净生态系统生产力(net ecosystem productivity,NEP)表征了陆地生态系统与大气之间的净碳交换,是定量评价陆地生态系统碳源/汇的重要指标[100],广泛应用于碳循环研究中。高原植被生物量的平均碳密度约为223 gC·m-2,碳库约为277 TgC[101]。1982年以来,高原植被生物量碳库呈波动增加趋势,总体表现为碳汇,但存在明显的区域异质性,其中高寒草甸草原增加显著,其他草地类型增幅较小,部分地区有所下降[101-103]。
由于高原土壤极大的空间异质性,加之不同估算模型的差异及模型对生态系统呼吸反演参数和土壤碳循环理解不足,目前高原植被生物量碳库的研究仍存在较大不确定性[104-105],且长时间序列变化及响应研究也相对薄弱,多集中于草地[106-107]。研究表明,高原植被碳储量因土壤和植被类型而存在较大的空间异质性[108],碳密度整体呈东南高、西北低的空间格局[108-109];2000年以来,高原植被碳汇功能逐渐增强,平均每年固碳量约-3.20~64.42 TgC,整体呈平稳上升趋势[103]。其中,2001—2015年高原草地碳汇区面积约86.38万km2,约占高原总面积的57.3%,草地NEP 整体呈波动增加趋势,增速为0.53 gC·m-2·a-1[图6(a)],但高原东部增加趋势明显,而中部和西南部减少趋势明显[110],草地退化[111]和土地利用格局变化[112]引起了高原植被有机碳释放[图6(b)]。由通量观测站数据构建的草地NEP估算模型也表明,2001—2010年三江源区草地NEP具有较大的空间异质性,大部分地区表现为碳汇,NEP 平均为41.8 gC·m-2,整体呈波动增加趋势,增速为5.4 gC·m-2·a-1[110]。优化后的CLM4.5 模型模拟结果显示,1970—2010年高原植被碳储量整体呈下降趋势,2006年出现转折后开始轻微上升,且自西北向东南呈先降后升的分布趋势;预计2020—2050年,除南部阔叶林外,高原植被碳储量整体呈持续上升趋势,增速为4.2 gC·m-2·a-1[105]。

图6 青藏高原2001—2015年草地生态系统NEP的变化趋势(a)(引自文献[110])和不同类型植被碳储量(b)(改自文献[112])Fig.6 The change trend of NEP in grassland ecosystem from 2001 to 2015 (a) (cited from literature [110]) and carbon storage of vegetation with different types (b) (modified from literature [112]) in the Tibetan Plateau
气候变化主要通过温度、降水的变化影响植被生产速率、凋零速率及微生物活性,改变地表凋零物和土壤有机碳的分解速率、土壤呼吸速率,进而影响土壤碳的蓄积过程[113]。然而,不同类型草地生态系统的碳循环过程对温度和降水变化的响应有所差异,高寒草甸的碳循环过程对温度变化的响应更敏感[114],而高寒草原的碳循环过程对降水变化的响应更敏感[115]。相比高寒草原,高寒草甸受干旱胁迫影响较小,升温引起的高寒草甸植被生产力增量更大,进而导致更多植被固定的有机碳输入到土壤中[116-117]。植被生产力的增加是高原主要的碳汇机制[79,118],水热匹配是气候变化过程中植被碳积累过程的关键驱动因素[119]。随着气候变化的影响,高原碳汇总体表现为增加[119],但升温引起土壤有机碳分解速率加快[120],致使高原草地碳储量增加不显著。此外,植物物候对碳积累也有重要影响[121],长期围封不利于高原高寒草甸植被碳储量的积累,尤其不利于地下根系碳储量的固存[122]。尽管目前已有不少有关植被生物量碳库的研究,但气候变化对高原不同植被类型生态系统碳源/汇动态的影响仍不明确[123-124]。
2.5 植物多样性
生物多样性给人类提供基本的生活环境和丰富的资源[125-126],而气温和水分的动态平衡则决定着植物多样性分布格局[127-128]。气候变化对生态系统水热条件影响显著,是导致生态系统物种组成和群落结构发生变化的主要原因之一[129-130]。近年来,高原植物多样性发生急剧变化[131]。尽管研究结果不尽相同,但气候变化显著影响了高寒草地植物群落、物种组成和植物多样性的事实毋庸置疑。
受气候变化影响,青藏公路124 道班的华扁穗草群落被适应干旱环境的矮嵩草群落替代,而原先的矮嵩草群落则慢慢演变成高山嵩草群落,且在进一步干旱化后形成沙生薹草群落[132]。野外增温模拟试验表明,增温会使高原植物群落物种丰富度快速降低26%~36%[127],尤其是高寒草原,但增温对高寒草甸群落多样性无显著影响[133],如海北高寒草甸植物物种多样性随增温呈先增后减的变化趋势,草甸、灌丛群落的物种数量分别减少7%、30%[134-135];在高原腹地典型的高寒草甸和沼泽草甸区,适度增温导致3个物种的重要值上升、5个物种的重要值下降,而过度增温则使物种趋于单一化[135-136]。然而,长期(10 a)的海拔梯度移栽试验分析发现[137],由于物种滋生的数量大于消失的数量,增温和降温均有利于高寒草甸植物物种丰富度的提高,这种效应取决于伴随气候变化的物种特性和数量。研究认为,气候变化可能导致21世纪全球植物多样性迅速降低[138],但长期来看物种丰富度会通过定居(散播和种子库)和竞争性释放(优势物种转移)的结合而恢复[139-140]。由此可见,气候变化显著改变了高原植被物种组成和群落结构,进一步影响植被群落的重组和更替[140],但未必会导致区域植物多样性的下降[141]。由于增温方法不一且研究时间普遍较短,研究结论还需进一步完善。
气候变化影响着高原植物群落的物种组成和多样性,从而改变了植被群落的结构和功能[4]。然而,气候变化对植物多样性的影响,目前仍是一个不断研究和认识的过程。一方面,气候变化引起一些敏感的生态系统发生变化,导致一些物种在局部地区消失;另一方面,气候变化影响了高纬度和高海拔地区适应寒冷气候的植被,升温使得这些物种“往北走、往上走”。此外,气候变化会引起物种间的关系发生变化,甚至会造成植被遗传多样性的消失[126,142-144]。
3 结论与展望
(1)近60 a来,青藏高原气温整体呈显著升高趋势,平均每10 a 升高约0.37 ℃,降水量整体呈增加趋势,平均每10 a 增加约10.40 mm,东南部表现暖干化趋势,西北部表现暖湿化趋势。
(2)高原植被在整体改善背景下存在局部退化态势,且年际波动明显,改善面积占总面积的67.7%~75.0%,其中显著改善面积约为30.0%,主要分布在中东部,而西部、南部和东北部的部分地区植被呈退化趋势。高原干旱和半干旱区植被NDVI增加面积占比明显大于湿润和半湿润区,但在植被改善大环境下原有退化植被的退化程度进一步加重,气候暖湿化对高原植被覆盖度、NDVI 为正面影响,但存在时空差异,湿润区植被NDVI 变化主要受气温影响,而干旱半干旱区受降水影响更明显。另外,气候因子对植被NDVI 的影响存在一定的时滞性,响应时间从当月到滞后6个旬不等。
(3)高原植被物候总体呈返青期提前、枯黄期推后、生长期延长的趋势,且存在明显的空间异质性,返青期提前的区域主要集中在高原东部。气候变化对植被物候的影响机制较为复杂,除气温和降水外,极端天气和积雪对物候期的影响也不容忽视。
(4)高原植被NPP 整体呈显著增加趋势,但也存在较大的空间差异,祁连山南部及念青唐古拉山北部高寒草甸为显著增加区域,而藏北高原、西藏“一江两河”和三江源中西部地区则呈下降趋势,且2000年后高原植被NPP 增加速率变缓。不同类型植被NPP 的增加幅度不同,荒漠增量最大,其次是草甸、草原、森林和灌丛。气候变化主导了高原大部分地区植被NPP 变化,但定量化的关键影响因素一直存在争议。
(5)高原植被生物量碳库呈波动增加趋势,总体表现为碳汇,但也存在明显的空间异质性,高寒草甸草原增加显著,其他草地类型增幅较小,部分地区有所下降。植被生产力增加是高原主要的碳汇机制,但不同类型草地生态系统的碳循环过程对温度和降水的响应有所差异。由于土壤的空间异质性、不同估算模型间的差异及其对生态系统呼吸反演参数和土壤碳循环理解不足,目前高原草地生物量碳库的研究仍存在较大的不确定性,气候变化对高原不同植被类型生态系统碳源/汇动态影响仍不明确。
(6)高原植物多样性随气候变化发生了显著变化,尽管研究结果不尽相同,但气候变化显著影响了高寒草地植物群落物种组成和多样性这一事实毋庸置疑。由于模拟增温方法不同且研究时间普遍较短,研究结论仍需进一步完善。
气候变化是影响高原植被生态系统变化的主控因子,且这种影响存在时空不均衡性,而人类活动增强导致的高原植被退化也不容忽视。针对高原植被退化现象,国家实施了退耕还林、退牧还草、天然林防护等一系列重大工程与生态环境保护措施,已取得了成效。然而,面对持续的气候变化和更加脆弱的生态系统,现有研究对高原植被及其与气候变化的相互作用仍存在一些认知上的不足,亟待去解决。
(1)现有高原植被对气候变化的响应研究依靠的卫星遥感影像尺度单一,无法准确反映地表实际情况,加之高原西部特殊地形、气候和环境条件使得气象站点分布稀少,造成西部藏北高原等地区观测存在许多盲点。此外,遥感反演及模式模拟缺乏观测基础,致使遥感监测和模式模拟结果难以比较。因此,未来应加强高原西部地区数据的组网观测对比及精度验证的遥感理论与技术研究,并强化多时空尺度的研究。
(2)当前气候变化对高原植被的影响研究多集中于气温、降水与植被的相互作用分析,而极端天气、蒸散发、土壤水分等也是影响植被分布及变化的重要因子,但多因子对植被生态系统影响的综合量化分析仍较为欠缺,且对植被关键耦合机制的影响仍然不确定。因此,未来应进一步加强多种气候因子与植被的内在机理研究,并综合量化分析其对高原植被生态系统的影响。
(3)受高原高海拔、复杂地形以及相对落后的基础设施影响,开展野外观测、控制试验与采样调查十分艰难。因此,未来应加强团队合作,逐步建立全域覆盖的长期观测网,共享数据,节约科研资源,减少不必要的重复采样。
(4)高原植被生态系统极为敏感、脆弱,局部退化现象仍未得到有效遏制。因此,高原草地适应性管理刻不容缓,如何平衡牧民生活水平和载畜量、保障技术和资金投入是一个关键问题,未来需要强化部门联动、提升生态保护修复技术、完善管理体系,加大生态保护工程建设力度和补偿政策,整体提升高原应对全球气候变化的能力。
