大麦种质资源表型性状遗传差异分析
2023-01-10马艳明赵连佳安学春
马艳明,赵连佳,王 威,安学春,向 莉,苗 雨
(1.新疆农业科学院农作物品种资源研究所,乌鲁木齐 830091;2.新疆巴里坤哈萨克自治县农业农机技术推广中心,新疆巴里坤 839200;3.新疆农业科学院奇台麦类试验站,新疆奇台 830052)
0 引 言
【研究意义】大麦是世界上种植面积和产量位于小麦、水稻和玉米之后第4位的禾谷类作物[1],在我国主要分布在长江流域、黄河流域和青藏高原[2]。大麦具有熟期早、丰产性好、生育期短、适应性广及抗逆性强等特点,作为粮食、饲料、酿造啤酒原料等广泛种植利用[3]。我国大麦还没有形成规模经济[4]。培育高产、优质、多抗性新品种是稳定和发展大麦产业的重要前提。遗传多样性是生物多样性的核心,也是作物种质资源和遗传育种研究的重要前提和基础。研究作物种质资源进行遗传多样性,对挖掘优良大麦品种和优异基因、改善品种遗传基础单一和防止基因流失具有重要意义[5]。【前人研究进展】分子标记技术评价大麦种质资源的研究已有报道[6-9],但育种实践中由于群体过大[10]、条件受限等原因未能广泛应用,而基于农艺性状的表型性状的多样性评价最为直观、经济、适合较大群体、应用更加简便[11]。其中作物的株高、单株穗数、穗长、穗粒数、千粒重等农艺性状是作物育种中重点性状[12, 13]。种质资源是大麦遗传育种和解析复杂性状的重要基础。【本研究切入点】目前,利用表型变异分析、相关分析、主成分分析和聚类分析对大麦农艺性状的评价研究较多,而对于新疆种植大麦种质资源多样性研究较少,且各研究所用的大麦种质资源材料来源不同,种植区域也不同,生态环境差异较大,表现出不同的遗传多样性[11, 14-16]。对大麦种质资源的表型性状多样性进行分析与评价,有助于丰富大麦育种亲本类型,加强优异基因的挖掘与利用。【拟解决的关键问题】分析新疆种植大麦种质资源的农艺性状表现,以157份大麦种质资源为材料,研究14个主要农艺表型性状,运用遗传多样性指数、主成分分析、相关分析及聚类分析评价其遗传多样性,为新疆大麦品种选育和品种改良提供参考。
1 材料与方法
1.1 材 料
以国家作物种质资源库(新疆分库)收集保存的157份国内外大麦品种(系)为参试材料。表1
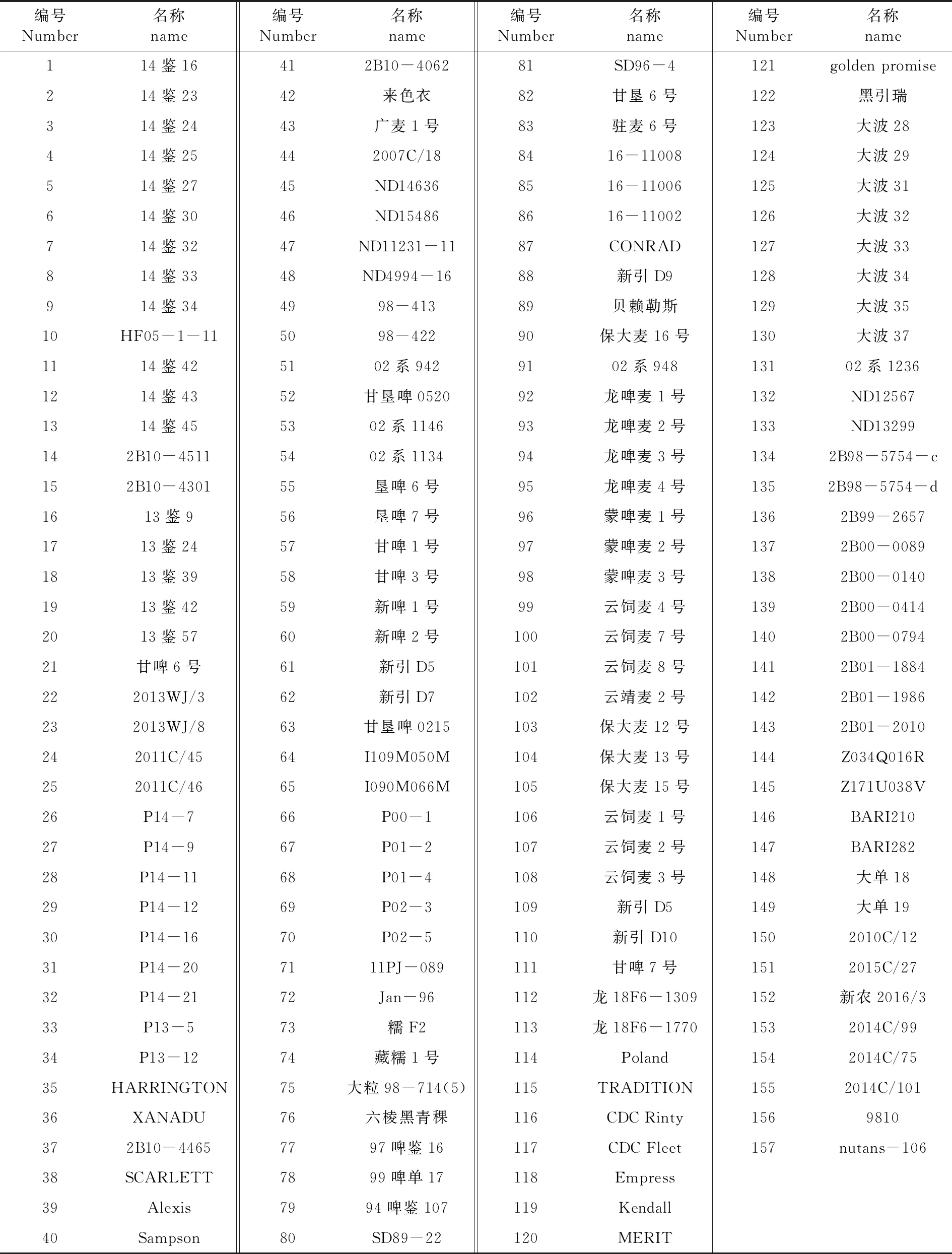
表1 供试大麦种质资源材料
2019~2020年将157份试验材料种植于新疆农业科院奇台麦类试验站(N44°13′,E89°12′),平均海拔823 m。采用随机区组设计,每个材料播种2行,行长2 m,行宽1.2 m,行距30 cm,株距3 cm,3次重复,于4月播种,采取常规田间管理。
1.2 方 法
以《大麦种质资源描述规范和数据标准》中的规定[17]为准,田间调查记载幼苗生长习性、株型、棱形、穗密度、穗姿、带壳性6个质量性状;成熟期随机选取长势一致、株型一致的10株植株室内考种,调查株高、单株穗数、穗长、每穗粒数、千粒重、粒长、粒宽、粒长/粒宽8个数量性状。
1.3 数据处理
使用 Microsoft Excel 365软件对数据进行初步整理,计算各性状的平均值、最大值、最小值、标准差和变异系数。遗传多样性指数(Shannon- Wiener diversity index,H′)的计算采用 Shannon-weaver,计算公式:H′=-∑PilnPi,式中,Pi为某一性状第i个级别出现的频率。对数量性状进行分级,对质量性状予以赋值,多样性指数的分级:计算参试材料总体平均数(X)和标准差(d),划分为10级,从第1级[Xi<(X-2d)] 到第10级 [Xi>(X+2d)],每0.5 d为1级,每一级的相对频率用于计算多样性指数[18]。
基于SPSS 21.0软件进行主成分分析、相关性分析及聚类分析,采用欧氏距离估算各材料间遗传差异,以离差平方和法进行聚类。
2 结果与分析
2.1 供试材料质量性状遗传多样性指数变化
研究表明,157份大麦种质资源的6个质量性状之间的遗传多样性指数变幅为0.18~0.65,存在广泛的遗传多样性,其中穗姿的遗传多样性指数最高(0.65),带壳性的遗传多样性指数最低(0.18),其余4个质量性状的遗传多样性指数从大到小依次为株型(0.64)、幼苗生长习性(0.45)、棱形(0.33)、穗密度(0.24)。种质的幼苗生长习性以直立型为主,所占比例为88%;株型以紧凑型为主,所占比例为67%;棱形以二棱大麦为主,所占比例达90%;穗密度以稀疏型为主,所占比例为94%;穗姿以直立型为主,所占比例为77%;带壳性以有皮壳为主,所占比例为96%。表2
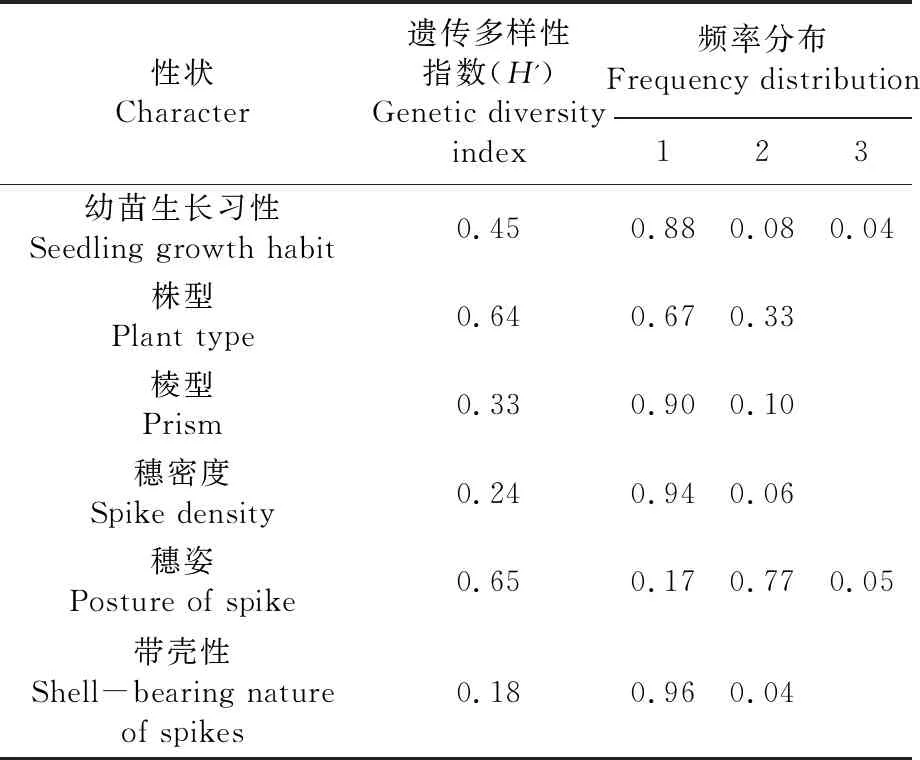
表2 8个质量性状的遗传多样性
2.2 供试材料数量性状遗传变异比较
研究表明,各性状的变异系数差异较大,除了粒宽和粒长/粒宽最小外,其余6个性状的极差较大,每穗粒数的变异系数最大(24.69%),粒宽的变异系数最小(4.07%),其余性状的变异系数从大到小依次为穗长(15.49%)、株高(10.50%)、单株穗数(8.13%)、千粒重(7.81%)、粒长/粒宽(6.57%)、粒长(5.42%),新疆大麦种质资源的每穗粒数、穗长在种质间差异较大,粒长和粒宽在种质间差异较小。
8个数量性状的遗传多样性指数相近,每穗粒数的最小(1.37),株高最大(2.04),其余6个性状的遗传多样性指数从大到小依次为单株穗数(2.03)、千粒重(1.98)、粒宽(1.95)、穗长(1.93)、粒长/粒宽(1.91)、粒长(1.90),供试材料在8个主要农艺性状上的遗传多样性水平普遍较高,遗传改良潜力较大。表3

表3 11个质量性状的遗传变异
2.3 供试材料数量性状相关性比较
研究表明,株高与穗长呈极显著正相关(相关系数为0.294),与粒长、粒长/粒宽呈显著负相关(相关系数分别为-0.195,-0.171),株高增加有利于穗长的增加,但会导致粒长和粒长/粒宽的减小;单株穗数与每穗粒数呈极显著负相关(相关系数为-0.293),与粒长/粒宽呈显著负相关(相关系数为-0.179),单株穗数增加会导致每穗粒数和粒长/粒宽的减小;穗长与粒宽呈显著正相关(相关系数为0.195),与每穗粒数、粒长、粒长/粒宽呈极显著负相关(相关系数分别为-0.270,-0.256,-0.375),穗长的增加有利于粒宽的增加,但会造成每穗粒数减少和粒长的减小,导致粒长/粒宽减小;每穗粒数与粒长/粒宽呈极显著正相关(相关系数为0.528),与粒长呈显著正相关(相关系数为0.166),与千粒重和粒宽呈极显著负相关(相关系数分别为-0.502,-0.532),每穗粒数的增加有利于粒长的增加和导致粒宽的减小,粒长的增加和粒宽的减小使粒长/粒宽增加,但会导致千粒重的减小;千粒重与粒长、粒宽呈极显著正相关(相关系数分别为 0.461,0.788);粒长与粒长/粒宽呈极显著正相关(相关系数为0.771),与粒宽呈显著正相关(相关系数为0.185);粒宽与粒长/粒宽呈极显著负相关(相关系数为-0.473)。大麦数量性状之间相互影响,彼此关联,存在此消彼长的关系。
8个数量性状中大部分性状间存在显著或极显著正相关或负相关。表4

表4 供试材料数量性状间的相关性
2.4 数量性状主成分特征值及贡献率变化
研究表明,选取前3个较大的特征值,代表8个数量性状73.351%的变异。入选的3个特征值、累计贡献率及8个质量性状的特征向量。第1主成分的特征值为2.730,贡献率为34.127%,是最主要的成分,穗长(0.508)、千粒重(0.556)、粒宽(0.760)具有较高的正载荷,此类因子均为大麦的产量因子;这3个性状的增加会增加大麦籽粒重量,但会导致每穗粒数(-0.818)、粒长/粒宽(-0.806)指数的下降,影响大麦产量。第2主成分的特征值为2.084,贡献率为26.053%,千粒重(0.774)、粒长(0.870)、粒宽(0.518)具有较高的正载荷,该主成分反映的主要是千粒重因子。第3主成分的特征值为1.054,贡献率为13.171%,株高(0.645)、穗长(0.479)具有较高的正载荷,此类因子反映的主要是株高-穗长因子;2个性状的增加会增加大麦的生物产量,但会导致单株穗数(-0.560)的减小,不利于大麦的产量的增加。表5
2.5 数量性状聚类平均值差异比较
研究表明,在欧氏距离系数为5时,可将157份大麦品种分为4大类群。
第Ⅰ类群包含72份材料,占供试材料的45.9%,这一类群主要表现为每穗粒数少、千粒重较高、粒宽宽,其余5个性状适中,多为高千粒重型资源。
第Ⅱ类群包含64份材料,占供试材料的40.8%,这一类群主要表现为矮秆、千粒重高、长粒,单株穗数较多、穗长较长、每穗粒数较多、粒宽较宽、粒长/粒宽较长,多为矮秆大粒型资源。
第Ⅲ类群仅包含12份材料,占供试材料的7.6%,这一类群主要表现为高秆、单株穗数多、穗长长、粒长短、粒长/粒宽短,每穗粒数、千粒重和粒宽适中,多为高秆多穗长穗型资源。
第Ⅳ类群仅包含9份材料,占供试材料的5.7%,这一类群所含材料最少,均为六棱大麦,主要表现为单株穗数少、穗长短、每穗粒数多、千粒重低、粒宽低、粒长/粒宽高,株高和粒长适中,多为少穗多粒型资源。图1,表6

表5 供试材料数量性状的主成分比较

表6 供试大麦3种类群材料8个数量性状的平均值差异比较

图1 供试材料基于8个数量性状聚类
3 讨 论
大麦的品种选育方式以种间杂交选育为主,收集和筛选丰富的大麦种质资源是提高大麦产量、品质和抗性的必要前提[19]。农艺性状是作物评价的直观依据,与作物的产量和品质相关,对其遗传多样性的分析是丰富作物种质资源类型,指导作物亲本选配,进行作物育种的基础[20]。徐肖等[21]对86份青藏裸大麦的主要农艺性状的表型多样性进行分析,发现8个质量性状和4个数量性状均存在着丰富的多样性,并且数量性状遗传多样性高于质量性状。夏腾飞等[14]对267份青稞种质的9个数量性状进行研究表明,各性状的遗传多样性指数相近,平均值为2.03,存在丰富的表型多样性。研究对大麦的6个质量性状进行分析发现,存在较为广泛的遗传多样性;8个数量性状的遗传多样性指数较接近,平均值为1.89,存在着丰富的多样性,并且数量性状的遗传多样性高于质量性状。研究结果同上述结果相一致,供试大麦资源具有较高的丰富度和均匀度,遗传多样性广泛,并且数量性状的变异较质量性状更为丰富,这可能与研究材料来源于不同的生源地,且不同地区生态环境差异大有关。
研究结果显示,各性状之间相互影响,彼此关联,提高某一性状可能会促进一些性状的提高,但会导致其他性状的降低,与前人研究结果相似[15]。如株高与穗长,穗长与粒宽,每穗粒数与粒长/粒宽、粒长,均呈显著或极显著正相关;而株高与粒长、粒长/粒宽,穗长与每穗粒数、粒长、粒长/粒宽,每穗粒数与千粒重、粒宽,均呈显著或极显著负相关。
主成分分析法已被广泛的用于小麦[22]、大麦[23]、水稻[24]等农作物的种质资源评价中,能够直观地区分不同种质的农艺、产量性状的优劣及评价特异种质[25]。
赵斌等[11]将111份大麦种质的7个农艺性状简化为3个主成分,累计贡献率80%,其中第1主成分反映的主要是产量因子,第2和第3主成分反映了株高、千粒质量,主成分内各性状之间之间表现为相互促进,相互抑制的关系。聚类分析已被广泛用于评价种质资源的差异性和分类[26, 27]。研究对供试157份大麦种质资源进行了主成分分析,归纳为3个主成分,累计贡献率达73.351%,包含了绝大多数农艺性状的信息。其中第1主成分为产量构成因子穗长、千粒重、粒宽;第2主成分为反映了千粒重因子;第3主成分为株高-穗长因子。第1主成分中产量因子间存在相互促进的关系,而产量因子和每穗粒数、粒长/粒宽性状间存在相互抑制的作用;第3主成分中株高和穗长性状间存在相互促进关系,而这两个性状和每穗粒数存在相互抑制的关系,产量相关因子间存在矛盾,与前人研究结果相一致。
4 结 论
将供试大麦材料划分为4个类群,第Ⅰ类群表现为每穗粒数少、千粒重较高、粒宽宽型资源;第Ⅱ类群属于矮秆大粒型资源;第Ⅲ类群属于高秆多穗长穗型资源;第Ⅳ类群属于少穗多粒型资源;各个性状表现各有优势。第Ⅰ类群、第Ⅱ类群所含的种质数多,第Ⅲ类群和第Ⅳ类群所含种质数非常少,四个类群中均含有新疆大麦材料,前3类群中新疆大麦材料分别占各类材料的42.5%、38.55%、38.5%,占比较多,参试大麦种质特性差异较大,可作为不同类型的优异育种亲本。
参考文献(References)
[1] 张国良, 戴其根, 张洪程, 等. 江苏省大麦生产现状分析及其发展对策[J]. 大麦科学, 2004,(4):6-9.
ZHANG Guolian, DAI Qigen, ZHANG Hongcheng, et al. Analysis of Barley Production Status and Development Strategies in Jiangsu Province[J].BarleyScience, 2004,(4): 6-9.
[2]徐先良, 赖勇, 王鹏喜, 等. 大麦亲本材料农艺性状鉴定及遗传多样性分析[J]. 麦类作物学报, 2013, 33(4):640-646.
XU Xianliang, LAI Yong, WANG Pengxi, et al. Identification of Agronomic Traits and Genetic Diversity Analysis of Barley Parent Materials [J].JournalofTriticeaeCrops, 2013, 33(4):640-646.
[3]刘亚楠. 大麦种质资源遗传多样性研究及种质的评价与筛选[D]. 扬州:扬州大学, 2017.
LIU Yanan.StudyonGeneticDiversity,EvaluationandSelectionofGermplasminBarley[D]. Yangzhou: Yangzhou University, 2017.
[4]曹灵灵, 王溶花. 我国谷物类粮食进口贸易现状及问题分析[J]. 粮食科技与经济, 2020,45(3):19-21.
CAO Lingling, WANG Ronghua. Analysis on the current situation and problems of grain import trade in China [J].GrainTechnologyandEconomy, 2020, 45(3):19-21.
[5]孟霞, 卓嘎, 大次卓嘎, 等. 西藏部分青稞主要农艺性状分析[J]. 麦类作物学报, 2010,30(6):1043-1047.
MENG Xia, Zhuoga, Dacizhuoga, et al. Analysis of Main Agronomic Traits of Several Tibetan Naked Barley [J].JournalofTriticeaeCrops, 2010, 30(6): 1043-1047.
[6]尚毅, 华为, 朱靖环, 等. 浙江省裸大麦地方品种遗传多样性分析[J]. 麦类作物学报, 2014, 34(7):922-928.
SHANG Yi, HUA Wei, ZHU Jinghuan, et al. Genetic diversity analysis of naked barley landraces in Zhejiang Province[J].JournalofTriticeaeCrops, 2014, 34(7):922-928.
[7]张宇, 司二静, 孟亚雄, 等. 不同来源大麦材料的农艺性状和籽粒蛋白质含量及群体遗传结构分析[J]. 麦类作物学报, 2015, 35(7):940-947.
ZHANG Yu, SI Erjin, MENG Yaxiong, et al. Analysis on Agronomic Traits, Grain Protein Content and Population Structure of Barley Germplasm from Different Geographical Origins [J].JournalofTriticeaeCrops, 2015, 35(7): 940-947.
[8]张超, 才让卓玛, 卡毛先, 等. 应用ISSR分子标记分析裸大麦的遗传多样性[J]. 分子植物育种, 2020, 18(4):1193-1201.
ZHANG Chao, Cairang Zhuoma, KA Maoxian, et al. Genetic Diversity of Naked Barley Analyzed by ISSR Molecular Markers [J].MolecularPlantBreeding, 2020, 18(4):1193-1201.
[9]余其, 王振波, 沈真辉, 等. 分子标记技术及其在大麦遗传育种中的应用[J]. 大麦与谷类科学, 2016, 33(2):1-5.
YU Qi, WANG Zhenbo, SHEN Zhenhui, et al. Molecular Marker Technologies and their Applications to Barley Genetics and Breeding [J].BarleyandCerealScience, 2016, 33(2):1-5.
[10]蔡羽, 杨平, 冯宗云. 大麦表型多样性分析及优异饲草种质资源筛选[J]. 植物遗传资源学报, 2019, 20(4):920-931.
CAI Yu, YANG Ping, FENG Zongyun. Characterization of Phenotypic Variation in Cultivated Barley Provided Elite Genetic Germplasm with Potential Breeding for Silage Barley [J].JournalofPlantGeneticResources, 2019, 20(4): 920-931.
[11]赵斌, 陈晓东, 季昌好, 等. 111份多棱大麦种质主要农艺性状的遗传多样性[J]. 大麦与谷类科学, 2020, 37(5):1-7.
ZHAO Bin, CHEN Xiaodong, JI Chanhao, et al. Analysis of the Genetic Diversity of the Main Agronomic Traits of 111 Multi-rowed Barley Germplasm Accessions [J].BarleyandCerealScience, 2020, 37(5):1-7.
[12]刘福. 引进大麦种质资源的评价与育种利用[D]. 杨凌:西北农林科技大学,2015.
LIU Fu.EvaluationandBreedingUtilizationoftheIntroducedBarleyGermplasms[D]. Yangling: Northwest A & F University, 2015.
[13]刘亚楠, 潘雨涵, 郭晖, 等. 六棱大麦种质表型遗传多样性评价[J]. 植物遗传资源学报, 2018, 19(5):846-856.
LIU Yanan, PAN Yuhong, GUO Hui, et al. Comprehensive Evaluation of Genetic Diversity in Six-rowed Barley Germplasm [J].JournalofPlantGeneticResources, 2018, 19(5): 846-856.
[14]夏腾飞, 王蕾, 徐金青, 等. 267份青藏高原青稞种质材料的表型多样性分析[J]. 西北农业学报, 2018, 27(2):182-193.
XIA Tengfei, WANG Lei, XU Jinqing, et al. The Genotypic Diversity Analysis of 267 Six-rowed Hulless Barley Accessions from the Qinghai-Tibetan Plateau [J].ActaAgriculturaeBoreali-occidentalisSinica, 2018, 27(2): 182-193.
[15]蒋莹, 常蕾, 王安, 等. 143份大麦种质资源主要农艺性状遗传多样性分析[J]. 江苏农业科学, 2020, 48(14):94-98.
JIANG Ying, CHANG Lei, WANG An, et al. Genetic diversity of main agronomic characters of 143 barley germplasm resources [J].JiangsuAgriculturalSciences, 2020, 48(14):94-98.
[16]李守明, 梁维, 魏凌基, 等. 新疆大麦种质资源农艺性状和醇溶蛋白遗传多样性分析[J]. 新疆农业科学, 2009, 46(2):269-274.
LI Shouming, LIANG Wei, WEI Lingji, et al. Agronomic Trait of Germplasm Resources and Genetic Diversity of Hordein in Xinjiang Barley [J].XinjiangAgriculturalSciences, 2009, 46(2): 269-274.
[17]张京,刘旭.大麦种质资源描述规范和数据标准[M].北京: 中国农业出版社,2006.
ZHANG Jing, LIU Xu.DescriptorsandDataStandardforBarley(HordeumvulgareL.) [M].Beijing: China Agriculture Press, 2006.
[18]Shannon C E, Weaver W. The mathematical theory of communication [J].TheUniversityofInois,Urbana,Chicago,USA, 1949:3-24.
[19]刘亚楠. 大麦种质资源遗传多样性研究及种质的评价与筛选[D]. 扬州:扬州大学, 2017.
LIU Yanan.StudyonGeneticDiversity,EvaluationandSelectionofGermplasminBarley[D]. Yangzhou: Yangzhou University, 2017.
[20]Campbell R D. Using phenotypic manipulations to study multivariate selection of floral trait associations [J].AnnalsofBotany, 2009, 103(9): 1557-1566.
[21]徐肖, 栾海业, 张英虎, 等. 青藏高原裸大麦种质资源形态多样性分析[J]. 浙江农业学报, 2019, 31(7):1037-1044.
XU Xiao, LUAN Haiye, ZHANG Yinghu, et al. Morphological Diversity of Hulless Barley Accessions from Qinghai-Tibetan Plateau [J].ZhejiangAgriculturalJournal, 2019, 31(7):1037-1044.
[22]李晶, 南铭. 俄罗斯和乌克兰引进冬小麦在我国西北地区的农艺性状表现和遗传多样性分析[J]. 作物杂志, 2019,(5):9-14.
LI Jin, NAN Ming. Analysis of Agronomic Characters and Genetic Diversity of 62 Winter Wheat Germplasms from Rgussia and Ukraine in Northwest China [J].JournalofCrops, 2019,(5): 9-14.
[23]王蕾, 徐金青, 夏腾飞, 等. 青藏高原青稞及其他地区大麦种子表型的多样性分析[J]. 西北农业学报, 2014, 23(3):40-44.
WANG Lei, XU Jinqing, XIA Tenfei, et al. Phenotypic Diversity Analysis of Seed Traits in Barley Germplasm from Qinghai-Tibetan Plateau and Other Regions [J].ActaAgriculturaeBoreali-occidentalisSinica, 2014, 23(3): 40-44.
[24]陈峰, 朱其松, 徐建第, 等. 山东地方水稻品种的农艺性状与品质性状的多样性分析[J]. 植物遗传资源学报, 2012, 13(3):393-397, 405.
CHEN Fen, ZHU Qisong, XU Jiandi, et al. Diversity Analysis of Agronomic and Quality Characters of Rice Landraces in Shandong[J].JournalofPlantGeneticResources, 2012, 13(3): 393-397, 405.
[25]要燕杰, 高翔, 吴丹, 等. 小麦农艺性状与品质特性的多元分析与评价[J]. 植物遗传资源学报, 2014, 15(1):38-47.
YAO Yanjie, GAO Xiang, WU Dan, et al. Multivariate Analysis and Evaluation of Agronomic and Quality Traits Based on Principal Components in Wheat[J].JournalofPlantGeneticResources, 2014, 15(1): 38-47.
[26]张鹏, 张海洋, 郑永战, 等. 芝麻种质资源因子分析及聚类分析[J]. 中国油料作物学报, 2008,(1):71-78.
ZHANG Peng, ZHANG Haiyang, ZHNEG Yongzhan, et al. Factor analysis and cluster analysis of sesame germplasm resources [J].ChineseJournalofoilCropSciences, 2008,(1): 71-78.
[27]张彩英, 常文锁, 谢令琴, 等. 小麦高代新品系鉴定的聚类分析[J]. 植物遗传资源科学, 2001,(4):29-33.
ZHANG Caiying, CHANG Wensuo, XIE Lingqin, et al. Cluster analysis on identification of new wheat lines of advanced generation [J].PlantGeneticResourcesScience, 2001,(4): 29-33.
