蓝藻膦酸酯代谢及其生态意义研究进展*
2023-01-13彭思慧黄淑敏林立洲郑凌凌宋立荣束文圣李春连
彭思慧,黄淑敏**,曾 莹,林立洲,郑凌凌,章 颖,宋立荣,束文圣,李春连,赵 亮,
(1:华南师范大学生命科学学院,广东省植物发育生物工程重点实验室,广州市亚热带生物多样性与环境生物检测重点实验室,广州 510631) (2:中国科学院水生生物研究所,淡水生态与生物技术国家重点实验室,中国科学院藻类生物学重点实验室,武汉 430072) (3:广东省科学院微生物研究所,华南应用微生物国家重点实验室,广东省菌种保藏与应用重点实验室,广州 510070) (4:浙江省越崎中学,绍兴 312050)
对蓝藻水华形成与优势维持机理的解析,是高效水华防控与减灾的必要前提[1-2]。在水生生态系统中,磷的循环是典型的沉积型循环,使磷元素成为水体最常见的生长限制因子[3]。因此,磷的高效获取与利用,是维持水华和水体蓝藻巨大生物量的重要过程,也是学界关注的热点问题。+5价的正磷酸盐是最易被浮游植物吸收利用的无机磷,故其亦成为水环境中最稀缺的磷形态[4]。当无机磷耗尽时,蓝藻转而利用溶解性有机磷(dissolved organic phosphorus, DOP)[5]。水体中的DOP主要由两类物质组成,即含磷酯键(C-O-P)的磷酸酯类化合物(phosphoester,图1A、B)与含有碳-磷键(C-P)的膦酸酯类化合物(phosphonate,图1C~E);其中磷酸酯类化合物可被生物高效利用供磷,而膦酸酯类化合物因其含化学结构稳定的C-P键,可抵抗各种水解、酶解与光分解等,常被归为生物不可利用磷[5-9]。

图1 几种常见的磷酸酯(A:核酸;B:磷脂)与膦酸酯(C:甲基膦酸;D:2-氨基乙基膦酸;E:草甘膦)(图中红色虚线圈表示C-O-P键;绿色实线圈表示C-P键)Fig.1 Formulas of several common phosphate esters (A: nucleic acid; B: phospholipids) and phosphonates (C: methyl phosphonic acid; D: 2-amino ethylphosphonic acid; E: glyphosate) (The red dotted circle shows the C-O-P bond; the solid green coil shows the C-P bond)
在近些年的研究中,越来越多的微生物(包括数十株蓝藻)被证实可以分解膦酸酯,显示了膦酸酯作为生物替代性磷源的可能性,深化了对水体磷循环的认识[7-15]。同时,膦酸酯在水环境中具有广泛来源,包括人工合成(如草甘膦、敌百虫等)与生物合成(如甲基膦酸)等,使其成为水体有机磷库的重要组成[7-9,15-17]。随着研究的深入,膦酸酯的生态风险日益凸显,其可作为环境污染物与磷源影响水体浮游生物竞争格局[18-24],而生物源的甲基膦酸亦可作为好氧甲烷生物发生的关键底物[14-15,25-26]。此外,经过长期的水质管理,部分富营养化水体已呈现常态化无机磷限制,但仍可见蓝藻水华周期性暴发[27]。作为广泛存在的生物可替代性磷源,膦酸酯在上述过程中发挥的作用值得深入探究。
本文从膦酸酯生物合成与分解机制出发,回顾了蓝藻膦酸酯代谢研究历史,对目前已有的蓝藻膦酸酯利用与合成研究进行梳理,并重点综述了蓝藻膦酸酯代谢过程生态风险与生态意义的研究进展;最后对本领域未来发展趋势进行了评述,以期为蓝藻水华形成与优势维持机理的研究探索新思路,为针对性控磷政策的制定提供理论依据。
1 磷与膦酸酯
有机膦酸酯是一类古老的分子,在原始的缺氧时期,其被认为是地球有机磷的主要形态[28]。膦酸酯中的磷为+3价,因此其亦构成了水体磷氧化还原循环中最主要的成分[6,29]。
1.1 水环境中的膦酸酯
膦酸酯在水环境中广泛存在,在海洋环境中,膦酸酯可占DOP的10%~25%[6,30-31];而在部分淡水水体中,DOP中膦酸酯可达35%[32-33]。水体中的膦酸酯主要由生物合成产生,但也有相当一部分膦酸酯来源于人类活动。
1.1.1 人工合成的膦酸酯 1944年,Pikl合成了氨甲基膦酸(aminomethylphosphonic),这是人类首次成功合成膦酸酯[7]。自此,越来越多人工合成膦酸酯被应用于商业,包括制药、农业、造纸与环境等领域。例如,氨基三亚甲基膦酸是水处理过程中常用的絮凝剂,泰诺福韦(tenofovir)被用于Ⅰ型艾滋病的治疗,三氯松(敌百虫)被用作杀虫剂,乙烯利被用作植物生长调节剂等[7-9,34-35]。此外,人工合成膦酸酯还常作为阻燃剂与增塑剂等被广泛使用[7-9]。
在所有人工合成的膦酸酯中,草甘膦(N-(膦酸甲基)甘氨酸,图1E)是最广泛使用的产品,占全球除草剂使用量的25%[36]。美国每年至少有2万t膦以除草剂的形式被释放到环境中[37],而我国2020年草甘膦产量已逾55万t[38]。尽管草甘膦具有高效、低毒等特点,但其大规模使用会引发水体污染、危害人类健康,已成为《生活饮用水卫生标准》(GB5749-2006)中严格控制的有机污染物,对其生态风险的研究也日趋深入。
1.1.2 生物源的膦酸酯及其生物合成 虽然科学家在1944年就已实现膦酸酯的人工合成,但直至1959年才首次鉴定到生物源的膦酸酯——2-氨基乙基膦酸(2-amino ethylphosphonic acid, 2-AEP),而2-AEP也被认为是野外丰度最高的生物源膦酸酯[7]。目前人们已在不同的原始生物中鉴定出多种生物源膦酸酯,如从弗氏链霉菌(Streptomycesfradiae)中分离的抗生素磷霉素(fosfomycin),从产绿色链霉菌(Streptomycesviridochromogenes)中分离的除草剂草胺膦(phosphinothricin)以及从海洋古菌Nitrosopumilusmaritimus中分离的甲基膦酸等[39-40]。上述膦酸酯常以胞外多糖、糖蛋白及磷脂的修饰基团或以抗生素的形式在生物中存在。膦酸酯基团的修饰,被认为可增加上述大分子的结构强度,以抵御磷酸酶酶解等[7-9]。
虽然生物源膦酸酯结构多样,但是其生物合成途径却极其保守:其共享生物合成前体,即磷酸烯醇式丙酮酸(phosphoenolpyruvic acid, PEP)、膦酸丙酮酸(phosphonopyruvate, PnPy)和膦酸乙醛(phosphonoacetaldehyde,PnAA)[7-9]。在磷酸烯醇式丙酮酸变位酶(PEP mutase,PepM, EC 5.4.2.9)和膦酸丙酮酸脱羧酶(PnPy decarboxylase,Ppd, EC 4.1.1.82)的作用下,PEP被转化为PnAA,并形成C-P键[7-9]。因此,pepM也常作为膦酸酯生物合成能力筛选的标记基因;研究发现,10%的细菌编码该基因[11]。PnAA作为其他膦酸酯的前体,可用于广泛的产物合成。例如,PnAA经过脱氢酶(PnAA dehydrogenase, Pdh)加氢形成羟乙基膦酸(hydroxyethylphosphonate, HEP),HEP再经由MPn合成酶(MPn synthase, MpnS)转化形成甲基膦酸(图2)[16]。作为甲基膦酸合成的关键酶,研究发现0.6%的海洋微生物含编码MpnS的基因,而这一丰度被认为可维持水体表层的高甲基膦酸浓度,以解释水体表面甲烷的超饱和现象[16]。
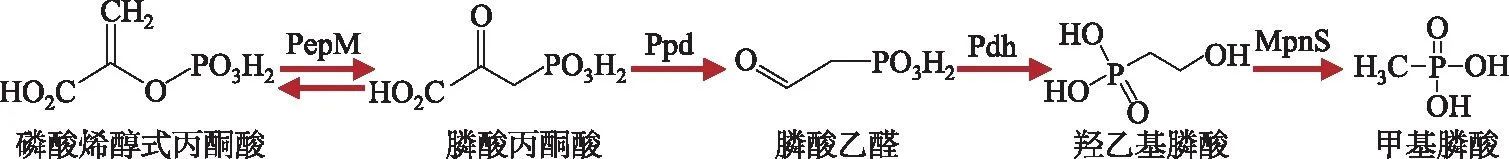
图2 甲基膦酸生物合成通路Fig.2 A biosynthetic pathway for methyl phosphonate synthesis
1.2 膦酸酯的生物利用
1963年,Zeleznick等首次发现大肠杆菌(Escherichiacoli)可分解甲基膦酸或乙基膦酸供磷[41],但当时并未阐明其利用机制。La Nauze与Rosenberg发现蜡样芽孢杆菌(Bacilluscereus)的细胞提取物可降解2-AEP[42];而Lacoste与Neuzil则进一步证实2-AEP的分解是由氨基转移与C-P键断裂两个步骤组成[43]。到目前为止,已发现三类膦酸酯生物利用途径,包括非底物特异的C-P键裂解酶(C-P lyase)途径和与底物特异的氧化裂解与水解裂解途径。其中,氧化裂解途径主要指特异性裂解2-AEP的膦酸酯加氧酶通路(phosphonate oxygenase, PhnY*Z);水解裂解途径包括膦酸丙酮酸水解酶(PnPy hydrolase, PalA)通路、膦酸乙醛水解酶(PnAA hydrolase, PhnX)通路与膦酸乙酸水解酶(phosphonoacetate hydrolase, PhnWAY)通路;上述通路的具体催化过程,在文献[7-9]中有详细描述,此处便不再赘述。
在上述膦酸酯利用途径中,C-P lyase途径是目前研究最为透彻的通路[7-9],已发现其可代谢至少25种膦酸酯底物[8]。该通路主要由膦酸酯降解(phosphonate degradation,phn)基因簇完成,phn基因簇由膦酸酯转运蛋白基因(phnC-E)、调节子基因(phnF)、C-P lyase复合体基因(phnG-M)与辅助基因(phnN-P)等组成[7-9];膦酸酯的裂解,首先由核苷酸磷酸化酶PhnI执行,在PhnGHL存在的情况下,PhnI催化膦酸基取代ATP的腺嘌呤形成三磷酸酯;后者经由磷酸水解酶PhnM作用,水解生成无机焦磷酸盐与5-磷酸核糖-1-膦酸酯;该膦酸酯再作为底物被PhnJ作用,经腺苷甲硫氨酸依赖过程裂解C-P键,并生成相应的烷烃与5-磷酸核糖-1,2-环状磷酸酯。
2 蓝藻膦酸酯利用与合成研究简史
作为水生生态系统中最重要的原核生物类群之一,蓝藻也被证实广泛参与膦酸酯的生物合成与分解代谢。相关研究成果深化了对蓝藻磷策略的认识,促进了对水体磷循环的全面理解。
2.1 蓝藻膦酸酯生物利用的研究进展
1991年,Orús与Marco对蓝藻降解敌百虫过程进行追踪,发现无菌蓝藻培养液可快速清除敌百虫;但其后续研究中发现,无细胞培养液和蓝藻培养液中的敌百虫降解情况无明显差异,因此排除了蓝藻对该膦酸酯降解的贡献[44]。Ravi则发现在磷限制条件下多变鱼腥藻(Anabaenavariabilis) ACA 101可分解草甘膦供磷,并提出可能是胞外碱性磷酸酶参与C-P键的裂解[45]。随后,Palenik等发现聚球藻(Synechococcus) WH8102可以膦酸酯为唯一的磷源维持细胞生长,并在其基因组中鉴定到了膦酸酯转运蛋白基因[46]。
在2006年,Dyhrman等在海洋固氮蓝藻红海束毛藻(Trichodesmiumerythraeum) IMS101基因组中鉴定到phn基因簇,这是首次明确蓝藻膦酸酯利用的遗传基础[10]。在上述工作中,作者比较了9株海洋蓝藻基因组(包括原绿球藻Prochlorococcus、聚球藻与鳄球藻Crocosphaera等),发现仅IMS101编码phn基因簇,提出该性状可使束毛藻在低磷条件下获得额外竞争优势;系统发育分析显示该基因簇极有可能通过水平基因转移获得,并在野外水体中鉴定到束毛藻膦酸酯转运蛋白基因(phnD)与C-P裂解酶基因(phnJ)的较高丰度表达,表明膦酸酯利用在束毛藻增殖过程中持续发挥积极作用;这一成果也开启了后续一系列蓝藻膦酸酯利用的研究[10]。表1总结了目前已报道的、经实验验证的可利用膦酸酯的蓝藻藻株。

表1 已发表的经实验验证可利用膦酸酯的蓝藻藻株汇总Tab.1 Summary of experimentally confirmed cyanobacterial strains that are capable of phosphonate utilization
2.2 蓝藻膦酸酯生物合成的研究进展
Dyhrman等发现两株红海束毛藻藻株(IMS101与ST6-5)可合成膦酸酯,其含量可占细胞总颗粒磷的8%~17%,该研究也是首次经实验确认浮游原核生物可以自主合成膦酸酯;作者认为束毛藻可能是海洋贫营养区膦酸酯的主要来源,并提出膦酸酯在海水中的积累会驱动浮游生物群落向膦酸酯利用细菌转变,进一步影响海洋碳、氮、磷循环[55]。在2022年,Acker等报道了一株原绿球藻SB可自主合成膦酸酯,且膦酸酯可占逾40%的细胞总磷;通过单细胞基因组测序与分析,作者确定膦酸酯合成功能在原绿球藻与海洋浮游细菌主要类群SAR11中广泛分布[56]。
3 基因组学助力膦酸酯代谢蓝藻的挖掘
基因组测序技术的发展带来了蓝藻基因组数据量的井喷[57-58],也为膦酸酯生物代谢与生物合成蓝藻的挖掘提供了坚实的数据基础。
3.1 基于基因组比对的蓝藻门膦酸酯利用功能调查
Willis等对9株水华蓝藻拟柱孢藻(Cylindrospermopsisraciborskii)进行基因组测序分析,发现所有藻株均编码phn基因簇[13]。同年,Teikari等对NCBI数据库中500株蓝藻基因组进行phn基因簇搜索,共发现28株蓝藻编码该基因簇,覆盖拟柱孢藻、鱼腥藻、束毛藻与节球藻(Nodularia)等常见水华蓝藻属[14]。笔者团队在前期完成中国科学院水生生物研究所淡水藻种库160株蓝藻基因组测序的基础上[58],于2020年6月结合NCBI数据库中2408个蓝藻基因组,对蓝藻门phn基因簇的分布进行了全面调查[15]。结果显示,总共31属151株蓝藻基因组编码phn基因簇(包括37株淡水藻种库蓝藻,其基因簇结构如图3所示),其中9个属为首次报道含该基因簇;后续分析蓝藻表型与C-P裂解酶基因phnJ系统发育的相关性发现,大部分(141/151=93.4%)含该基因簇的蓝藻为丝状藻,仅8株为单细胞蓝藻,且未见微囊藻或原绿球藻编码该基因簇(图4);此外,我们验证了5株含phn基因簇蓝藻(表1)利用膦酸酯的能力,确定4株蓝藻可高效分解甲基膦酸供磷,这表明蓝藻phn基因簇的编码与确切的膦酸酯利用能力存在较强相关性[15]。Acker等在构建完成海洋浮游微生物单细胞基因组测序数据集基础上,对其中蓝藻的膦酸酯生物利用途径进行调查,确定了2-AEP特异性的膦酸酯加氧酶通路(PhnY*Z)在海洋蓝藻的广泛分布[56]。

图3 中国科学院水生生物研究所淡水藻种库37株蓝藻基因组中phn基因簇结构示意图(红色字体表示该藻株同时编码参与膦酸酯生物合成的pepM与ppd基因[15]。箭头方向表示基因簇内各基因方向)Fig.3 The phn gene cluster schematics of 37 identified cyanobacterial strains from the Freshwater Algae Culture Collection at the Institute of Hydrobiology (FACHB), Chinese Academy of Sciences (strains in red font indicate their encoding of pepM and ppd genes which are responsible for phosphonate biosynthesis[15]. The arrows’ direction represent the direction of each gene in the clusters of each strain)
3.2 基于基因组比对的蓝藻门膦酸酯合成能力调查
目前,对膦酸酯合成蓝藻挖掘的研究较为缺乏。笔者团队基于上述2568个蓝藻基因组数据集,通过PepM与Ppd蛋白同源比对,确定13属81株蓝藻编码上述基因,其中7属11株藻同时编码phn基因簇(图4)。在潜在的膦酸酯合成藻株,可见念珠藻(Nostoc)、鱼腥藻、浮丝藻(Planktothrix)与眉藻(Calothrix)等蓝藻属;其中79株为丝状蓝藻,显示了丝状藻在膦酸酯代谢中的重要作用[15]。而Acker等则通过PepM蛋白保守结构“EDK (X)5NS”比对,发现原绿球藻属、节旋藻属(Arthrospira)与细鞘丝藻属(Leptolyngbya)部分藻株也编码该基因,并通过实验确定原绿球藻SB藻株可自主合成膦酸酯,进一步拓宽了膦酸酯合成蓝藻的范围[56]。

图4 151条蓝藻C-P lyase基因phnJ最大似然法系统系统发育树(以大肠杆菌菌株MG1655基因组phnJ基因为外类群。圆圈从外到内依次为:膦酸酯生物合成潜能注释、蓝藻表型注释、藻株来源生境注释[15])Fig.4 The maximum likelihood phylogenetic tree of phnJ gene sequences from 151 strains identified as carrying a phn gene cluster (Rings shown from outer to inner are: the presence of genes involved in phosphonate biosynthesis, annotation of cyanobacteria morphological subsections, and habitat classification[15])
4 蓝藻膦酸酯代谢的生态意义
膦酸酯生物利用与合成蓝藻的发现,明确了膦酸酯的蓝藻可利用性,确认了蓝藻在磷氧化还原循环中的重要作用,促进了对磷生物地球化学循环的全面认识[10]。同时,基于绝大多数蓝藻[14-15,56]与真核藻类(目前仅记录一株绿藻与一株金藻可代谢膦酸酯)[59]无法利用膦酸酯的事实,膦酸酯(包括生物源与人工源)在水体中的积累,改变了水生生态系统有机磷库的组成,也必然会驱动浮游生物竞争格局的改变,存在一定的生态风险。此外,草甘膦的大量使用已对水生生态系统产生深远影响,而部分蓝藻藻株可利用草甘膦的特性是评估这一影响的关键性状。本节同时对这一领域进展进行梳理。
4.1 蓝藻膦酸酯利用的生态意义及生态风险
4.1.1 蓝藻膦酸酯利用的生态意义 研究发现,phn基因簇的表达受低磷条件诱导,并受PhoB/R双组分系统调控[7]。因此,蓝藻膦酸酯的利用被认为是藻株对低磷生境的适应性演化[10]。当水体无机磷耗尽时,多数蓝藻可通过合成碱性磷酸酶水解胞外有机磷酸酯供磷,这也是最为人熟知的蓝藻磷策略之一[4-5]。在这种情况下,利用DOP中剩余难转化的膦酸酯部分,将赋予这些藻株额外的竞争优势,而这一策略也被证实在自然水体中广泛存在[10-15]。如Dyhrman等在3份海洋水样中的2份发现了束毛藻phnD与phnJ基因的高效表达[10];Ilikchyan等建立了基于RT-PCR的聚球藻与原绿球藻phnD定量方法,发现聚球藻相关基因的表达丰度与深度及水体磷浓度存在相关性,而原绿球藻phnD的表达量在所有样本中趋于恒定[60];Ilikchyan等同时还确定了两种藻phnD基因的表达量在马尾藻海的季节变化模式[61];Feingersch等则通过对全球海洋调查宏基因组数据集分析,确定原绿球藻是海洋表层膦酸酯生物利用的主要贡献者[62];Teikari等也观察到水华蓝藻泡沫节球藻(Nodulariaspumigena)相关基因在波罗的海的广泛表达[14]。上述结果,或许能部分解释这些蓝藻种类在相关生境的优势地位。
相比之下,淡水生境中相关研究较为缺乏。Ilikchyan等通过相同的方法在美国伊利湖(Erie Lake)等数个淡水水体检测到聚球藻phnD基因的表达[60]。笔者团队基于151条蓝藻phnJ基因数据集,分别设计了针对拟柱孢藻(图4红色进化枝)与部分念珠藻目(Nostocales,图4绿色进化枝)phnJ基因的荧光定量PCR引物,确定相关基因在广东大沙河水库的拟柱孢藻水华样品中丰度较高。同时,在已公布的25株拟柱孢藻基因组中的23株编码phn基因簇,也暗示该性状在拟柱孢藻增殖过程中的保守性与重要性[15]。
4.1.2 蓝藻膦酸酯利用的生态风险 拟柱孢藻是热带与亚热带地区最常见的淡水水华蓝藻类群。Bai等发现,尽管铜绿微囊藻(Microcystisaeruginosa) strain 205与拟柱孢藻FACHB-1096均可高效利用2-AEP,但在两株藻以2-AEP为唯一磷源共培养时,在设置的3种起始浓度下(25∶75、50∶50、75∶25),拟柱孢藻FACHB-1096均在实验结束时成为优势种[50]。笔者团队则利用转录组测序手段,对念珠藻(Nostocsp.)FACHB-892甲基膦酸利用过程进行追踪,发现甲基膦酸可使细胞从缺磷条件下恢复并维持细胞稳态;同时观察到了一个缺磷条件下被抑制的非核糖体多肽合成基因簇(常与蓝藻藻毒素合成相关),在甲基膦酸添加之后正常表达。因此,作为一种广泛存在的磷源与环境污染物,膦酸酯在驱动特定蓝藻水华暴发与藻毒素合成过程中的生态风险应当获得足够重视与深入研究[15]。
4.1.3 蓝藻草甘膦利用的生态风险 草甘膦是一种被广泛使用的广谱除草剂,其作为甘氨酸类似物,可通过抑制5-烯醇式丙酮酰莽草酸-3-磷酸合酶(5-enolpyruvyl-shikimate-3-phosphate synthase, EPSPS),导致莽草酸途径中芳香族氨基酸的生物合成受到抑制,进一步影响必需蛋白质和次生代谢产物的合成,阻碍植物和微生物的生命能量途径[36,47]。随着其大量使用,草甘膦通过直接喷洒、风流、地表径流与地面冲刷等方式进入水环境,目前已在多数水体检测到草甘膦及其代谢产物氨基甲基膦酸(aminomethylphosphonic acid,AMPA)的存在[18-24,63]。
通常观点认为,草甘膦及其衍生物对水生生态系统存在双重影响,同时作为磷源与除草剂存在。早在1991年,科学家便发现蓝藻对草甘膦的普遍耐受[64],这种抗性应与其抗草甘膦EPSPS酶结构及草甘膦代谢能力相关[45,65]。研究发现大部分蓝藻具有草甘膦耐受能力;而包括绿藻与硅藻在内的真核藻类,被证实对草甘膦敏感。如Vera等[19]与Pérez 等[21]在室外建造与自然湖泊特征形似的人工池塘,比较添加草甘膦和不添加草甘膦池塘中的淡水微生物群落结构,其实验结果表明商用草甘膦的添加会导致优势群落从对草甘膦敏感的绿藻和硅藻转变为可利用草甘膦的蓝藻(以超微蓝藻为主)。Huntscha等则发现Lake Greifensee水体中草甘膦与AMPA的快速消耗伴随着蓝藻数目的快速上升[23]。在2020年,Berman等为确定草甘膦的使用对自然水体浮游植物群落的影响,调查了南美大草原地区(使用草甘膦超过30年) 52个湖泊和巴塔哥尼亚地区(几乎没有草甘膦使用历史)24个湖泊,结果显示,持续的草甘膦使用几乎对蓝藻没有毒性作用,反而促进了可利用草甘膦的超微蓝藻生长;草甘膦的损耗和蓝藻的增长发生在相似的时间,而这一过程也对该地区蓝藻水华的发生产生了潜在影响[24]。
从表1中不难发现,蓝藻对草甘膦的可利用性与其是否编码phn基因簇关系不大。尽管目前已证实草甘膦与AMPA的生物降解主要归因于C-P lyase途径与phnWAY途径,但这并不足以解释表1中多数藻株的草甘膦利用能力。后续对这些藻株草甘膦及AMPA利用机制的全面解析,是准确评估草甘膦之于水生生态系统风险的基础与前提。
4.2 蓝藻膦酸酯代谢与水体磷循环
目前对膦酸酯生物学功能的解析尚不充分,常认为其作为修饰基团维持生物大分子结构稳定性或作为抗生素等次级代谢产物存在。如Acker等发现膦酸酯生物合成能力在海洋原绿球藻的广泛存在,同时细胞又分配高达40%的磷用于膦酸修饰基团合成,表明其参与了原绿球藻关键生物过程;进一步对膦酸酯合成蓝藻的分布特征研究发现,其相对丰度在低营养的表层水体较低并随着深度增加而上升,至营养相对丰富的深层叶绿素最大层(deep chlorophyll maximum layer)到达峰值,提出蓝藻膦酸酯的合成是在相对富营养条件下的一种生存策略;同时考虑到捕食者与病毒在该层的高丰度,作者更倾向于认为膦酸酯作为生物大分子的修饰基团,使原绿球藻抵御浮游动物捕食与病毒侵染[56]。
同时,Acker等也尝试通过实验否定膦酸酯合成作为蓝藻额外磷库的假设,其对无菌原绿球藻SB(可合成膦酸酯但无膦酸酯利用通路)磷饥饿过程中的膦酸酯含量进行追踪,发现随着磷饥饿的深入原绿球藻选择合成更多的膦酸酯而非消耗原有膦酸酯;同时,作者发现尽管膦酸酯分解代谢与合成代谢功能在原绿球藻中广泛存在,但极少有藻株同时具备这两个功能,进一步佐证了其观点[56]。上述观点更多停留在细胞生物学层面,而未从宏观的磷循环角度解释。在生物的生长过程,磷元素被用于各类有机物的合成;而当生物死亡分解时,细胞物质重新释放入环境,有机物可被生物利用进入另一轮的细胞生长,周而复始。而膦酸酯被同一类群具有分解代谢功能的部分利用,或被共生类群中具有分解代谢功能的部分利用,同样可以维持种群或共生群落的生长与增殖。比如,在我们的研究中发现,尽管同时拥有膦酸酯分解代谢与合成代谢功能的藻株数目较少(仅7属11株)[15],但我们常可在野外水体观察到分解代谢优势类群与合成代谢优势类群的共存,如拟柱孢藻属与浮丝藻属[66]、Okeania与Moorea[67]等。总的来说,蓝藻的膦酸酯利用不仅是为其自身供磷的过程,还维持了活跃的磷氧化还原循环,对生态系统的能量流动与物质循环有重要意义。因此,蓝藻膦酸酯生物合成驱动的水体有机磷库组成与浮游生物群落变化,以及这一过程对水体磷循环的影响值得进一步探究。
5 蓝藻甲基膦酸代谢与水体好氧甲烷发生的潜在关系
在所有蓝藻膦酸酯代谢生态风险中,最受关注的则是甲基膦酸代谢对水体好氧甲烷发生的潜在贡献。水生生态系统贡献了全球近一半的甲烷排放,对其内部甲烷发生机制的全面解析是实现“双碳”目标的关键一环[68]。通常认为,甲烷的生物发生是基于产甲烷古菌 (细菌)在严格厌氧条件下完成,甲烷在水体厌氧底层产生之后向上迁移,此过程伴随着甲烷的微生物氧化,其浓度应从底层到表层逐渐降低[69-71]。但野外观察发现,水体次表层存在甲烷过饱和现象,这一现象被称为甲烷悖论(methane paradox),暗示该水层存在好氧的甲烷发生过程[25-26,72-78]。尽管目前该过程对全球甲烷排放的具体贡献仍存在争议[79-81],但好氧甲烷发生又是切实发生并被频繁观察到的[25-26,72-79]。目前已被证实的好氧甲烷发生机制包括可溶性有机物的光催化产甲烷[81-83]、特殊底物的生物脱甲基作用产甲烷[25-26,74]、藻类光合[75,84]和固氮作用[85]等。其中蓝藻的甲基膦酸利用也被认为是水体好氧甲烷发生的可能来源之一[25-26,74]。
Karl等发现,添加甲基膦酸的海洋原水可高效产生甲烷。通过培养实验证实束毛藻在海水代谢甲基膦酸产甲烷过程中发挥重要作用[25]。Teikari等对两株泡沫节球藻进行甲烷排放模式研究,发现两株蓝藻均可高效产生甲烷,并确定其积累模式为线性[14]。笔者团队对3株蓝藻的甲基膦酸代谢过程进行追踪,确定在整个生长周期体系均可高效代谢甲基膦酸释放甲烷,同时正磷酸盐的添加可大幅降低甲烷排放速率。而模拟12 h∶12 h昼夜比的36 h甲烷积累实验显示,产甲烷多集中于光周期,在夜晚则减弱。上述结果与已报道的多个好氧甲烷发生特性一致,暗示了蓝藻的膦酸酯利用对水体好氧甲烷发生的贡献[15]。
表2总结了部分已发表的好氧甲烷发生研究中对蓝藻参与度的评述。从表中不难发现,不同文献中的主要结论可能存在矛盾,这也体现了好氧甲烷发生机制的多样性与复杂性。总的来说,水生生态系统中好氧甲烷的发生应该是多种生物参与、多机制的整合结果,这些机制随着水体类型的不同、环境因素的差异会出现不同的组合变化。而基于蓝藻的膦酸酯利用在这一过程中持续发挥着积极的作用。对该过程甲烷排放模式与通量的具体解析将有助于水生生态系统产甲烷机制的全面认识。另一方面,全球气候变化被认为是蓝藻水华暴发频次与强度增加的驱动因子之一,而如果水华蓝藻可以通过代谢水体中广泛存在的甲基膦酸产生甲烷,那么这一过程必将正反馈于全球变暖[1-2]。因此对这一过程的持续追踪不仅有助于解析蓝藻水华发生机理,也将推动蓝藻水华生态风险的全面评估。

表2 部分已发表的好氧甲烷发生研究中对蓝藻参与度评述Tab.2 Participation of cyanobacteria in published filed observations on oxic methane production
6 研究展望
生物利用膦酸酯的生态意义不仅局限于磷循环:如Gomez-Garcia等[12]发现,聚球藻代谢甲基膦酸产生的甲烷,可被藻细胞用作碳源以维持生长,从而参与碳循环;Chin等[86]发现了一条非磷限制依赖的2-AEP特异性利用通路,显示该膦酸酯可能作为氮源维持生物生长;在2021年,Murphy等[87]则发现一个非氮磷浓度依赖型2-AEP特异性转运蛋白复合体,提出2-AEP是可能水体初级生产者重要的氮源与磷源。因此,后续研究应重点关注蓝藻膦酸酯代谢新通路的挖掘与验证,以最终确定蓝藻膦酸酯(包括人工合成源与生物源)代谢的生态意义与生态风险。
此外,藻际细菌可能在蓝藻膦酸酯代谢过程中发挥重要作用。藻际环境是藻菌关系发生的主要界面,其定义为浮游植物细胞周围富含其分泌的有机基质的区域,是水生细菌的重要微环境[88]。研究发现蓝藻与藻际细菌间存在多方面的功能互补,藻际细菌在蓝藻生长及水华生消过程中发挥重要作用;同时蓝藻可根据自身需求,选择性招募藻际细菌[89-91]。而膦酸酯利用细菌在野外水体广泛存在,如Villarreal-Chiu等基于功能基因比对,发现近16%细菌基因组编码phn基因簇,而40%细菌基因组中存在一种或多种膦酸酯生物利用途径[11]。在2011年,Saxton等对草甘膦污染水体膦酸酯代谢途径进行分析,也发现浮游细菌而非蓝藻是草甘膦降解的主要执行者;其进一步提出在自然环境中微生物以群落行使功能,而群落的集合功能(the collective ability of the community)远比个体代谢能力重要[18]。此外,我们在实验中发现膦酸酯利用藻际细菌在微囊藻水华生物样本与实验室纯培养蓝藻藻际中广泛存在(笔者团队未发表数据)。因此,对膦酸酯利用藻际细菌在蓝藻,尤其是不编码膦酸酯代谢通路蓝藻的膦酸酯利用过程中作用的研究,将有助于全面理解蓝藻膦酸酯利用的确切机制及其对磷循环的具体贡献。
