橡胶树耐低温评价及爆皮流胶差异性比较
2023-01-07程琳琳谢贵水王纪坤彭文涛周立军
程琳琳,安 锋,谢贵水,王纪坤,彭文涛,周立军
(1. 中国热带农业科学院橡胶研究所, 海口 571101;2. 海南儋州热带农业生态系统国家野外科学观测研究站, 海南 儋州 571737)
【研究意义】巴西橡胶树[Heveabrasiliensis(Willd. ex A. Juss.) Muell. Arg.],以下简称橡胶树,原产地的生长环境造就了其对高气温的要求。虽然,我国在橡胶树北缘栽培中取得了卓越的成就,但是由于冬春季受寒潮及冷空气的影响,寒害仍是橡胶树栽培与发展的限制性因子[1]。橡胶树在不同类型的寒害情况下,主要表现为枝叶干枯、茎干黑斑、树体各部位爆皮流胶等受害症状[2]。因寒害导致的茎干爆皮流胶不仅会导致树皮溃烂坏死,还会引起后期小蠹虫、病原菌的入侵,造成寒害后的次生病虫害,致使橡胶树的抗逆性下降,并可加重因寒害造成的经济损失[3-4]。因此,探索不同橡胶树品种耐寒性及低温所致爆皮流胶的差异对于橡胶树抗寒栽培管理措施的实施以及爆皮流胶机制研究至关重要。【前人研究进展】近年来,以生理指标反映橡胶树耐寒能力的应用较为广泛。李玲等[5]低温胁迫2个橡胶树品种芽接苗,测定其电导率并进行恢复,根据2个品种低温下表现及不同组织电导率的变化情况,确定了较为抗寒的品种。孔广红等[6]以低温胁迫下,叶片活性氧相关成分含量、抗氧化相关物质含量及酶类活性高低来反映橡胶树耐寒与非耐寒品种间的差异。结合隶属函数法及主成分分析法分析低温下橡胶树不同品种(系)的生理指标差异,不仅可以比较不同品种(系)自然低温下的抗寒能力[7-9],而且可以判断人工模拟不同类型寒害条件下,不同橡胶树品种的耐寒性强弱[10]。【本研究切入点】由于自然及人工模拟低温条件下,橡胶树幼苗极少发生爆皮流胶寒害症状[11],所以将低温下生理生化指标与爆皮流胶情况的差异相结合来研究橡胶树不同品种寒害与爆皮流胶的耐受性鲜有报道。【拟解决的关键问题】本研究选择热研7-33-97、热研7-20-59、湛试327-13、93-114等4个橡胶树品种一年生芽接苗为材料,结合设置的人工模拟低温条件,测定低温下其叶片叶绿素荧光参数和部分生理生化指标的变化差异,通过隶属函数法、主成分分析法对其耐寒性进行比较,并结合低温条件所致爆皮流胶的情况分析,以期探索橡胶树爆皮流胶耐受性的机制,为完善橡胶树抗寒评价体系和抗寒栽培技术研发提供理论依据。
1 材料与方法
1.1 材料
供试材料为国家热带植物种质资源库—橡胶树(National Tropical Plants Germplasm Resource Center-Rubber Tree)提供的一年生芽接苗(2020年5月芽接,高度为0.8~1.0 m),品种为热研7-33-97、热研7-20-59、湛试327-13及93-114。
1.2 研究方法
1.2.1 试验设计 选择长势基本一致的4个橡胶树芽接苗(每个品种20株)品种,放置于人工气候室内进行低温胁迫处理。在低温胁迫的第0、2、4、6天测定各品种各植株相同部位叶片的叶绿素荧光参数和生理生化指标,0 d作为对照。低温胁迫6 d后,在人工气候室内恢复1周,将所有植株移入自然环境中。从植株开始出现流胶症状时至移入自然环境条件前,记录每个品种流胶情况。在移入自然环境55 d后,统计其存活情况。低温胁迫期间人工气候室的控制条件为:12 h光照,12 h黑暗,相对湿度为80%;光照强度为75~100 μmol/(m2·s);光照时段温度控制在(16±1)℃,黑暗时段温度控制在(4±1)℃。恢复期间控制条件为:除温度控制在(26±1)℃之外,其他同于低温胁迫时情况。
1.2.2 测定方法 采用WALZ PAM-2500便携式叶绿素荧光仪测定叶片最大光化学效率(Fv/Fm)。各时间梯度生理指标测定时各品种各植株至少选择2个叶片,各品种各时间梯度至少选取3个植株的叶片,各品种各时间梯度的叶片混合后设置3个重复。参照李小琴等[8]的方法,采用METTLER TLEDO-FE38电导率仪测定电导率。叶绿素含量采用混合液法[12]测定。采用苏州科铭生物技术有限公司的试剂盒测定橡胶树植株叶片可溶性糖含量、可溶性蛋白含量、丙二醛含量、SOD活性。采用广陆数显游标卡尺(111N-101-10G)测定橡胶树芽接苗流胶情况(点状流胶测流胶点的直径,条状流胶测流胶的长度和宽度)后计算其面积。
1.3 数据处理
叶绿素荧光参数及生理生化指标数据为3次重复的平均值,采用Excel和DPS进行数据统计与分析,多重比较采用Duncan’s新复极差法,用OriginPro 2018进行绘图;用隶属函数法[13]对4个橡胶树品种芽接苗进行耐寒性比较,用IBM SPSS软件进行主成分分析。
2 结果与分析
2.1 低温胁迫下不同品种叶片生理生化指标的变化
2.1.1 相对电导率和丙二醛含量 由图1-A可知,低温胁迫的过程中,4个橡胶树品种与各自对照相比,相对电导率均增加,各品种相对电导率呈先升高后下降趋势。但是,有别于热研7-20-59、湛试327-13和93-114 3个品种相对电导率逐步升高而后下降趋势,热研7-33-97相对电导率表现为陡然升高后逐步降低,它在第2天的相对电导率比其对照升高69.31%,均高于热研7-20-59、湛试327-13和93-114 3个品种的27.80%、17.84%和40.10%;而后在第4~6天,热研7-33-97相对电导率与其对照相比,变化幅度均小于其他3个品种。在低温过程中,热研7-33-97相对电导率均显著高于热研7-20-59。
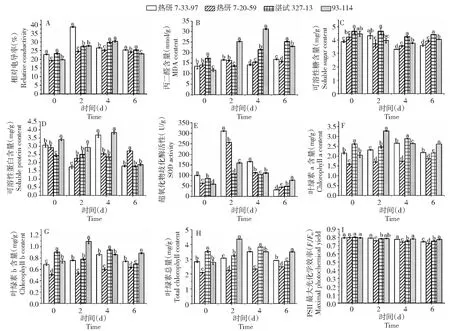
数值为3个数值的平均值,误差棒为标准差,小写字母表示相同时间梯度下品种间存在显著差异(P<0.05)
2.1.2 可溶性糖和可溶性蛋白含量 如图1-C所示,在受低温的过程中,除热研7-33-97可溶性糖含量为“升—降—升”趋势外,另外3个品种的可溶性糖含量均为先降后升趋势。胁迫4~6 d的过程中,4个品种可溶性糖含量均低于其对照。低温下,湛试327-13可溶性糖含量最高,而且变化幅度也是最小,与其对照相比,其降低幅度仅为0.08%、8.78%和5.43%,均低于其他3个品种。在胁迫的4~6 d,4个品种间可溶性糖含量的差异均达到显著水平(P<0.05),湛试327-13和93-114的可溶性糖含量均高于热研7-33-97和热研7-20-59。
由图1-D可以看出,受低温的影响,4个品种可溶性蛋白含量变化的趋势不一致:热研7-33-97和93-114呈“降—升—降”趋势,在低温的第4天,热研7-33-97和93-114的可溶性蛋白含量增幅均较大,热研7-33-97可溶性蛋白含量与其对照相比,增幅为20.12%,93-114与其对照相比,增加12.43%;热研7-20-59呈先降后升趋势,与其对照相比,可溶性蛋白含量均降低;湛试327-13呈先升后降趋势,在低温的第2天,可溶性蛋白含量增加,后期又呈现逐步下降趋势。
2.1.3 超氧化物歧化酶活性 如图1-E所示,低温条件下,4个品种的超氧化物歧化酶活性的变化趋势保持一致,均为先升高后下降的趋势。低温条件的第2天,与各自对照相比,热研7-33-97、热研7-20-59、湛试327-13、93-114超氧化物歧化酶活性增幅最大,分别为其对照的2.11倍、3.10倍、26.09%、1.75倍。低温条件的第6天,虽然93-114超氧化物歧化酶活性有所下降,但是与其对照相比,其变化幅度仍然增加,而其他3个品种与各自对照相比变化幅度均降低。同时,除了低温胁迫的第4天,其他时间梯度,4个品种超氧化物歧化酶活性的差异均达到显著水平(P<0.05)。
2.1.4 叶绿素含量 由图1-F、1-G、1-H可见,低温胁迫下,热研7-33-97叶绿素a、叶绿素b、叶绿素总量为先升高后下降的趋势,低温的第4天,其值最高。热研7-20-59叶绿素a、叶绿素b、叶绿素总量一直呈现逐步增加的趋势,可能是低温逆境下叶片失水所致,但是其含量均显著低于其他3个品种。湛试327-13 叶绿素a、叶绿素b、叶绿素总量均呈“降—升—降”趋势,低温的第4天,其叶绿素a含量、叶绿素b含量、叶绿素总量最高。93-114叶绿素a、叶绿素总量呈先升后降趋势,而叶绿素b含量呈“升—降—升”趋势,低温胁迫的第2、6天,其叶绿素a含量、叶绿素b含量、叶绿素总量最高。与其对照相比,93-114叶绿素a含量、叶绿素总量增加幅度均高于其他3个品种,分别为60.13%、28.53%、27.50%和56.50%、25.50%、25.26%。
2.1.5 PSII最大光化学效率 由图1-I可得知,随着低温时间的延长,4个品种的PSII最大光化学效率呈逐步下降的趋势,热研7-33-97、湛试327-13和93-114的PSII最大光化学效率均高于热研7-20-59,其差异达到显著水平(P<0.05)。与各自对照相比,热研7-33-97的 PSII最大光化学效率降低幅度分别为0.28%、2.42%、6.17%,均小于热研7-20-59的5.08%、6.96%、9.09%;93-114 的 PSII最大光化学效率降低幅度分别为1.06%、1.61%、2.60%,均小于湛试327-13的2.33%、5.78%、5.87%。
2.2 隶属函数法比较橡胶树品种耐寒性差异
采用隶属函数法对4个橡胶树品种低温胁迫下各项生理生化指标分别进行计算,比较其耐寒性。由表1可以看出,4个品种的平均隶属度从大到小的排序为: 93-114>湛试327-13>热研7-33-97>热研7-20-59,但是93-114 和 湛试327-13的差值并不大。
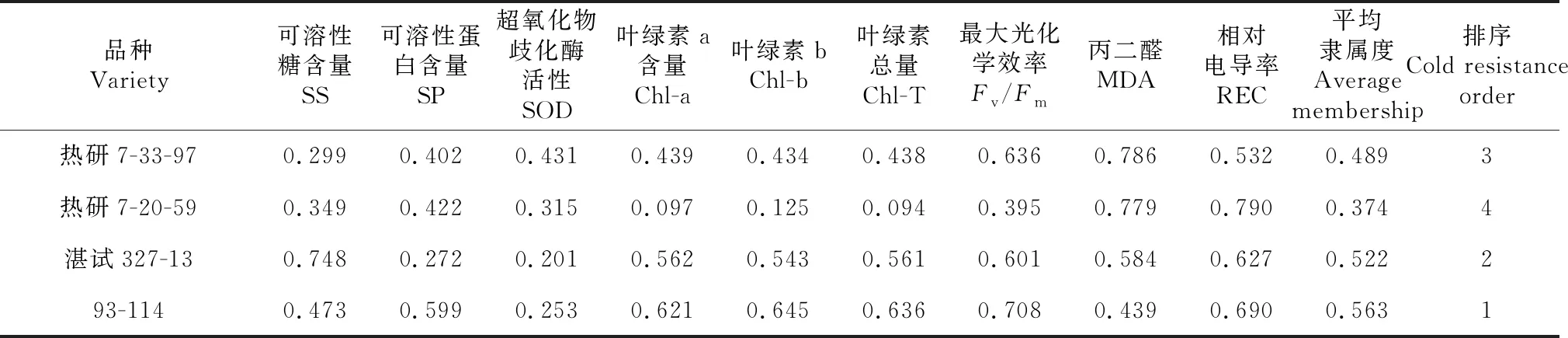
表1 不同品种生理生化指标隶属函数值及平均隶属度
2.3 主成分分析法比较橡胶树品种耐寒性差异
对4个橡胶树品种生理生化指标进行主成分分析,其结果见表2。由主成分分析结果可以看出:前3个主成分的特征值分别为5.330、2.086和1.584,贡献率分别为59.226%、23.173%和17.600%,前3个主成分的累计贡献率为100%,可以反映低温胁迫下橡胶树生理生化指标的全部信息。由表3可以看出:第1主成分中,具有较高荷载的为叶绿素a含量、叶绿素b含量、叶绿素总量及最大光化学效率;第2主成分中,具有较高荷载的为可溶性糖含量、SOD活性和相对电导率;第3主成分中,具有较高荷载的为可溶性蛋白含量。

表2 主成分特征值及累计贡献率
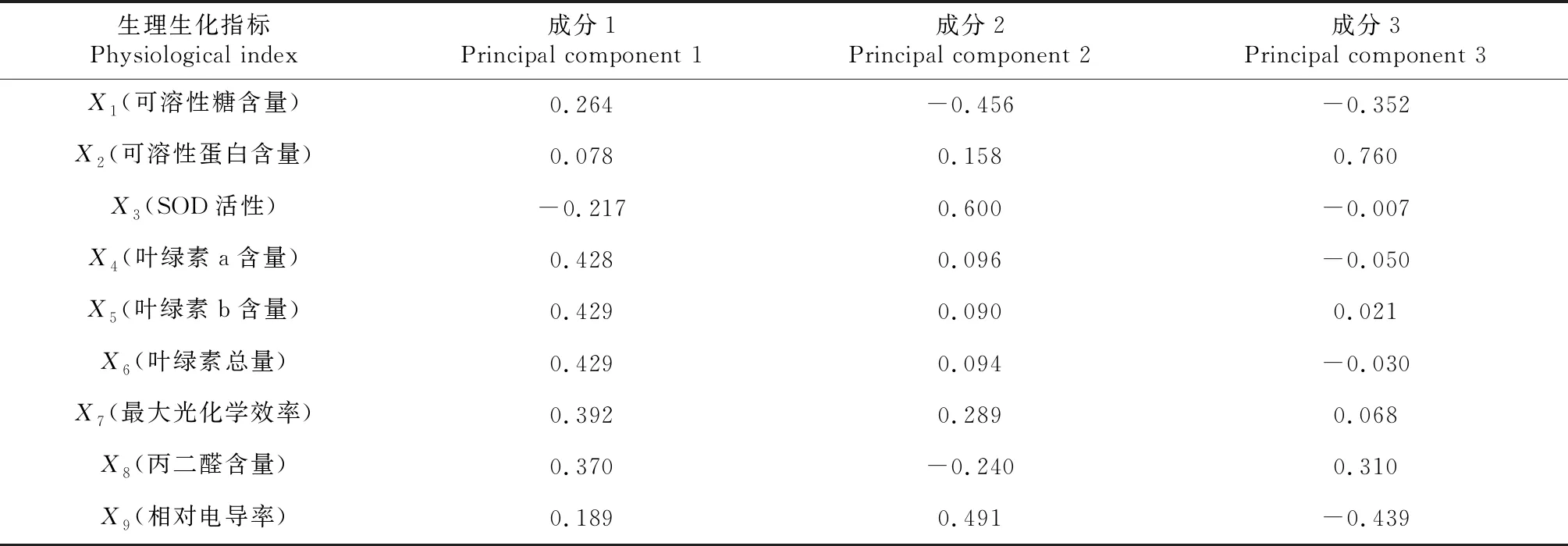
表3 主成分荷载矩阵
根据主成分分析结果,可得出3个主成分函数表达式。
果然周副书记很老到,他说,我带你到几个部门走走。几天来,周书记马不停蹄地带到我找电力、税务、银行、火车站等部门单位化缘,又找到镇上很有些实力企业老板拉了些赞助,五万元的经费基本上没问题。
Y1=0.264X1+0.078X2-0.217X3+0.428X4+0.429X5+0.429X6+0.392X7+0.370X8+0.189X9
Y2=-0.456X1+0.158X2+0.600X3+0.096X4+0.090X5+0.094X6+0.289X7-0.240X8+ 0.491X9
Y3=-0.352X1+0.760X2-0.007X3-0.050X4+0.021X5-0.030X6+0.068X7+0.031X8-0.439X9
由3个主成分函数表达式,分别计算出4个品种耐寒性的综合得分值:Y=0.592Y1+0.232Y2+0.176Y3。由表4可知,低温胁迫下,不同橡胶树品种主成分综合得分排序为:93-114 >湛试327-13>热研7-33-97>热研7-20-59。
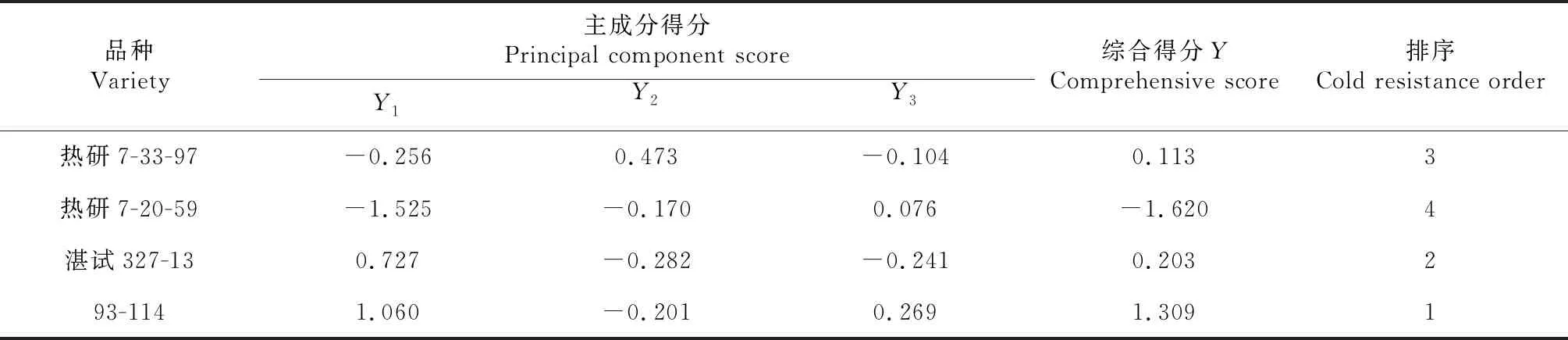
表4 不同橡胶树品种主成分得分及排序情况
2.4 不同橡胶树品种爆皮流胶差异性比较
在低温胁迫的第4天,橡胶树芽接苗植株开始出现爆皮流胶症状。流胶产生于植株绿色茎干的叶痕、芽眼、节间等位置,或者出现在总叶柄、总叶柄的叶枕、腋芽处。与此同时,热研7-33-97和湛试327-13植株茎干流胶伴随出现黑斑;热研7-33-97和93-114叶片出现黑斑。由表5可以看出,低温胁迫下,热研7-20-59流胶植株数最多,热研7-33-97最少。橡胶树各品种总流胶个数与植株存活率情况与隶属函数法及主成分分析法得出的品种耐寒性保持一致;而流胶面积大小,即流胶量的大小与耐寒性并不一致,这可能与品种胶乳含量、干胶含量、总固形物含量等因素有关。
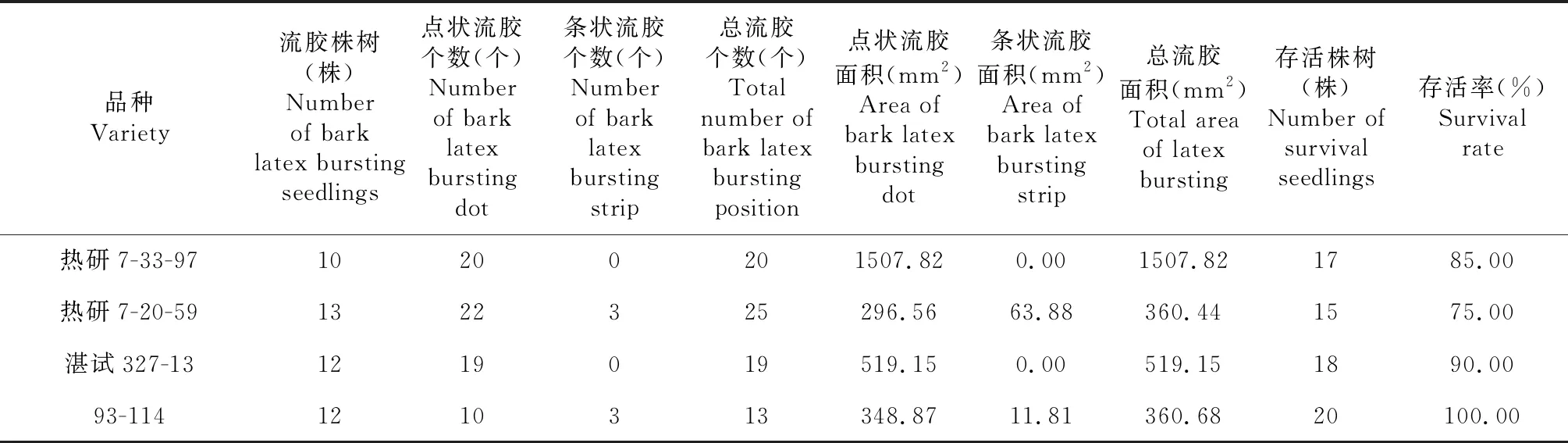
表5 不同橡胶树品种流胶差异情况比较
3 讨 论
在不利的温度环境下,植物受影响最大的细胞器就是叶绿体,异常的环境温度导致光合电子传递性能的降低及影响PSII,使得多余能量产生一系列活性氧,光合作用和叶绿素合成均因活性氧受到抑制。活性氧不仅影响氧化还原信息转导,而且它的积累会对细胞产生毒害[14]。作为评价光合能力的重要指标,叶绿素在收集、传导、转化、吸收光能中具有重要作用[15],叶绿素减少会降低植物光保护的性能[16]。 另一方面,叶绿素的合成涉及一系列的酶促反应,过程较为复杂。环境温度一旦下降,由于碳固定减慢,导致叶绿体中的“激发压力”增加,库容量就会减少[17],叶绿素合成中相关催化酶活性也受影响,所以叶绿素合成与功能发挥受抑制程度存在差异,以致本研究中不同橡胶树品种之间叶绿素a、叶绿素b、叶绿素总量变化也存在着差异。PSII最大光化学效率(Fv/Fm)常被用作植物逆境条件下抗性高低的重要指标[18]。当光能超过植物光合作用所需时,光合效率就会大幅下降,出现光抑制,尤其是在逆境下,植物的光抑制最为明显[19]。本研究中,低温胁迫下,热研7-20-59的最大光化学效率均小于其他3个品种,也就意味着其对低温最为敏感,4个橡胶树品种的PSII受低温的影响,其开放程度明显降低,光合性能受到影响[20]。一旦植物受不利温度的胁迫,叶绿体受损伤,导致光合电子传递性能降低及PSII受抑制,光能转化为热能,植物借助热耗散消耗多余热量,拮抗过剩能量对其本身的损伤,多余的能量就以活性氧的形式散发出来,致使细胞脂膜过氧化产生丙二醛,丙二醛的积累反过来加剧植物受损伤的程度[21]。随着低温胁迫时间的延长,与各自对照相比,4个橡胶树品种叶片丙二醛(MDA)含量最终均达到显著水平,也就意味着低温对各品种植株细胞膜均造成了损伤[22],细胞膜的透性就逐步增大,胞内电解质外渗随之增加,即相对电导率上升[23]。在低温下,4个橡胶树品种植株叶片相对电导率与各自对照相比,其电导率均呈增加的趋势,但后期由于活性氧清除系统所涉及的渗透调节物质及相关酶的作用,植物拮抗低温逆境的防御功能逐步显现。作为渗透调节物质之一,可溶性糖可维持细胞液的流动性与细胞功能的正常性,降低植物在低温下的伤害[24]。在低温环境下,湛试327-13植株叶片可溶性糖含量是最高的,而且变化幅度也是最小的。而且湛试327-13和93-114植株叶片可溶性糖含量均高于其他2个品种,所以其耐寒性与其可溶性糖含量较高是密不可分的。作为亲水性胶质,可溶性蛋白可降低细胞冰点[25],参与低温逆境信号转导、细胞修复调控等[26],会随着植物低温驯化而升高[27]。本研究中,与各自对照相比,除了热研7-20-59以外,其他3个橡胶树品种叶片可溶性蛋白(SP)在低温下均出现了增幅的峰值,所以可溶性蛋白积累在降低热研7-33-97、湛试327-13和93-114 3个品种的渗透势,提高它们的耐寒性方面起到了较好的作用。逆境下,SOD可清除植物细胞内产生的活性氧,维持细胞完整性[28],从而提高植物的抗逆性[29]。低温逆境中,SOD活性被激活,植物的耐寒性提高,植物产生了对低温的自我保护能力,但是随着低温过程的延长,植物受低温影响程度加大,SOD活性而逐步下降[30],这与本研究中4个橡胶树品种SOD活性的变化趋势是一致的。
橡胶树的根、茎、叶(叶片、小叶柄、大叶柄)中均存在产生胶乳的乳管,但在茎干中最多[31]。由此,本研究中4个橡胶树品种在植株的茎干、叶部位均发生了爆皮流胶症状。不同品种乳管个数存在差异,胶乳浓度(干胶含量、总固形物含量)[32-33]和因植株营养情况而致的凝固酶的差异,导致其流胶性能的差异[31]。另一方面,受低温胁迫的影响,可能由于细胞壁中纤维素、半纤维素和木质素等各组成成分在细胞壁中的分布、结合方式以及各组成成分本身的性能改变造成植物细胞壁及宏观力学性能的改变[34],以致于不同橡胶树品种低温下乳管破裂的方式以及破裂程度不同,最终导致4个橡胶树品种流胶株数、流胶面积与其耐寒性并不完全一致。
以单一生理生化指标判断植物的耐寒性较为困难,因为同一物种的不同品种之间的同一生理生化指标变化规律并不完全相同,以单一生理生化指标来判断品种间的耐寒性势必造成评判的片面性及结果的差异、不客观性[35]。与此同时,由于橡胶树耐寒性是较为复杂的数量性状,而且对不同类型的低温寒害的表现在品种间的差异较大[36]。因此,结合隶属函数法、主成分分析法对本研究中的4个橡胶树品种耐寒性进行综合评价,可以避免单一生理生化指标评判的局限性和各指标信息的重叠性,使结果更加客观、全面[37]。
4 结 论
本研究通过隶属函数法、主成分分析法对低温胁迫下4个橡胶树品种叶片相对电导率、丙二醛含量、可溶性糖含量、可溶性蛋白含量、超氧化物歧化酶活性、叶绿素含量及最大光化学效率等指标进行其耐寒性的综合评价。结果表明,隶属函数分析法与主成分分析法评价的耐寒性顺序为:93-114 >湛试327-13> 热研7-33-97>热研7-20-59。因低温导致的4个品种流胶株数的排序是:热研7-33-97<湛试327-13 = 93-114<热研7-20-59。从以上结果可以看出,93-114和湛试327-13是较好的耐寒性橡胶树品种,适合于寒害较为严重的地区种植,热研7-33-97是胶乳含量较高、耐寒性中等的橡胶树品种,适合于寒害不太严重地区种植[38]。橡胶树不同品种因低温所致的爆皮流胶的耐受性机制及爆皮流胶耐受性与耐寒性的关系有待进一步探索。
