城镇化背景下深圳典型流域鱼类群落特征及驱动因子
2023-01-13王伟民陈宇顺
屈 霄,刘 晗,2,阳 敏,2,辛 未,王伟民,陈宇顺,2,*
1 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,武汉 430072 2 中国科学院大学,北京 100049 3 深圳市环境监测中心站,深圳 518049
自工业革命以来全球范围内城镇化进程不断加快,范围急速扩张,城镇化已经成为社会经济发展与现代文明的重要标志。特别地,自20世纪90年代来我国经历了世界上规模最大的城镇化过程,城镇化率由19.4%迅速攀升至2020年的63.89%[1]。城镇化进程中,土地类型的改变、生产生活方式的转变和人类活动干扰频次的增加,均会对区域各类生态系统产生深远的影响[2—3]。就水生态系统而言,城镇化能够通过影响水体物理和化学特征及生物间相互作用,进而影响到水生态系统结构与功能,主要表现为水质污染、水生生物多样性降低、群落结构改变、生态系统服务功能下降等[2—6]。
鱼类作为水生态系统能量流动和物质循环的顶层生物,其群落组成及分布特征往往是对水生态系统变化的适应表现。城镇化对鱼类自然栖息地的改造,使得自然连续的栖息地加速演变成为破碎斑块状栖息地,并因“不透水表面”的增加,自然水流流态也随之发生改变[5,7]。Brown等定量分析了美国九大城市圈城镇化与鱼类群落结构间的相关关系,结果发现不同程度的城镇化对鱼类群落结构有着显著影响[2]。由城镇化引起的水质污染和栖息生境的改变,不仅能够造成土著敏感鱼类种群数量下降甚至消失[8],还为外来耐受性鱼类的定殖和扩散提供了适宜的条件[9,10],进而使得鱼类区系同质化愈发明显[11—12]。
自改革开放以来,深圳作为我国经济发展的排头兵,城镇化建设取得长足发展,市域面积由327.5km2增长到1997km2,常住人口由31万人增长到1343万人[13]。荀斌等分析了1980年至2010年深圳城市扩展模式,发现30年间深圳土地利用结构发生巨大变化,城市用地面积迅速增加,景观破碎化程度增加,形状趋于复杂化[14]。在深圳快速城市化过程中,空间扩张和资源环境的不合理利用给深圳城市生态系统带来深刻影响,如生态系统生产力降低、暴雨洪水灾害、环境污染、城市热岛效应等[15—17]。然而,城市域内各式生产生活的痕迹都终将通过径流汇集到河流和湖泊之中,深圳市水生态系统遭受着前所未有的胁迫,水生生物的栖息环境不断恶化[18—20]。
现阶段有关于深圳水生态环境的研究仍多关注于水体理化指标的状况,对水生生物特别是鱼类的研究依旧非常缺乏。同时,深圳为典型的快速城镇化城市,为研究城镇化对鱼类群落的影响提供了良好的素材。因此,本研究着眼于水生生物群落如何响应快速城镇化引发的河流生境变化这一关键科学问题,选取深圳市域内处于不同城镇化程度的两个典型流域,从鱼类群落结构着手,探究城镇化对鱼类群落及多样性的生态学效应及作用机制,以期为高度城镇化流域水生态系统保护和生物多样性维持提供科学依据及理论参考。
1 材料与方法
1.1 研究区域与样点布设
深圳地处广东省南部,地势东南高,西北低,以平原和台地地形为主,属于亚热带海洋性季风气候,雨量充沛但时空分布不均。域内土地覆盖总体表现为东部以林地覆盖为主,中部与西部以建设用地覆盖为主。流域面积大于100 km2的河流有5条,即深圳河、观澜河、茅洲河、龙岗河、坪山河,其中观澜河和坪山河流域均为东江水系支流。观澜河为东江水系石马河的上游段,在企坪断面以下进入东莞市境内,深莞交界断面以上流域面积189.3 km2,干流长20 km,河流平均比降2.12‰。深圳域内以观澜河流域建设用地面积占比最高,为63.55%;其次为林地32.89%。流域内总体工业发达、人口密集。观澜河流域内河流均属雨源性河流,河流水环境承载能力小、自净能力弱。因此,由于流域内工业及生活污水的注入,观澜河污染相对较严重。坪山河为东江水系淡水河一级支流,深圳市内流域面积129.4 km2,干流长25 km,河流平均比降为9.34‰。坪山河流域土地覆盖以林地为主,占流域面积57.43%,其次为建筑用地34.34%。坪山河流域南侧地势较高,以丘陵和低山为主,支流众多,环境保持较好;北侧地势较低,以台地和平原为主,支流较少。坪山河流域河流大致可划分为山区型河流和城镇型河流两种类型,山区型河流自然环境相对较好,污染小、人类活动干扰少;城镇型河流沿河排污众多,污水超过河流自净能力,污染相对较重。
本研究在观澜河流域和坪山河流域,沿上游至下游的纵向梯度布设监测样点。样点布设原则为:(1)样点的空间分布确保样点覆盖流域水系,并且平衡不同大小类别的河流的采样点数量;(2)样点避开较明显的人类活动干扰点(如排污口、水坝等);(3)样点包括或接近深圳市水文水质监测站点。因此,共在观澜河流域实际布设监测样点13个,在坪山河流域实际布设样点11个(图1)。
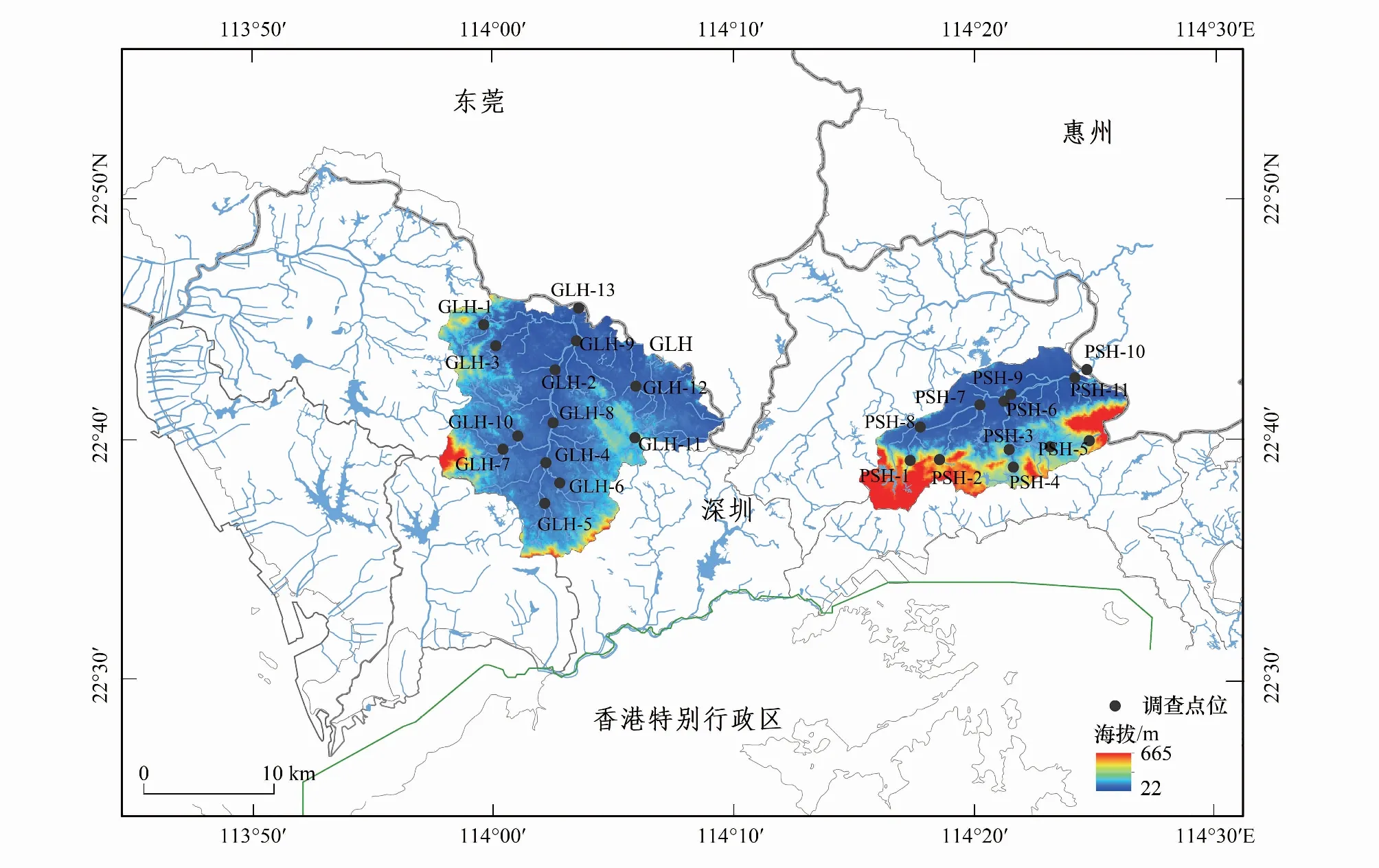
图1 观澜河和坪山河流域鱼类群落调查区域与样点分布Fig.1 Study area and sampling sites in the Guanlan River and Pingshan River Basins
1.2 样品采集
分别在2019年8月(丰水期)和11月(枯水期)对观澜河流域(13个样点)和坪山河流域(11个样点)共24个样点进行野外调查和鱼类样品采集。通过在每样点上下游100m河段设置拦网边界,使用小型便携式脉冲捕鱼仪(FLW-X388000w,12V,30—120Hz)在该河段拦网边界内各类生境中反复作业30min进行鱼类样品采集。鱼类取样时遵循够用即可,尽量少捕的原则,如渔获物总重较多则在记录总重后,随机取一定重量样品进行测量,其余鱼类样本均当场放生。采集后的标本在新鲜状态下进行鉴定,统计每种鱼类数量和重量。疑难种以甲醛进行固定后带回实验室进一步鉴定。在保证各河段采样强度和时间一致的情况下,将每河段采集到的鱼类尾数和重量分别表征为该河段鱼类的丰度和生物量。
由于监测样点包括或接近深圳市水文水质监测站点,选择与鱼类采样同一时期、采样站点重叠或最接近的深圳市水质监测 (依据生态环境部水质监测与分析标准方法)样点的数据,包括水深(WD, m)、水温(WT, ℃)、透明度(SD, cm)、pH值(pH)、电导率(COND, μS/cm)、溶解氧(DO, mg/L)、浊度(NTU)、流速(FLOW, m/s)、河宽(RW, m)、悬浮物(SSD, mg/L)、高锰酸盐指数(Pe, mg/L)、化学需氧量(COD, mg/L)、五日生化需氧量(BOD, mg/L)、总氮(TN, mg/L)、总磷(TP, mg/L)、氨氮(NH4-N, mg/L)等指标。
选取每样点上游2km距离建立500m缓冲区,计算缓冲区内各类土地类型覆盖面积及比例以表征该样点土地利用方式。以SPOT遥感卫星影像作为数据源,提取2019年8月—9月云量较少、成像清晰度高的研究区域影像。对获取的影像使用ENVI 5.3软件进行辐射定标和FLAASH大气校正预处理。预处理后的影像采用分类树回归算法(CART)进行土地利用类型监督分类提取。土地利用类型分为以下几类:城镇用地、道路、其他不透水、林地、草地、裸地、水域。
1.3 数据分析
生物多样性的计算采用种类丰富度、Shannon-Wiener多样性、Margalef丰富度、Simpson优势度等指数。采用相对重要性指数(IRI)确定各流域优势种类[21]。应用双因素方差分析(Two-way ANOVAs)检验季节、流域及其交互作用对鱼类丰度、生物量、物种丰富度、Shannon-Wiener多样性、Margalef丰富度、Simpson优势度指数的影响。如果流域与季节存在交互作用,则对试验组差异分别进行单因素方差分析(One-way ANOVA)。如果单因素方差差异显著,则进行多重比较;如果方差具有同质性,就进行LSD检验;如果方差不具同质性,就选用非参数Games-Howell检验。为了减少“双零效应”影响,采用基于Bray-curtis距离的冗余分析模型(dbRDA)分析群落结构对环境因子的响应。在进行dbRDA分析进行前,将环境变量(pH值除外)进行log10(x+1)转换,并去除方差膨胀因子(VIF)超过20的变量,以避免变量之间存在高度共线性。环境变量的重要值按其单独解释物种数据的方差值大小排序,其解释的显著性由Monte Carlo 检验确定。以上统计分析均使用“vegan”程序包在R 3.5.1软件中完成。
2 结果与分析
2.1 种类组成及生物量分布
2019年在观澜河流域和坪山河流域共采集到鱼类3184尾,53.38kg,共31种,隶属于5目13科27属(表1)。所获鱼类以鲤科鱼类为主,占所有种类38.7%,其次为鳅科(12.9%)、平鳍鳅科(9.7%)。其中,观澜河流域共采集到鱼类18种,以鲤科6种最多,其次为慈鲷科3种,平鳍鳅科和鲇科种类均未采集到;坪山河流域共采集到鱼类26种,鲤科11种最多,其次为鳅科和平鳍鳅科种类各3种,合鳃鱼科和攀鲈科种类均未采集到。丰水期在观澜河流域和坪山河流域共采集到鱼类26种,其中观澜河流域鱼类15种,坪山河流域鱼类19种。总体上枯水期鱼类种类稍少于丰水期,共采集到鱼类24种,观澜河12种,坪山河21种。

表1 观澜河流域和坪山河流域鱼类种类组成
双因素方差分析发现仅不同季节对鱼类丰度有显著性影响,而不同流域和季节间交互作用并无显著性影响(表2)。整体上,枯水期鱼类丰度显著高于丰水期鱼类丰度(P<0.05),观澜河枯水期鱼类丰度为(85.4±68.5)尾,显著高于丰水期(34.9±42.1)尾;而坪山河流域丰水期鱼类丰度(52.3±39.5)尾与枯水期(95.0±68.0)尾并不显著。就流域而言,丰水期和枯水期坪山河流域平均鱼类丰度(73.6±58.5)尾,均高于观澜河平均鱼类丰度(60.2±61.3)尾,但没有显著性差异(图2)。季节、流域及其交互作用虽对鱼类生物量无显著性影响,但鱼类生物量呈现出与丰度相似的趋势,观澜河和坪山河流域枯水期生物量同样高于丰水期生物量,但没有显著性差异。与鱼类丰度相反的是,观澜河流域平均鱼类生物量(1320.9±988.1)克,高于坪山河流域平均鱼类生物量(865.6±760.1)克(图2)。
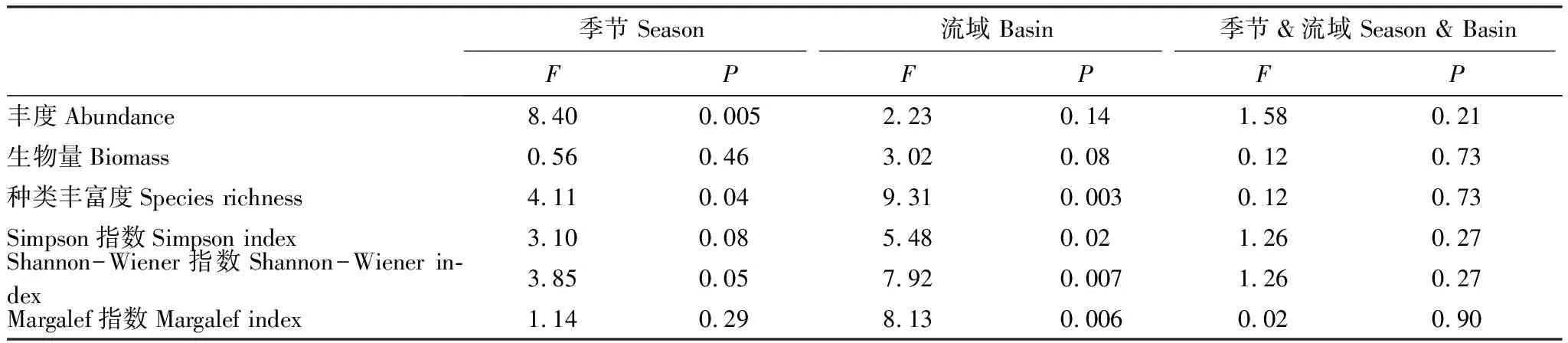
表2 季节与流域间的鱼类群落双因素方差分析
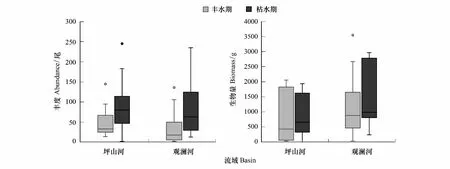
图2 观澜河流域和坪山河流域鱼类丰度和生物量Fig.2 Fish abundance and biomass in the Guanlan River and Pingshan River Basins
2.2 优势种及生物多样性
以相对重要性指数(IRI)衡量观澜河流域和坪山河流域鱼类优势种类,发现外来种尼罗罗非鱼、尖齿胡鲶、齐氏罗非鱼在观澜河和坪山河流域均已成为了主要优势类群,其中观澜河流域鱼类优势种基本以外来鱼类为主(表3)。坪山河流域不同点位间鱼类优势种差别较大,在城镇点位中鱼类优势种以尼罗罗非鱼、尖齿胡鲶、齐氏罗非鱼等外来入侵鱼类为主;而在山区点位中土著鱼类宽鳍鱲、异鱲、子陵吻鰕虎鱼等优势度相对较高。不同时期间,观澜河和坪山河流域鱼类优势类群变化不大。从监测样点外来种与土著种占比来看,仅在坪山河山区部分点位以土著种为主,外来入侵种比例较小;坪山河城镇点位和观澜河流域外来入侵种丰度平均占比达92.5%(图3)。
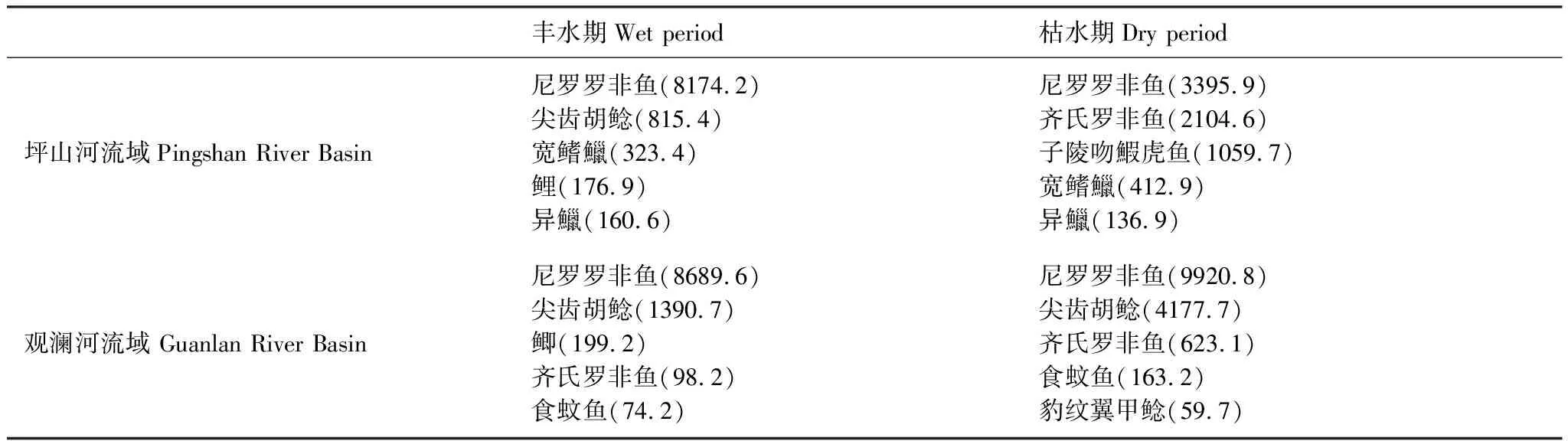
表3 观澜河流域和坪山河流域鱼类相对重要性指数
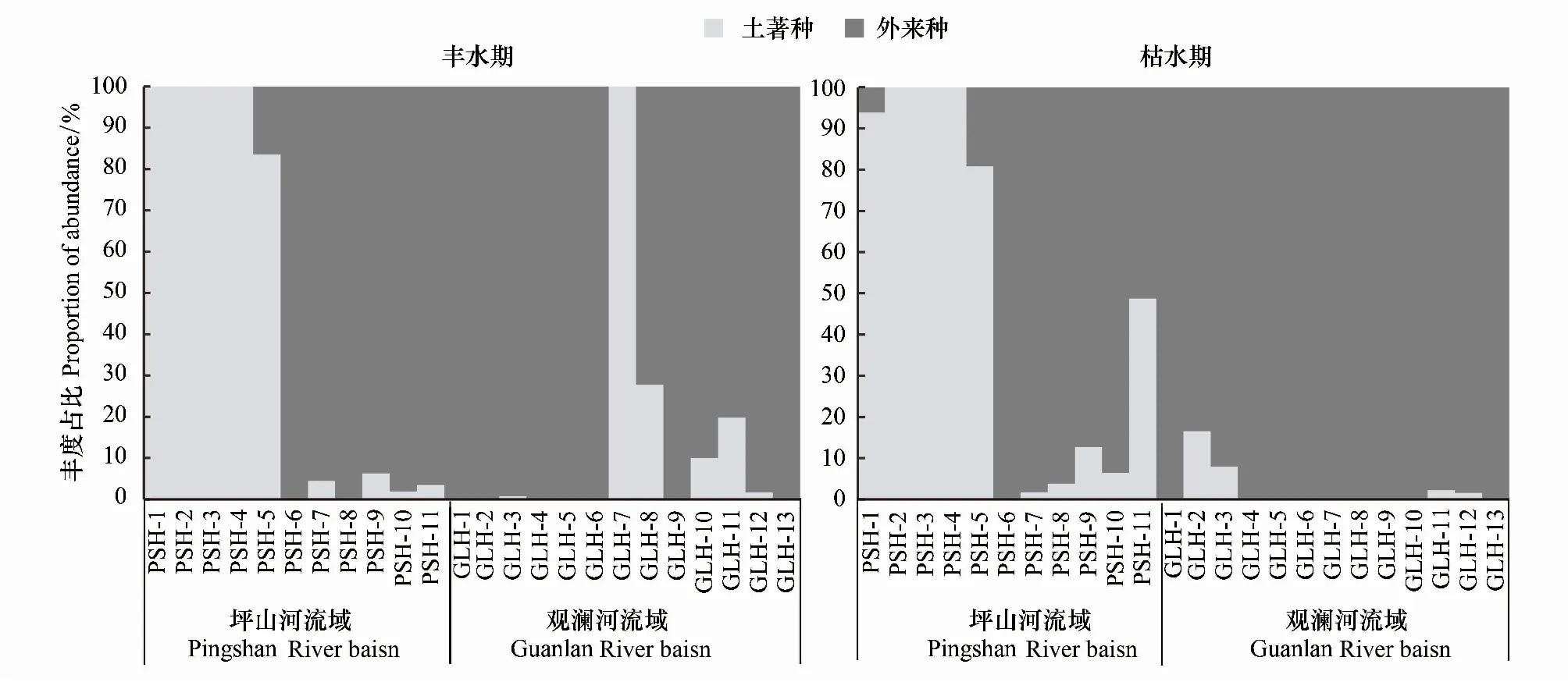
图3 不同水期观澜河流域和坪山河流域外来种与土著种鱼类丰度占比Fig.3 In wet period and dry period proportion of abundance of exotic and indigenous species in the Guanlan River and Pingshan River Basins
在生物多样性方面,不同季节和流域均对种类丰富度有显著影响,其交互作用无显著影响(表2)。具体而言,坪山河流域丰水期和枯水期种类丰富度指数均显著高于观澜河流域(P<0.05),而同一流域内枯水期种类丰富度指数高于丰水期,但不存在显著性差异(图4)。整体而言,观澜河和坪山河流域Shannon指数偏低(0—1.6),不同季节和流域对其有显著性影响;枯水期平均Shannon指数(0.73)稍高于丰水期(0.49);坪山河流域平均Shannon指数(0.77)高于观澜河流域(0.45)。仅枯水期坪山河流域Shannon指数(0.95±0.48)显著高于观澜河流域(0.51±0.32)(P<0.05)(图4)。Margalef指数与种类丰富度指数变动趋势较为类似,坪山河流域丰水期Margalef指数(0.72±0.35)和枯水期(0.87±0.46)均显著高于观澜河流域(丰水期,0.41±0.43;枯水期,0.52±0.35)(图4)。观澜河和坪山河流域Simpson优势度指数均相对较低,物种组成较不均匀。枯水期平均Simpson指数(0.38)稍高于丰水期(0.27)。在季节、流域及其交互作用中,仅不同流域对Simpson指数有显著影响,具体为枯水期坪山河流域Simpson指数(0.49±0.23)显著高于观澜河流域(0.27±18)(P<0.05)(图4)。
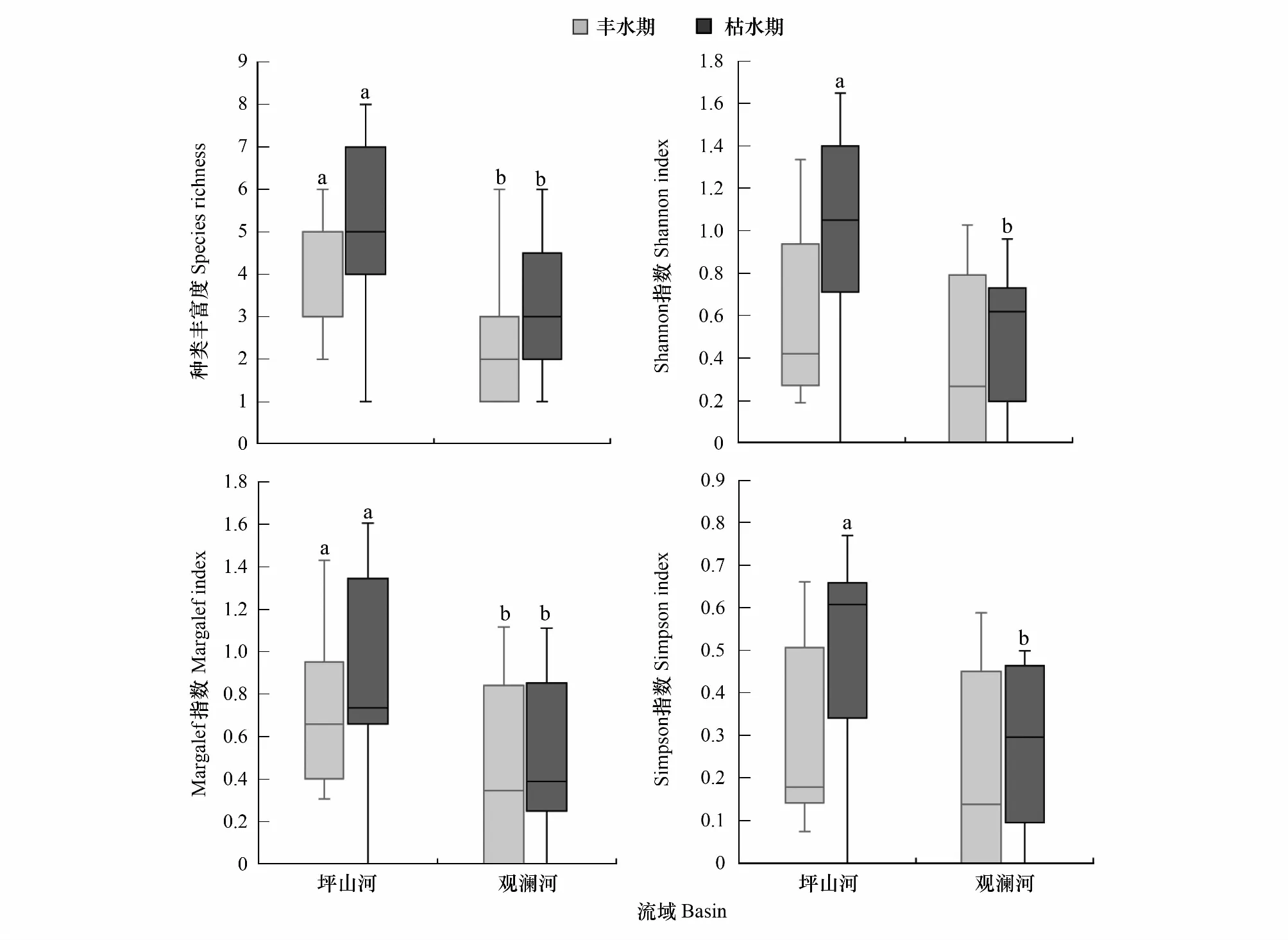
图4 观澜河流域和坪山河流域鱼类多样性指数:种类丰富度指数,香浓-维纳多样性指数,马格列夫丰富度指数,辛普森优势度指数Fig.4 Fish diversity: index of species richness, Shannon index, Margalef index, Simpson index in the Guanlan River and Pingshan River Basins箱型图上边缘不同小写字母表示流域间有显著性差异
2.3 鱼类群落与环境因子相关关系
观澜河流域与坪山河流域环境因子的差异主要表现在建设用地占比、高锰酸盐指数、化学需氧量、生化需氧量、营养盐等方面的差异(表4)。具体而言,观澜河流域样点缓冲区内建筑占比、道路占比均显著高于坪山河流域(P<0.05);相应的坪山河流域林地占比则显著高于观澜河流域(P<0.05)。在河流物理化学性质方面,观澜河流域物理属性如河宽、水深、流速等,与坪山河流域无显著性差异,而水体理化性质总磷、总氮、氨氮、化学需氧量、生化需氧量、高锰酸盐指数等,观澜河流域显著高于坪山河流域(P<0.05),为其1.5—9.0倍。不同水期间,除水温、河宽、流速外,观澜河和坪山河流域河流物化性质变动相对较小。
丰水期鱼类丰度与环境因子db-RDA分析结果显示(图5),全模型检验显著(P=0.005),第一、二主成分共解释了物种丰度累计方差的51.2%。河宽(0.48)、建筑用地占比(0.48)、溶解氧(-0.47)、浊度(0.46)与第一轴相关性较高;而道路占比(0.67)、总磷(0.60)、pH(0.55)、高锰酸盐指数(0.54)、水温(0.51)、化学需氧量(0.50)与第二轴相关性高。对各环境因子进行999次Monte Carlo模型检验发现,建筑用地占比为影响观澜河流域和坪山河流域鱼类丰度格局的关键环境因子(F=2.60,P=0.003)。枯水期第一、二主成分共解释了物种丰度累计方差的57.6%,全模型检验P值为0.012,模型验证通过(图5)。总氮(0.73)、浊度(0.64)、建筑用地占比(0.64)、河宽(0.61)、道路占比(0.55)、水温(0.52)、林地占比(-0.52)与第一轴相关系数较高;对第二轴贡献率较高的环境因子有:生化需氧量(0.64)、道路占比(0.56)、林地占比(-0.45)等。Monte Carlo模型检验发现,总氮为影响枯水期鱼类丰度分布的显著性因子(F=4.27,P=0.001)。
对丰水期鱼类生物量应用db-RDA分析发现,第一、二轴共解释了环境—物种变异的48.1%,全模型验证P值为0.009,模型验证通过(图5)。与轴一较相关的环境因子有:建筑用地占比(0.54)、河宽,0.48)、总氮(0.45)、溶解氧(-0.44);与轴二较相关的环境因子为总磷(0.51)、道路占比(0.50)、化学需氧量(0.48)、林地占比(-0.47)。对各环境因子进行999次Monte Carlo模型检验发现pH(F=2.24,P=0.006)、建筑用地占比(F=1.63,P=0.042)为影响丰水期鱼类生物量组成的关键环境因子。而枯水期,第一、二主成分共解释了观澜河和坪山河流域鱼类生物量组成累计方差的54.9%,全模型验证P值为0.048,模型验证通过(图5)。道路占比(0.77)、林地占比(-0.75)、建筑用地占比(0.64)、生化需氧量(0.62)、总氮(0.57)对轴一的贡献率较高;浊度(0.69)、总氮(0.55)对轴二的贡献率较高。Monte Carlo模型检验发现,道路占比为影响枯水期鱼类生物量分布的显著性因子(F=3.51,P=0.001)。
3 讨论
3.1 鱼类群落结构变动及主要驱动因素
观澜河流域与坪山河流域鱼类群落结构的差异主要表现在种类组成、优势类群和生物多样性的不同。具
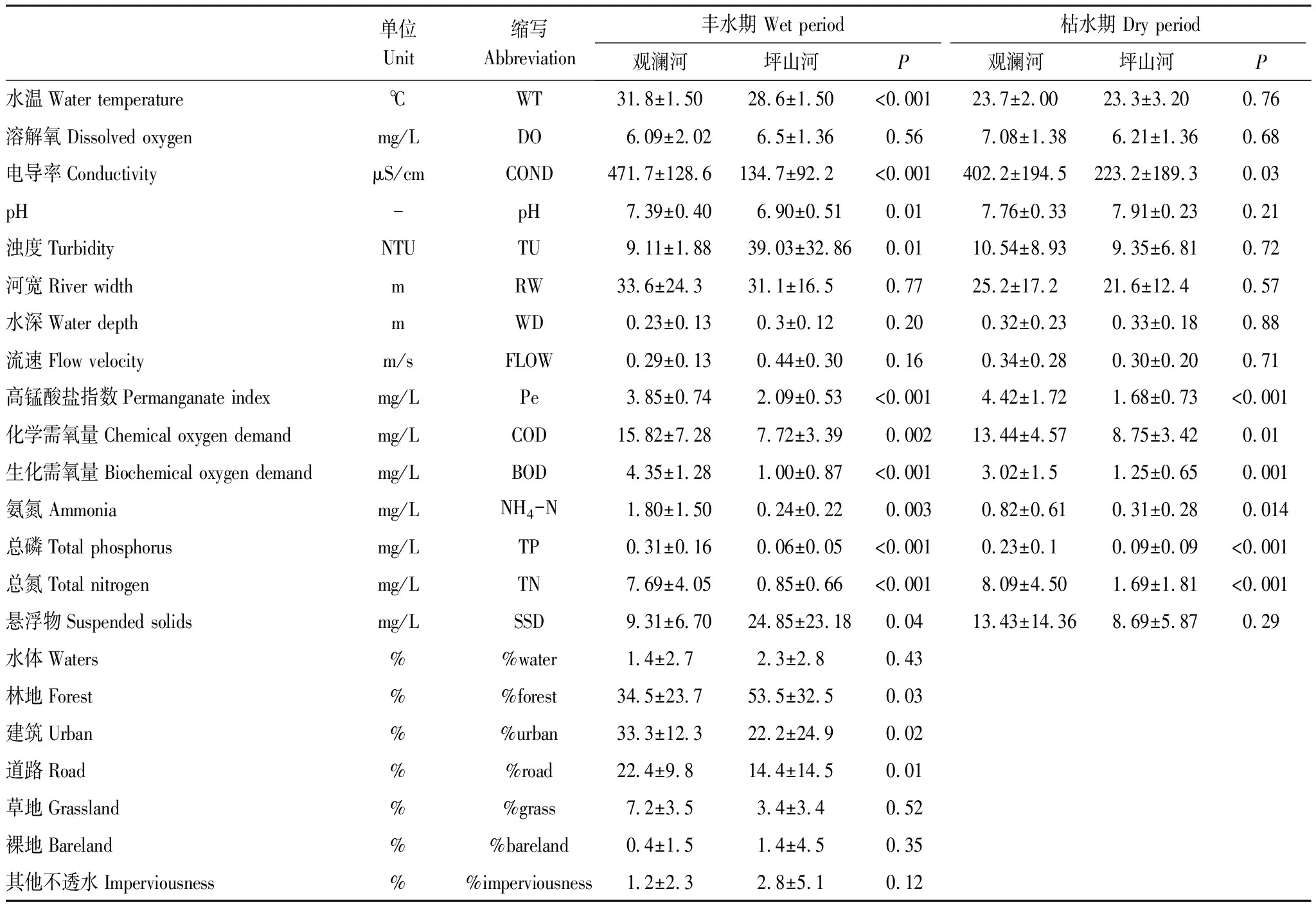
表4 观澜河流域和坪山河流域环境因子
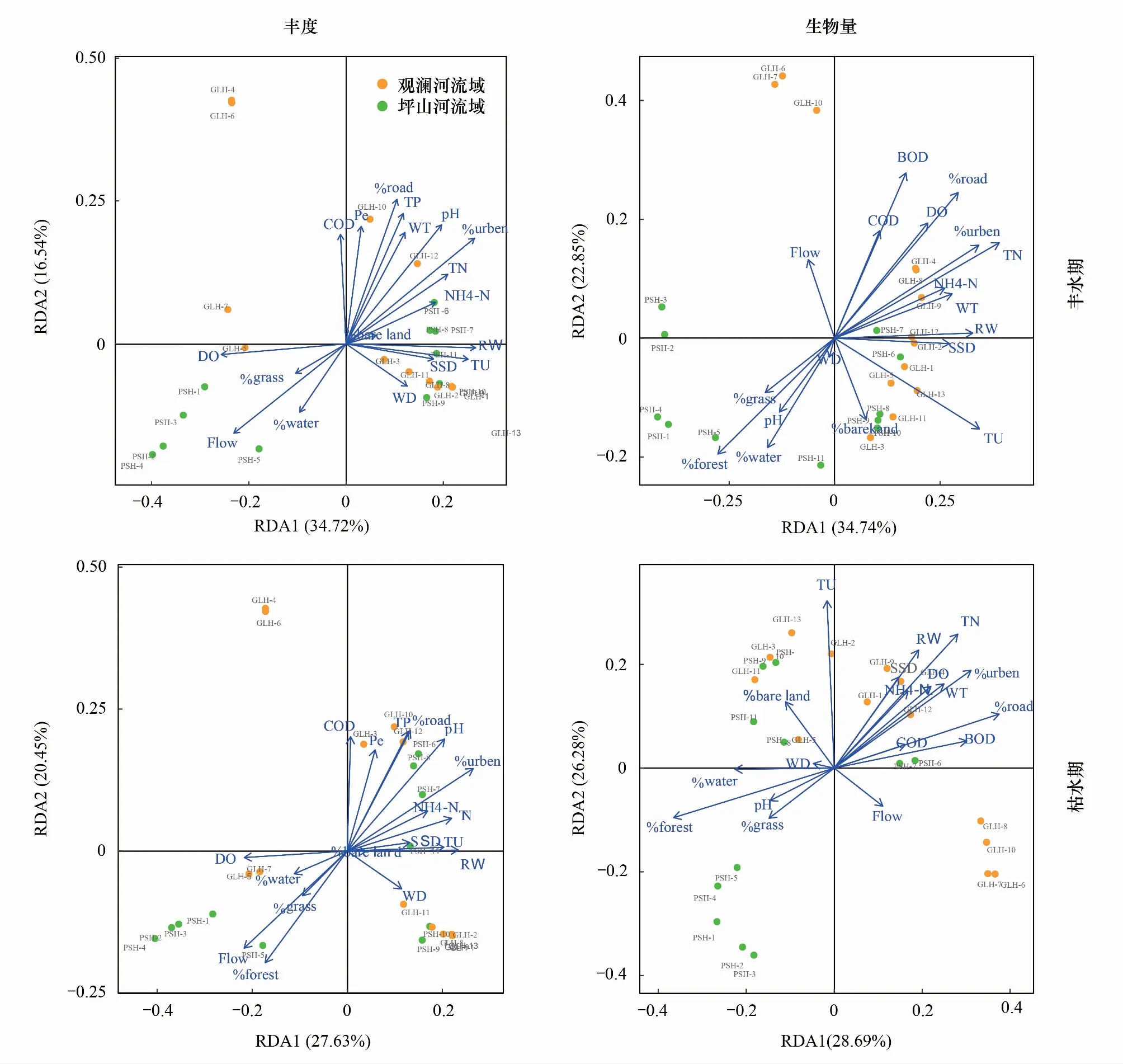
图5 不同水期观澜河流域和坪山河流域鱼类丰度和生物量与环境因子基于距离的冗余分析Fig.5 Distance-based redundancy analysis between fish abundance, biomass in dry season in the Guanlan River and Pingshan River Basins and environmental factorsWT: 水温Water Temperature; DO: 溶解氧Dissolved Oxygen; COND: 电导率Conductivity; TU: 浊度Turbidity; RW: 河宽River Width; WD: 水深Water Depth; FLOW: 流速Flow Velocity; Pe: 高锰酸盐指数Permanganate Index; COD: 化学需氧量Chemical Oxygen Demand; BOD: 生化需氧量Biochemical Oxygen Demand; NH4-N: 氨氮Ammonia; TP: 总磷Total Phosphorus; TN: 总氮Total nitrogen; SSD: 悬浮物Suspended Solids; %water: 水体Waters; %forest: 林地Forest; %urban: 建筑Urban; %road: 道路Road; %grass: 草地Grassland; %bareland: 裸地Bareland; %imperviousness: 其他不透水Imperviousness.
体而言,观澜河流域种类组成上以耐污广布种和外来种为主,如鲤Cyprinuscarpio、鲫Carassiusauratus、泥鳅Misgurnusanguillicaudatus、尼罗罗非鱼Oreochromisniloticus、尖齿胡鲶(又名革胡子鲶)Clariasgariepinus、豹纹意甲鲶Pterygoplichthyspardalis等;而坪山河流域鱼类组成上则以土著敏感种类为主,如异鱲Parazaccospilurus、横纹南鳅Schisturafasciolatus、越鲇Siluruscochinchinensis、麦氏拟腹吸鳅Pseudogastromyzonmyersi等。Scott研究了北美36个不同城镇化程度流域的鱼类群落结构,发现广布耐污鱼类更能适应因城镇化所引发的栖息地特征改变,而土著敏感鱼类较少能适应这一变化[8]。城镇化作为主要的人类活动方式之一,其在发展进程中对原有河流栖境进行一系列改造,出现了诸如生产生活排放的污水影响水体水质、“不透水面积”促使河流原有水文情势发生改变等问题[22—23]。国内外关于城镇化对鱼类群落结构影响的研究,均发现城镇化对资源的开发利用改变了原有栖息地特征,使得土著敏感性鱼类逐渐被广布耐受性鱼类所替代,改变了鱼类区系特征[8,24—27]。本研究的结果也同样验证了这一观点,这种现象在坪山河流域内部城镇河网点位和山区河网点位鱼类组成差异上更为明显。同时,环境因子与鱼类群落组成的冗余分析也发现,驱动观澜河流域和坪山河流域鱼类群落差异的关键环境因子为建筑用地占比、道路占比、总氮等,进一步验证了城镇化对鱼类群落的影响。冗余分析也显示出建筑用地占比、道路占比等表征城镇化的指标与水体生化需氧量、化学需氧量、总氮、总磷、氨氮等有着明显的正相关关系。水体理化因子的单因素方差分析同样表明,观澜河流域水体总磷、总氮、氨氮、化学需氧量、生化需氧量、高锰酸盐指数等含量显著高于坪山河流域,为坪山河流域的数倍之多。高度城镇化及其附带其他人类活动已经对观澜河流域河流理化性质造成了较大影响。
同时,值得注意的是,在观澜河流域和坪山河流域城镇河网点位中外来鱼类尼罗罗非鱼、齐氏罗非鱼、尖齿胡鲶、豹纹意甲鲶、食蚊鱼已经逐渐成为了优势类群,与土著鱼类竞争生态位和生存空间。近期调查研究表明,外来鱼类已经在广东、广西主要河流内成功建立稳定种群,并逐步向珠江全流域扩散,甚至在海拔相对较高的南盘江流域也有出现[28—30]。罗非鱼和尖齿胡鲶作为养殖品种、豹纹意甲鲶作为观赏鱼类、食蚊鱼为控制孑孓被先后引入国内,因其生长快、繁殖力强、适应性强等特点,较土著鱼类生存能力更强[31—32]。由于外来入侵鱼类适应能力强,缺少天敌,极易在新栖息地中爆发性增殖,减少土著种类生存空间,使入侵水域生物多样性丧失,进而破坏生态系统稳定性,改变其原有结构和功能[33—34]。在本研究两次调查中观澜河流域部分点位如沙元埔、坂田河、大水坑等,仅能够采集到以尼罗罗非鱼为主的外来物种,土著种已经很难采集到。
3.2 基于鱼类多样性的城镇化河流管理与保护建议
河流作为自然资源的载体,也是水陆生态系统物质交换和能量转化的重要纽带,为人类提供了重要的生态系统服务,如供水、调蓄洪水、休闲、生物多样性等。随着城镇化进程的扩张和加速,越来越多的自然河流将演变成为受人类活动干扰的城镇河流。基于本研究结果发现,城镇化的建设与开发改变了流域土地利用性质、增加了河流水体营养盐和还原性物质量,从而直接作用于原有土著鱼类栖息生境,造成土著敏感鱼类消失,生物多样性下降,并为外来物种成功入侵提供了便利条件。鱼类作为水生态系统能量流动和物质循环的顶层组分,其群落的结构性变化往往会对水生态系统稳定与功能产生影响。因此,建议城镇河流水生态健康管理与保护的重点包括以下几个方面:(1)以土著鱼类多样性恢复为目标,提高河流生物多样性;(2)河流治理应注重生态功能的恢复,特别是适宜水生生物生存的生境条件的提升;(3)实施河流分类分区管理与保护,制定“一河一策”保护与治理规划;(4)推动生态环境管理由水质目标管理向水生态目标管理转变,建立完善的、长期运行的区域水生态监测体系和网络。
