大围山蕨类多样性及生境因子对干扰的响应
2023-01-09杨逢春刘景欣黄华平构箭勇文慧婷袁欣宇马秀花朱兴旺阳晓倩PrueksakornKritanaSarathchandraChaya
杨逢春,刘景欣,黄华平,构箭勇,文慧婷,李 叶,袁欣宇,马秀花,朱兴旺,阳晓倩,何 琼*,Prueksakorn Kritana,Sarathchandra Chaya
(1.宜宾职业技术学院现代农业学院,四川 宜宾 644003;2.中国科学院西双版纳热带植物园环境教育中心,云南 勐腊 666302;3.中国热带农业科学院环境与植物保护研究所,海南 海口 570100;4.云南省红河哈尼族彝族自治州气象局,云南 蒙自 661100;5.玛希顿大学环境与资源研究系,泰国 佛统府 73170;6.中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085;7.斯里兰卡拉贾拉塔大学应用科学学院生物科学系,斯里兰卡密亨塔勒 50300)
近年来,随着人类对森林的不断开发,原始植被遭到了严重破坏。这些地方的生态环境和主要环境因子(如水分、温度等)都会受到一定的影响。随着各国对生态的关注,关于人类干扰对不同生态系统植被群落影响的相关研究也越来越多。崔玉华等[1]研究表明,在小范围林区,未干扰林分内的物种间关系表现为无关联,中度干扰表现为正相关,严重干扰表现为负相关,而适当的干扰有利于林分内物种间对环境资源的协同利用。郑绍伟等[2]通过研究木本植物多样性对人为干扰的响应来评估原始林受破坏程度和植物群落稳定性减弱程度,结果表明,干扰程度对乔木占比影响不显著,而与灌木占比呈正相关。此外,森林的乔木密度、高度、高盖度和胸径等受人为干扰后存在差异[3]。对于草原生态系统,放牧是导致物种多样性丧失的主要原因,样地密度、形状复杂度和边缘密度对系统发育多样性也有负面影响[4]。
大围山位于云南省东南部,属亚热带海洋性气候区,年均温16~22.6℃,年均降水量1 777.7 mm,空气相对湿度达85%,是我国稀有的湿润雨林[5]。近年来该地区遭受道路修建、旅游开发、农业种植、采茶采药等不同程度的人为干扰。在这些干扰下,该地区主要环境因子如何变化,植物与植被如何响应有待进一步研究。针对这些问题,本研究以对环境因子较敏感的蕨类植物作研究媒介,研究了以下内容:(1)人为干扰对大围山林下蕨类植物多样性的影响。(2)人为干扰对大围山水分、温度等主要环境因子的影响。(3)人为干扰下主要环境因子变化与蕨类植物多样性的关系。
1 研究方法
1.1 干扰程度的定义以及研究位置的选择
根据人为干扰的程度不同选择了三个研究位置,分别为样带1(海拔1490 m)、样带2(海拔1 533 m)、样带3(海拔1 630 m),此三个研究位置海拔相近(1 490~1 630 m)、气候环境相似(表1)。每个样带取10 个样方,每个样方的面积为1 m2。参照郝建锋等[6]对人为干扰强度的划分方法,结合研究区域实际情况作适当调整,划分为以下3个等级:
轻度干扰:没有或少有游客到达,没有或少有人类活动的痕迹。
中度干扰:有部分游客到达,草本层植被遭受轻度践踏,有部分人为折枝痕迹。
重度干扰:人类活动频繁,选择性采伐明显,有较多垃圾。
1.2 数据处理
1.2.1 样方设计与环境因子测量
每个调查位置选取长100 m 样带为调查对象,每隔10 m 设置一个1 m×1 m 样方,共10个,统计各样方内的蕨类植物种类及数量,记录、采集标本并拍照。并检测林冠开阔度(Canopy openness)、空气相对湿度(Relative humidity)和表层土温(Surface temperature)等三个主要环境因子,测定每个样方中心点的上述数据。林冠开阔度的获取使用180o鱼眼镜头(Canon,EF 8~15mm f/4L USM),早晨8 点之前在样方中心点水平向天空拍照,然后采用Image J软件计算空白区域。
1.2.2 数据统计分析
Alpha多样性指数,包括H:香农—威纳指数(均值);Hmax:香农—威纳指数(最大值);J:物种分布均一度指数;Dominant_index:显著性指数。
物种丰富度和稀有性指数,包括Species_estimated:在t个样方内的物种数量(累计值);Singletons:在t个样方内只有一个植株的物种数量(累计值),可理解为单个体种;Uniques:仅出现在一个样方中的物种数量(累计值),可理解为稀有种或孤立种;ACE:基于物种丰度的覆盖度指数(均值);ICE:基于物种发生率的覆盖度指数(均值);Chao2:物种丰富度指数。
将数据分为两组不同的数据矩阵。第一组数据矩阵为基于样方的物种个体数量,用于计算与物种丰度相关的值。第二组数据矩阵则只计算物种的出现次数,不计算个体数,用于计算与物种在样方内出现频率相关的值。所有数据矩阵全部加载到EstimateS(version 9.1)进行999 次计算。然后将计算结果输入R 程序,使用lme4程序包生成简单线性模型GLMM[7]。每条样带的样方内物种数量的期待值Species_estimated是基于个体在样方内的分布进行计算,使用Vegan 程序包进行多变量的分析。最后,使用Indicspecies程序包开展基于物种个体数和出现频率的指示种分析[8],多级模式分析(Multilevel pattern analysis)显著性水平控制在P<0.05。所有的模型均由卡方检验,显著性水平P<0.05。模型的残差由Shapiro—Wilk test[9]进行正态性评估(P≥0.5)。
2 结果和分析
调查结果表明,在三条样带中共发现蕨类植物14 科22 属29 种,476 株。其中,攀缘卷柏(Selaginella helferiWarb.)植株数量多,占总株数的38.03%;美丽复叶耳蕨(Arachniodes speciosa(D.Don)Ching)在12 个样方中均出现,出现频率最高,占总样方数的40%。在三条样带中,样带1 的蕨类植物最为丰富,共21 种,占总物种数的72.41%,蕨类个体287 株,占总株数的60.29%。
2.1 干扰强度对主要环境因子的影响
与样带2和样带3相比,样带1在受到重度干扰后,林冠开阔度(CO)增加,阳光辐射量明显增加,空气相对湿度(RH)明显降低,表层土温(ST)明显增加。
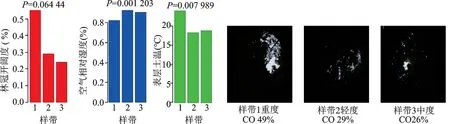
图1 不同干扰程度下的林冠开阔度、空气相对湿度和表层土温Fig.1 CO,RH,and ST in different levels of disturbances
2.2 不同干扰强度对样带间多样性变化的影响
不同干扰程度下,蕨类植物多样性的变化产生差异。多样性指数H和最大多样性指数Hmax在样带间的差异达到显著水平(P<0.05),说明随着三条样带之间受干扰程度不同,环境差异明显,进而影响了蕨类植物多样性指数(图2)。

图2 不同干扰程度下样带中香农—威纳指数H、香农—威纳指数(最大值)Hmax、均一度指数J和显著性指数Dominant_indexFig.2 Diversity index H,Diversity index the maximum Hmax,evenness index J,and Dominant_index in transects
受轻度干扰的样带2位于大围山自然保护区外缘,天然植被完整,林冠茂密,但多样性指数仅为1.83,样带内最常见物种为华中瘤足蕨(Plagiogyria euphlebia(Kunze)Mettenius),植株个体数最多的为攀缘卷柏,共45 株。受中度干扰的样带3 为人工种植的杉木林(Cunninghamia lanceolata(Lambert)Hooker),林冠茂密,林下间种少量茶(Camellia sinensis(Linnaeus)Kuntze),林下蕨类植物多样性指数为2.33,样带内最常见物种为蛇足石杉(Huperzia serrata(Thunb.ex Murray)Trev.),其在该样带内植株个体数量最多,共23 株。受重度干扰的样带1 位于水库边游客步道边坡,不定期的人为修整,林下蕨类植物多样性指数1.99,样带内最常见物种为姬蕨(Hypolepis punctata(Thunb.)Mett.);攀援卷柏的植株个体数最多,共136株。
优势度指数Dominant_index与均一度指数J均未呈现出显著性差异。优势种的计算与该物种植株的数量在整个样带中所占比例呈正相关,物种能适应多个环境因子复合作用,在对环境因子的耐受区间比较大的情况下,则该物种的优势度指数偏大。占据样方数量较多的种,不一定就是该样带的优势种。因此在受到不同干扰的三个样带中,蕨类物种单优群落出现的概率相同。攀缘卷柏为样带1和样带2的优势种,蛇足石杉为样带3的优势种。
2.3 不同干扰强度对样带内多样性变化的影响
不管在何种干扰强度下,三个样带中林下蕨类植物的多样性与样带长度和样方数量均呈显著正相关(P<0.05)。说明沿着调查路线的延伸,蕨类植物的多样性都显著增加。尽管某些地方受人为干扰(踩踏、植被修剪、边坡垮塌等),但因靠近水源地,植被覆盖良好,水分和热量能满足林下蕨类植物的生长繁殖需求,故多样性指数依然保持了较好的增长潜力(图3)。
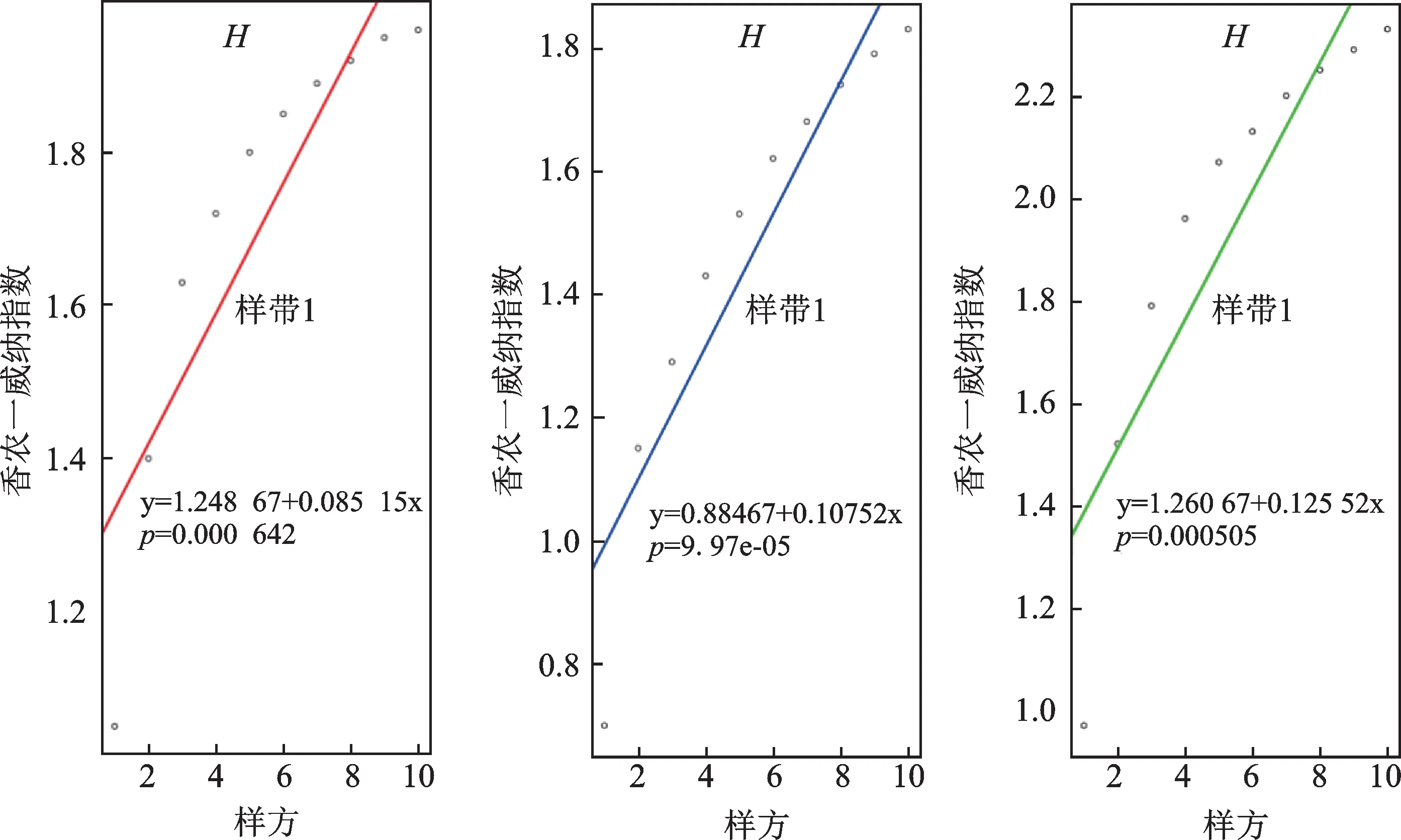
图3 不同样带内蕨类植物多样性变化趋势Fig.3 The trends of diversity indices of ferns in different transects
2.4 不同干扰强度对物种丰富度和稀有性指数的影响
2.4.1 物种期待值/累积曲线Species_estimated
该曲线表明物种数量随调查路线的延长而增加,样带1 高于其他样带。说明重度干扰后物种增加,且增加的趋势最快,这与其他研究者[10,11]的结果类似。证明环境在重度干扰下发生了变化,林下蕨类物种获得了更多光照和CO2供给,土壤温升高。光照强度和时长的提高不仅能促进蕨类植物的生理活动,增加养分合成,且能激活土壤中休眠蕨类孢子。食用双盖蕨(Diplazium esculentum(Retz.)Sm.)、东北蹄盖蕨(Athyrium brevifronsNakai ex Kitagawa)等孢子的休眠期可长达1~5年,遇合适的光照和温度条件才可萌发[12]。
2.4.2 单个体种/孤独种Singletons
受干扰最严重的样带1,其Singletons指数明显高于其他样带,而受干扰程度最轻的样带2,指数值最低。样带1 中出现孤独种为大叶黑桫椤(Alsophila giganteanWall.ex Hook.),紫萁(Osmunda japonicaThunb.),里白(Hicriopteris glauca(Thunb.)Ching),多羽凤尾蕨(Pteris decrescensChrist),华中瘤足蕨;样带2 中有姬蕨(Hypolepis punctate(Thunb.)Mett.),滇南桫椤(Alsophila austroyunnanensisS.G.Lu),四回毛枝蕨(Leptorumohra quadripinnata(Hayata)H.Ito);样带3 中有里白(Hicriopteris glauca(Thunb.)Ching)。所有孤独种均为土生大株型,根状茎直立或斜升,能支撑较大羽状叶。要维持较大的植株个体就得占用更多的水分、光照和空间资源,而一些体型较小种类如假鞭叶铁线蕨(Adiantum malesianumGhatak)和星毛蕨(Ampelopteris prolifera(Retz.)Copel)对资源的需求较少,而且也没有足够大的植株去竞争上层生存空间,因此它们采用另一种策略,发展数量,占据了底层空间。这种现象在环境资源充沛、生态位丰富的热带森林里是一种正常现象。
2.4.3 单样方种/稀有种指数Uniques
这个指数主要是通过区分对生境需求差异较大的物种,借以指征同一个样带内存在的多种不同生境,也可以用来识别极小种群。Uniques曲线与Singletons曲线走势基本一致,样带1 >样带3 >样带2。样带1 和样带3 在第四个样方之后,稀有种指数没有继续增加,样带3甚至还呈现下降的趋势,样带2虽然指数较低,但是保持持续增长的态势,随着调查路线的延长,还将有更多的稀有种被发现。说明受到干扰程度最低的样带2,其生境异质性保持良好,随着样带的延伸,生长基质、林冠变化、水分条件、温度条件、种间竞争等环境因子将会出现更多的组合变化,蕨类植物的生长繁殖环境也越丰富。样带1 和样带3 在受到短期的人为干扰之后,出现了较多的差异化生境,为林下蕨类植物创造了生长的机会,但是可持续性较差,随着调查路线的延长,差异化生境不再增加,Uniques稀有指数亦不再增加。
2.4.4 物种丰富度指数ACE、ICE和Chao2
三者都用于估计样带/样方内的物种丰富度,只不过ACE是基于物种个体丰度而计算,ICE是基于物种的发生率来计算,而Chao2则是通过Singletons的结果经过二次修正而得,尽管是一个理论丰富度,但是更接近真实的值[13-14]。计算结果如图4显示,ACE和Chao2均呈逐渐上升的趋势,而ICE指数在前面1~3 个样方呈上升,而后面逐渐下降并趋于平缓,其值稳定在21.02 到27.59 之间。但是整体上,受到重度人为干扰的样带1 的物种丰富度明显高于其它两条样带。在受到严重干扰的情况下,样带1 和2 在受到短期的人为干扰之后,出现了较多的差异化生境,为林下蕨类植物创造了生长的机会,这一结果符合Connell的干扰假说[15]。但需要注意的是,在重度干扰情况下,由于环境可持续性较差,后期的生境将趋向于单质化,样方中新物种的发生率将逐渐降低,这与图形显示的结果是一致的。
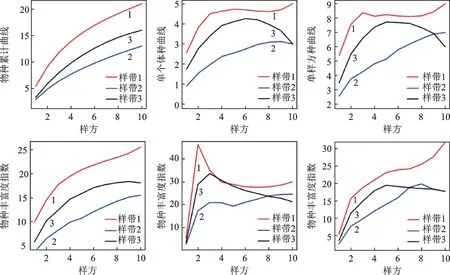
图4 样带及样方内物种数量及丰富度变化Fig.4 The variance of species number and abundance in transects and plots
2.5 不同干扰程度下,指示种的差异
指示种与特定的生活环境相关,它的出现与缺失、种群密度变化、传布和繁殖成功率变化能反映其生活环境的改变。经多级模式分析(Multilevel pattern analysis)一共筛选得6个指示种,其在不同干扰程度下的分布及指示性有差异(表1)。在受干扰程度最轻的样带2,能够用以指征环境的林下蕨类物种最多,指示的生境也最多。受到严重干扰的样带1有4个指示种,而受到中度干扰的样带3,出现3个指示种。蛇足石杉在样带3 中大量出现,10 个受调查的样方中有7 个样方发现该种;相对于样带2,仅在1个样方中有发现;样带1则无发现。

表1 指示种在各样带内的分布及显著性水平Tab.1 Distribution and significance of indicators in different transects
3 结论和讨论
3.1 不同干扰程度下,林下蕨类植物群落物种组成的变化
蕨类植物不同物种之间对生态因子耐受性的差异(生态幅)使得它们在不同干扰强度下产生的生态环境变化作出差异化响应,这是本研究的理论基础。由于立地特征是影响物种组成的最重要的因素[16],而干扰可以增加立地条件生境异质性,所以,干扰对林下蕨类植物群落更新或重建过程中物种组成的变化必然会起到重要的作用。
重度干扰往往是以对植物物种的“去除”为结果,但其作用过程中物种组成上的变化却不同[17-19]。如样带1中不定期对行人步道两侧过度生长的植株进行修剪和铲除,一些恢复较快的物种如卷柏属Selaginella在修剪过后获得足够光照快速生长,并在早期的竞争中获得优势,而恢复较慢的物种如下延叉蕨(Tectaria decurrens(Presl)Cop.)则在干扰后的演替中处于劣势或彻底消失。受中度干扰的样带3属早期人为干预下达到的顶级群落,物种构成稳定,林间层次明显[19],其干扰主要来源于采茶、采药及游客野外大小便。在此情况下,除对蛇足石杉的个体数量造成变化外,蕨类群落物种组成基本稳定,无类似样带1 的演替现象发生。样带2 属天然林,已达到自然演替顶级,物种构成稳定、层次丰富,除偶尔抵达的护林员和野生动物造成轻度践踏外,未受到其它干扰,因此其林下蕨类植物群落稳定。
3.2 不同干扰程度下,林冠开阔度与蕨类植物生长繁殖的关系
相较于样带1,样带2 和样带3 的林冠开阔度要低得多,且因林冠完整性未受到破坏,因此各个样方之间的开阔度无较大的波动,林下空间的生态因子未受到影响,生态系统比较稳定。样带1 在受到重度干扰的情况下,林冠开阔度急剧增加,与之关联的生态因子如日照长度、辐射强度等明显增加,空气流动性加大、相对湿度显著降低,地表漫反射增多、土壤增温明显。对蕨类植物而言,光照和地温的变化对打破蕨类孢子的休眠,促进孢子萌发,促进配子体和孢子体形态建成具有重要的意义[20-22]。林冠打开后,喜光喜温型的蕨类物种得以快速发展。因此我们在样带1发现的林下蕨类物种数量与植株个体数量要明显多于其它两条样带。
3.3 不同干扰程度下,水分条件与蕨类植物种丰富度的关系
附生型蕨类已被证明对空气中水分以及环境的郁闭度有着强烈的依赖性[23-24],而土生蕨类植物是否也有着相同的依赖性呢?杨逢春等[25]在元江干热河谷的调查结果表明,在水分极度缺乏的情况下,蕨类物种数量极少且以旱生型为主,如毛叶粉背蕨(Aleuritopteris squamosa(C.Hope & C.H.Wright)Ching)、旱生卷柏(Selaginella stauntonianaSpring)以及蜈蚣草(Pteris vittataLinnaeus)。早先已发表的研究亦证明空气中的水分是某些蕨类植株形态建成、生物量积累和配子体正常发育的关键条件[12,26-28]。但是,在大围山云雾林环境下,由于三条样带皆靠近水库,水源稳定、湿度大[5]。尽管重度干扰情况下,空气相对湿度有所下降,但是根部水源供给没有变化。不同样带间水分条件差别不大,对不同干扰程度下蕨类植物丰富度的影响较小。
3.4 不同干扰程度下,生境多样化与蕨类植物稀有指数和物种丰富度的关系
根据生境异质性假说,不同的物种需要不同的生境,生境复杂的区域往往拥有更为丰富、多样的生物类群;反之,在生境相对较为单一的区域,其物种的丰富度也相对较低[29-30]。张伶俐等[31]对云南地区蕨类植物多样性分布格局的研究表明,生境异质性对多样性格局的影响力有重要作用。在生境越复杂的地方,发现的蕨类植物的种类也越多。在受到重度或中度人为干扰之后,为林下蕨类植物创造了生长的机会,它们的物种数量和植株数量均有所增加,但是由于可持续性差,随着样带的延伸,生境趋同性明显,其Uniques曲线逐渐趋于平滑。而在轻度干扰下,原始生境中差异化生境依然存在,以致Uniques曲线持续增长,物种丰富度也同时保持持续增长的态势,这也是热带云雾林演替到顶级状态,对空间资源利用高度细化的体现。
3.5 不同干扰程度下,指示种的差异
绝大多数蕨类植物对外界自然环境的要求十分严格,对生态环境具有高度的敏感性,不同的物种对环境具有严格的选择性,只有满足条件才能够生存下去[28,32-33]。本研究中,在轻度干扰环境筛选出的指示种最多,重度干扰次之,中度干扰最少。因为轻度干扰下,保留的原始生境最多,重度干扰虽然破坏了原有的生境,但是由于改变了光照、温度等主要生态环境因子,多种生态因子重新组合之后,又形成了新的生境,因此给其它蕨类物种的出现创造了机会。而中度干扰的杉木林,物种结构比较单一、群落层次少、生境一致性较高,因此环境指示种较少,这与Bellone[29]和Gao[30]等的研究结果是一致的。需要注意的是,本研究中指示种是程序计算的结果,需要结合实际考虑,如姬蕨和华南毛蕨是亚洲广布种,此处作为指示种没有意义。蛇足石杉对杉木林生境有偏好,有强烈的指示性。

附表 所有样带内蕨类植物名录(采用秦仁昌系统)Appendix Ferns list in all transects(Following system of Ren-Chang Qin)
