云南杞麓湖浮游植物的群落特征及相关因子的研究
2022-12-23江卓韵潘晓赋罗旭孔德平吴松刘淑伟褚荣波王茉
江卓韵潘晓赋罗旭孔德平吴松刘淑伟 褚荣波王茉
(1.西南林业大学生物多样性保护学院,云南省高校极小种群野生动物保育重点实验室,云南昆明 650233;2.中国科学院昆明动物研究所遗传资源与进化国家重点实验室,云南昆明 650223;3.云南省生态环境科学研究院,云南昆明 650034;4.云南亚美湖泊水质治理有限公司,云南玉溪 652700)
浮游植物在湖泊生态系统食物链和水体生产力中的重要作用,以及对水体中许多物质的敏感性,使其在湖泊生态系统的演替和稳定研究中具有重要意义[1]。作为湖泊生态系统中的初级生产者,浮游植物通过光合作用产生的氧气是水体溶解氧的主要来源,将光能转化为生物能并储存无机碳,是物质循环和能量流动的关键过程[2-3]。浮游植物还与环境因子有着复杂的关系,其群落的种类组成、细胞丰度和物种多样性会随着物理、化学和生物因素的改变而产生不同程度的变化[4-7]。在不同条件下,影响浮游植物群落结构变化的生态因素也不尽相同,其主要驱动因素也存在一定差异[8]。
杞麓湖位于云南省玉溪市通海县,属于珠江流域。该湖东西较长,南北较窄,四周群山环抱,是一个典型的高原封闭新月形断陷盆地[9-10]。湖区水资源短缺,还处于面源污染的聚集地,长期的污染导致生态环境脆弱,流域内污水治理速度赶不上水污染速度,从而严重制约该流域的可持续发展[10-12]。此外,杞麓湖关于浮游植物生态学方面的研究既不全面也欠缺连续性。从20世纪80年代开始,有研究人员初步了解到该湖浮游植物的优势种为微囊藻(Microcystis)[13]。20世纪末到21世纪初,研究人员结合藻类和水质进行监测,发现杞麓湖水体中藻量减少,透明度增加,而造成这种现象的具体原因并不清楚[14]。也有多位学者对包括杞麓湖在内的13个云南中南部湖泊进行浮游植物和水资源调查[15-16],显示这些湖泊的浮游植物总生物量与温度显著相关,但均未单独对杞麓湖进行深入细致的研究。直到杨鸿雁等[17]在2017—2018年针对杞麓湖浮游生物群落的季节性演替进行了初步研究,结果显示浮游植物和浮游动物季节性演替均受温度影响。纵观以往对杞麓湖浮游植物的研究,或仅集中在群落特征的方向上,或有少数结合2种类群的时期变化并分别探讨与环境因子关系。目前尚未见有针对湖内浮游植物与浮游动物、理化因子之间耦合关系的研究。
由于湖区生态环境破坏和水资源污染等问题,使作为九大湖泊之一的杞麓湖成为中央环保督察组督察重点之一[18-19]。浮游植物是重要水质指示生物,通过掌握其群落结构的演替规律,分析环境因子对浮游植物群落结构的影响,有助于了解湖泊生态系统的健康状况。因此,本研究结合生物和非生物因子,通过了解浮游植物群落组成和关键理化因子的时相变化,分析杞麓湖浮游植物群落结构的演替规律,初步探讨湖内关键理化因子以及浮游动物对浮游植物群落结构的影响,深入揭示该湖浮游植物群落周年演替的动力因素,进一步完善杞麓湖浮游植物生态学方面的研究,以期为湖泊生态和生物多样性的恢复提供参考依据。
1 材料和方法
1.1 采样点和采样时间
采样时间是2020年1—12月,每月采样1次。根据湖库面积和湖盆,兼顾湖泊中心、湖湾和入流等区域,考虑排污设施的位置和人为活动影响程度等因素[20],选取了6个采样点(图1),分别是S1(湖管站)、S2(红旗河口)、S3(湖北岸)、S4(湖心)、S5(马家湾)和S6(落水洞)。

图1 杞麓湖水样采集样点分布图Fig.1 Distribution of water sampling site in Qilu Lake
1.2 样品测定与数据收集
1.2.1 关键理化因子检测数据收集
水样样品用5 L采水器采集水面下0.5 m左右混合水样5 L,低温保存运回实验室进行理化因子的检测。总氮(TN)、总磷(TP)、化学需氧量(CODcr)和氨氮(NH3-N)在实验室内进行检测[20]。
1.2.2 浮游生物鉴定及相关数据收集
浮游生物的定性和定量的采集分析方法均参照SL167-2014[21]。定性样品采集用25号浮游生物网采表层0.5 m左右处的水样后固定,用以保存浮游植物。定量样品分为2种:一种是采集湖面表层0.5 m左右的混合水样1 L后固定,静置48 h,用虹吸法浓缩至50 mL,用以保存浮游植物、原生动物和轮虫;另一种是用5 L采水器采集10 L混合水样,水样经25号浮游生物网过滤后浓缩至50 mL固定,以保存桡足类和枝角类。所有样品均以样品体积与鲁哥试剂为100∶1的比例进行固定。
定性样品分析使用德国Carl-Zeiss,Axio Imager. A2显微镜在10~100倍物镜下基于传统形态学的鉴定方法。浮游植物和原生动物、轮虫、桡足类和枝角类定量样品的分析分别使用0.1、1 mL和5 mL的计数框,使用视野法鉴定计数。浮游植物按照细胞个数计数,枝角类和桡足类将样品分为若干次全部计数。
浮游生物的物种鉴定参照《中国淡水生物图谱》[22]《中国淡水藻类:系统、分类及生态》[23]和《淡水微型生物图谱与底栖动物(第二版)》[24]等分类书籍。
1.3 分析方法
1.3.1 数据处理
浮游生物丰度的计算根据SL167-2014[21]。浮游植物生物量的计算根据浮游植物的细胞相对密度大多接近于1,细胞体积(mL)相当于细胞质量(g),直接由体积换算为生物量(湿质量)[1]。原生动物和轮虫生物量的计算采用体积换算法,根据不同种类的体形,按最近似的几何形测量其体积;枝角类和桡足类生物量的计算采用测量不同种类的体长,用回归方程式求体重进行[19]。
浮游植物优势度(Y)使用Dufrêne法计算,Y>0.02的物种为优势种[25]。浮游植物多样性指数分别计算Shannon-Wiener多样性指数(H′)[26]和Pielou均匀度指数(J)[27]。
1.3.2 数据分析
分析浮游植物群落结构的时相差异。首先对于不同月份浮游植物丰度、生物量和多样性指数的差异使用非参数Kruskal-Wallis检验。其次,基于Bray-Curtis距离矩阵,采用ANOSIM分析不同月份间浮游植物物种平均丰度的差异。最后,使用SIMPER分析,选取造成群落差异贡献率为前5的种类。
分析理化因子、浮游动物对浮游植物群落结构的影响。首先,对数据进行log (x+1)的转换,使用线性回归分析关键理化因子、浮游动物总丰度对浮游植物总丰度的影响。然后,使用DCA分析,得到前4个轴中最大值小于2,所以选择RDA进行相关分析。
所有的数据分析和图形绘制通过Origin Lab.2021和R 3.6.2进行。
2 结果与分析
2.1 浮游植物群落结构特征
2.1.1 种类组成、丰度、生物量和多样性指数及其时相变化
本研究共鉴定出浮游植物147种,隶属于8门75属。主要包括绿藻门(77种,占比52.38%)、硅藻门(31种,占比21.09%)和蓝藻门(21种,占比14.29%),其余裸藻门、黄藻门、金藻门和甲藻门共18种,共占比12.24%。
杞麓湖浮游植物丰度和生物量的时相变化范围 分别 为6.20×107~46.09×107个/L和9.33~94.37 mg/L;两者均值的时相变化相似,呈现单峰趋势。由图2可知,在丰度和生物量上,蓝藻门全年均处于绝对优势;蓝藻门丰度和生物量占比全年均高至50%以上,其所占比例的时相变化范围分别为65.31%~92.78%和53.34%~96.88%。
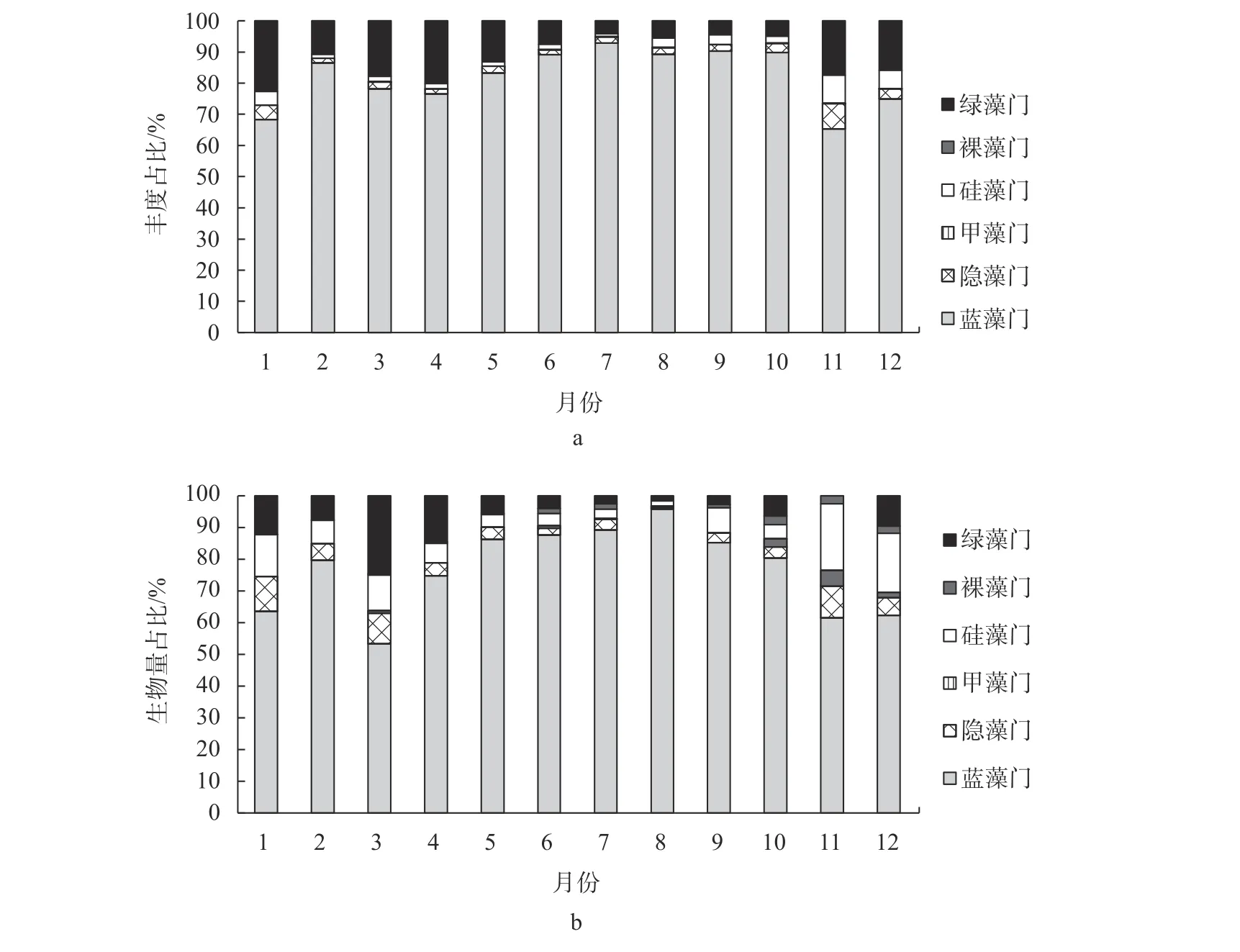
图2 杞麓湖浮游植物各类群丰度和生物量比例的时相变化Fig.2 Period variation of the proportion of phytoplankton abundance and biomass in Qilu Lake
浮游植物多样性指数的时相变化均呈波浪形,见图3。浮游植物Shannon-Wiener多样性指数(H′)(图3c)和Pielou均匀度指数(J)(图3d)的时相变化范围分别为2.19~3.59和0.36~0.58。其中,Shannon-Wiener多样性指数的值越大,群落结构越复杂,群落稳定性也会相应增加[28]。Pielou均匀度指数反映某个群落或生境中各物种多度分布的均匀程度[29]。而杞麓湖浮游植物多样性指数在春冬季偏高,夏秋季偏低。因此,春冬季浮游植物群落稳定性强,均匀程度高;夏秋季群落稳定性相对弱,均匀程度较低。
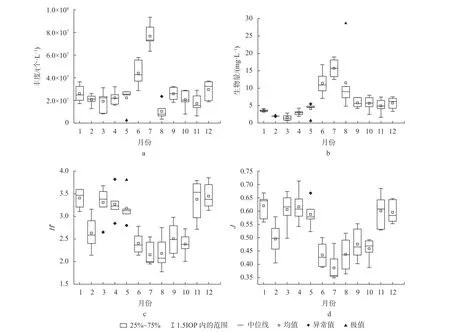
图3 杞麓湖浮游植物丰度、生物量和多样性指数的时相变化Fig.3 Period variation of phytoplankton abundance, biomass and diversity indices in Qilu Lake
2.1.2 优势物种的特征
不同月份杞麓湖浮游植物优势物种共17种,隶属于4门。根据杞麓湖浮游植物优势物种的时相变化特征可以分为阿氏浮丝藻(Planktothrix agardhii)主导时期和水华束丝藻(Aphanizomenon flosaquae)主导时期,其中阿氏浮丝藻是1—4月份和8月份优势度最高的优势物种,且这段时期阿氏浮丝藻丰度占比的范围在8.39%~36.36%。水华束丝藻是5—7月份和9—12月份优势度最高的优势物种,这段时期其丰度占比的范围在13.96%~36.35%。
2.1.3 不同月份群落结构差异
通过非参数Kruskal-Wallis检验分析不同月份之间浮游植物的群落结构,结果显示不同月份间浮游植物丰度、生物量和多样性指数均存在显著差异(P< 0.05)。
通过ANOSIM分析不同月份间(66组)浮游植物平均丰度的差异,结果显示各月份之间组间差异均大于组内差异(R>0),其中有64组月份间浮游植物平均丰度差异显著(P<0.05)(表1)。选择其中浮游植物平均丰度差异大且显著(均为R>0.9,P<0.05,表1中加粗的数据)的16组进行SIMPER相似性分析(表2),结果显示阿氏浮丝藻和水华束丝藻是造成杞麓湖浮游植物群落结构差异显著的主要物种。

表1 杞麓湖不同月份间浮游植物平均丰度的ANOSIM分析Table 1 ANOSIM analysisof phytoplankton average abundance in different monthsin Qilu Lake
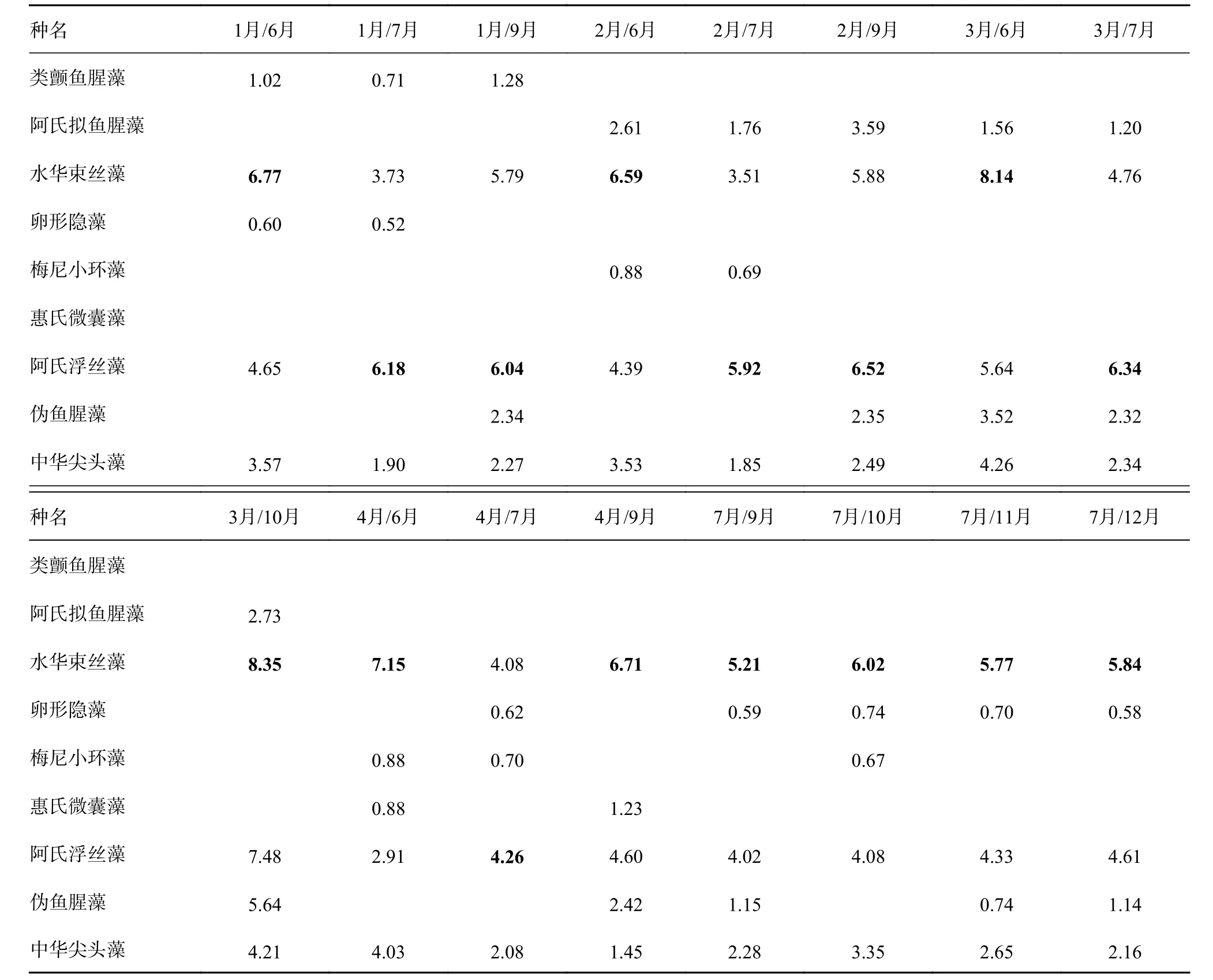
表2 杞麓湖16组月份间浮游植物平均丰度的SIMPER相似性分析Table 2 SIMPER similarity analysisof phytoplankton average abundance in 16 grouped months in Qilu Lake%
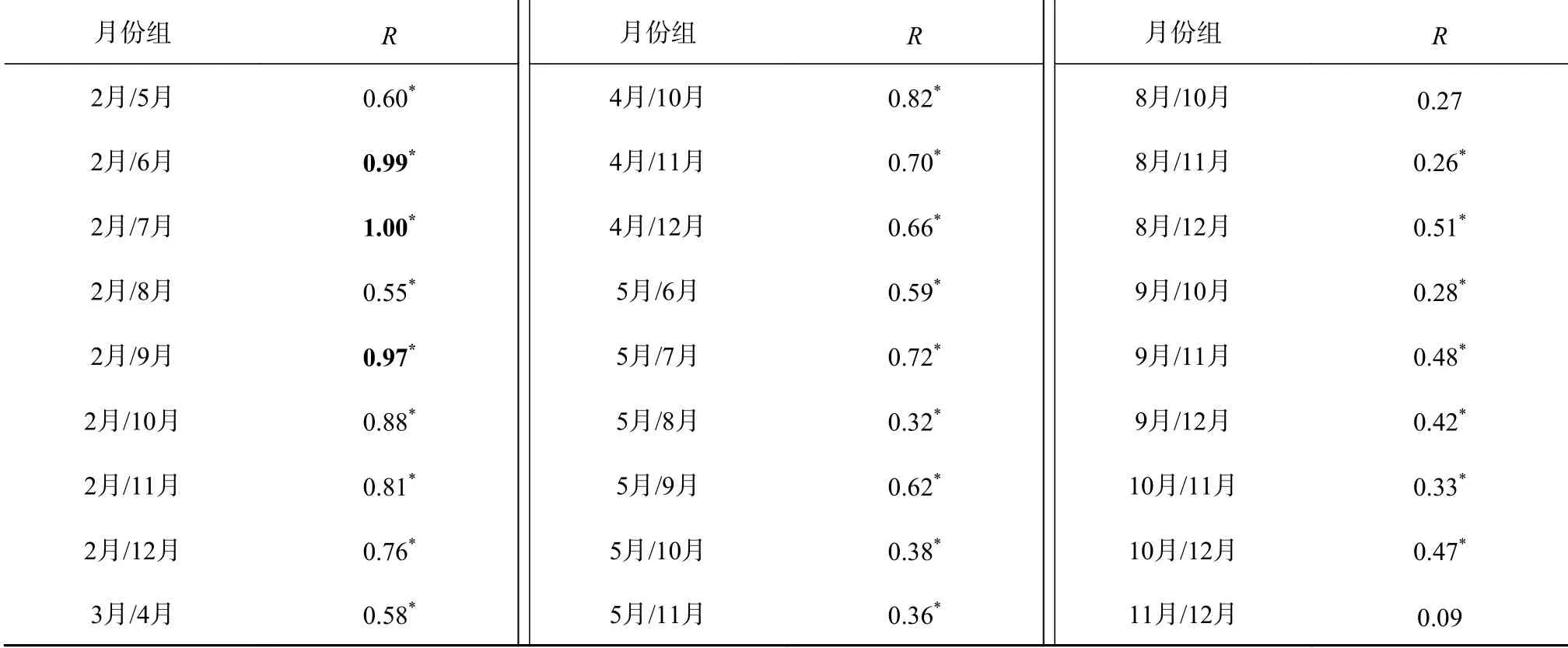
续表 1
2.2 关键理化因子的时相变化特征
杞麓湖水体中总氮(TN)和化学需氧量(CODcr)的浓度均指示该湖水体的水质为V类[30]。CODcr的浓度范围为48.67~59.33 mg/L,其时相变化呈下降趋势(图4a);氨氮(NH3-N)的浓度范围为0.31~1.12 mg/L,其时相变化呈波动上升趋势(图4a);总氮(TN)和总磷(TP)的浓度范围分别为2.93~9.66 mg/L和0.10~0.25 mg/L,两者均呈现先升后降的时相变化,均在5月份达到最高值(图4b)。杞麓湖还表现出高氮低磷的特征,总氮与总磷比例(TN∶TP)范围在14~40之间,其时相变化不稳定(图4b)。
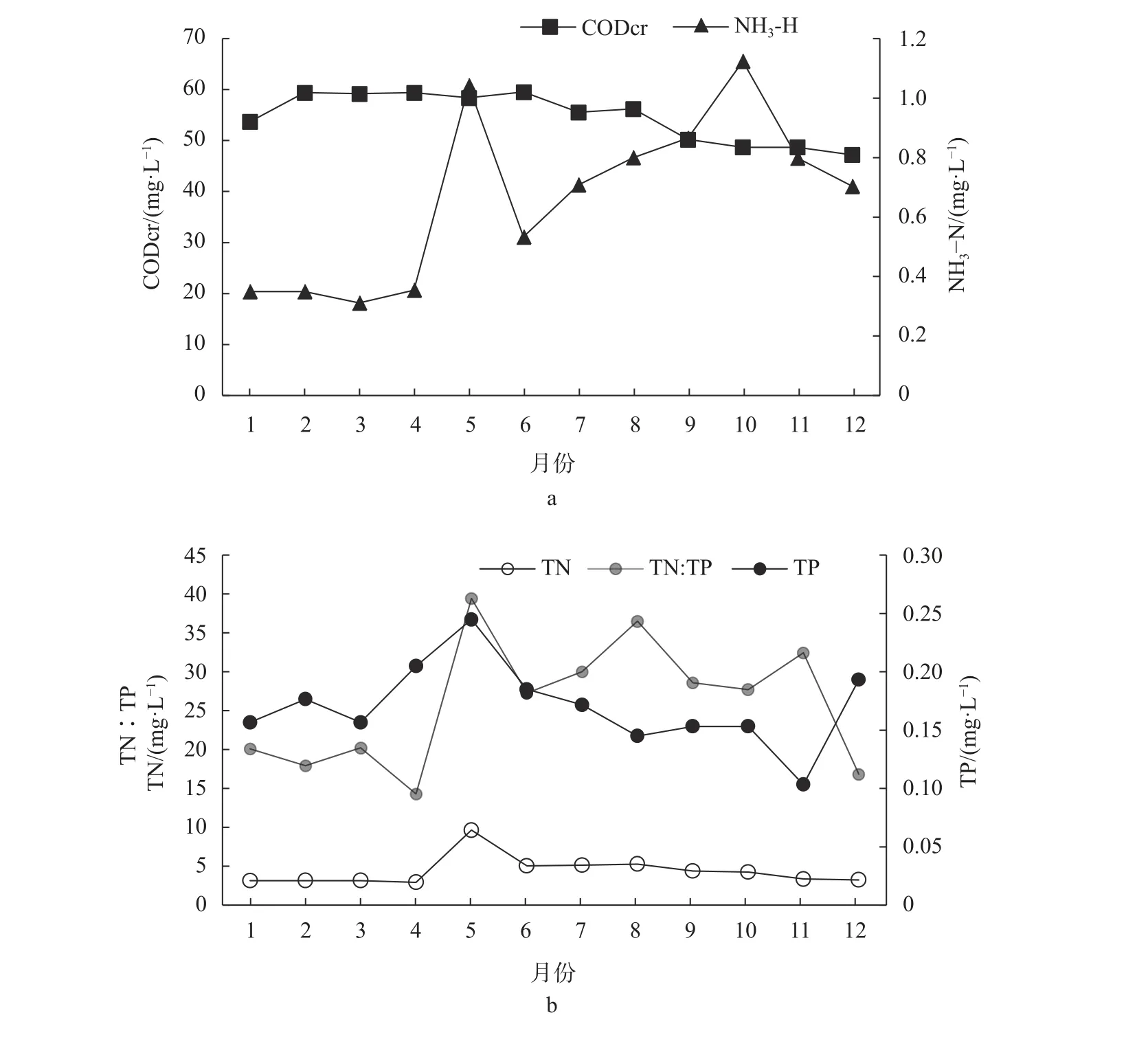
图4 杞麓湖关键理化因子的时相变化Fig.4 Period variation of key physicochemical factorsin Qilu Lake
2.3 浮游植物与关键理化因子、浮游动物的关系
本研究调查得到杞麓湖浮游动物的总丰度为15.40× 104个/L,总生物量为27.29 mg/L。其中,轮虫和原生动物在数量上占主要优势。通过以浮游动物总丰度和关键理化因子作为自变量,浮游植物总丰度为因变量,进行线性回归分析,结果显示仅环境因子中总氮(TN)、总磷(TP)以及TN/TP对浮游植物总丰度有显著影响(P<0.05)。
通过对造成不同月份间浮游植物群落结构差异贡献前5的物种的平均丰度与关键理化因子、浮游动物平均丰度的相关分析,结果(图5)显示,杞麓湖浮游植物受到理化因子和浮游动物的共同影响,其中在数量上占主要优势的阿氏浮丝藻和水华束丝藻,分别受到化学需氧量(CODcr)和原生动物中钟虫(Vorticellasp.)的影响。此外,通过分析不同月份间浮游植物各类群的丰度和生物量所占比例与关键理化因子、浮游动物各类群丰度和生物量所占比例,结果显示(图6),蓝藻门生物量所占比例与总氮(TN)成正相关,隐藻门生物量所占比例与总氮(TN)成负相关。

图5 造成杞麓湖不同月份浮游植物丰度差异贡献前5的物种与环境因子之间的相关分析Fig.5 Correlation analysisof top 5 phytoplankton speciesin abundancewith environmental factors in Qilu Lake in different months
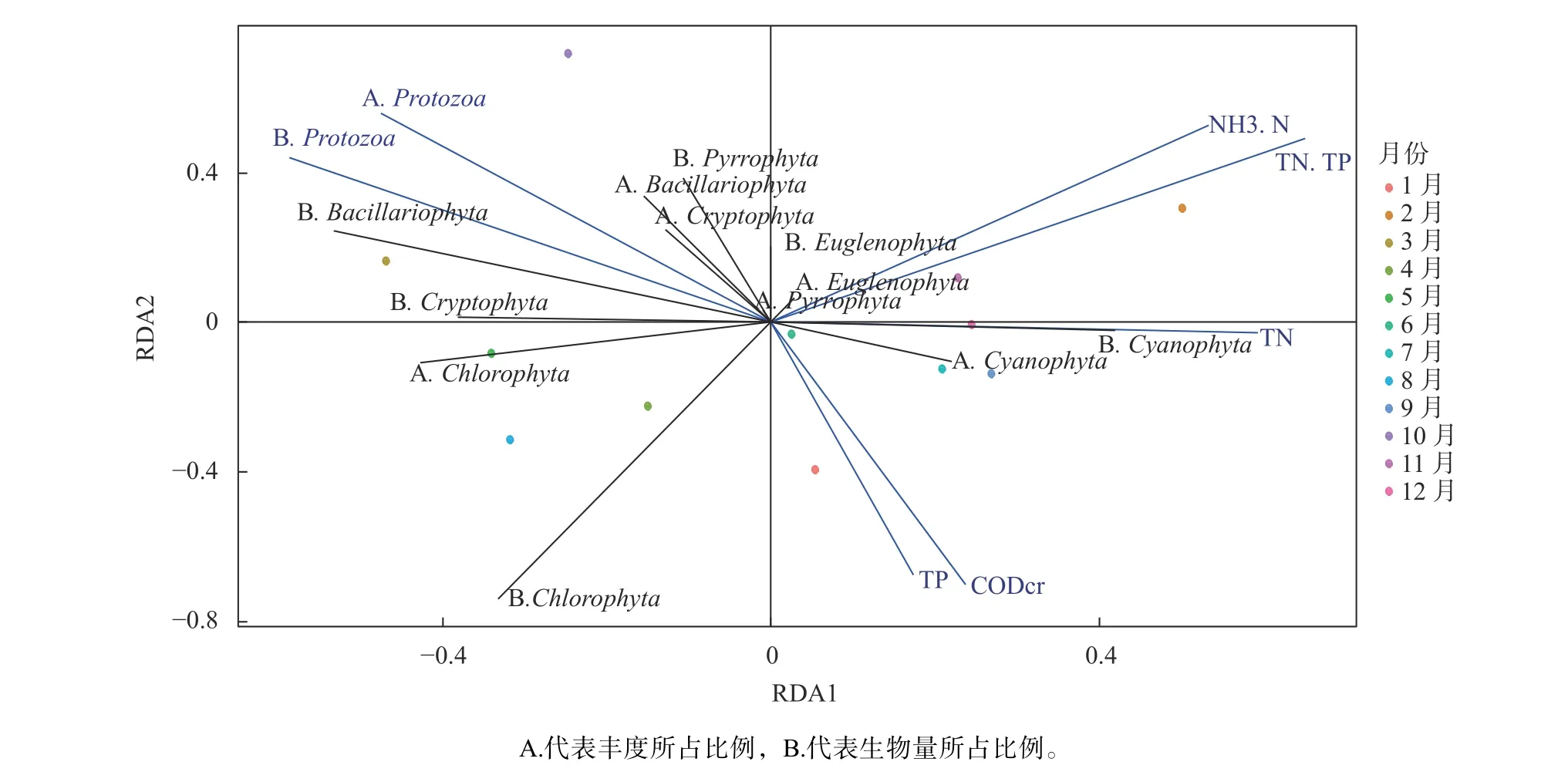
图6 杞麓湖不同月份浮游植物各类群丰度和生物量占比与环境因子之间的相关分析Fig.6 Correlation analysis of the rate of phytoplankton abundance and biomass with environmental factors in Qilu Lake in different months
3 结论与讨论
3.1 杞麓湖浮游植物群落特征及演替
本研究发现杞麓湖浮游植物包括绿藻门、硅藻门、蓝藻门和裸藻门等淡水生态系统较为常见的8个门类,相比于1982—2018年的调查[13,16-17],浮游植物种类组成并未发生明显的变化。该湖浮游植物群落结构与其他九大高原湖泊中的阳宗海、滇池、星云湖和异龙湖的单一型浮游植物群落结构相似,由蓝藻门藻类占绝对优势[16]。在这些高原湖泊中,阳宗海和异龙湖的浮游植物以阿氏浮丝藻和水华束丝藻为主要优势物种[16,31],与本研究结果相似。从1980年至今,与一直以微囊藻为主要优势物种的滇池和星云湖[13,16,32-33]相比,杞麓湖浮游植物的优势物种从1980年和1990年的微囊藻[13],到2016年的史式棒胶藻(Rhabdogloea smithii)、水华束丝藻、阿氏浮丝藻和螺旋藻(Spirulina)[16],再到2017—2018年的阿氏浮丝藻和孟氏浮丝藻(Planktothrix mougeotii)[17],逐渐演替为2020年的阿氏浮丝藻和水华束丝藻等。可能是因为杞麓湖、以及类似变化下的阳宗海和异龙湖其氮磷比远高于滇池和星云湖的氮磷比[16],微囊藻容易在低氮磷比的湖中爆发,而高氮磷比的湖泊更适合水华束丝藻这种固氮型丝状蓝藻的大量繁殖有关,这种趋势与Yamamoto和Nakahara的研究结果[34]也相似。与2017—2018年的结果相比[17],本研究调查得到2020年杞麓湖浮游植物的细胞数量有明显下降。除此之外,相比于更早些的研究[13,16-17],该湖蓝藻细胞的丰度和生物量所占比例也有下降趋势。国内外多位学者证明,鲢对蓝藻有较高的消化率[35-39]。大量实验证明,鲢、鳙等鱼类通过控制难以被浮游动物捕食的大型丝状蓝藻的数量,降低了个体较大的浮游植物的量[40-44]。近十年来,杞麓湖累计投放1000余万尾的滤食性鲢鳙[45],可能在一定程度上控制了湖中占主要优势的大型丝状蓝藻的数量,对湖内浮游植物数量的减少起到了一定的作用。
本研究还发现杞麓湖浮游植物群落结构存在明显的季节性演替。7月份,湖内浮游植物数量达到全年最高值,蓝藻相对丰度达到全年最高水平(92.78%),浮游植物群落结构稳定性和均匀度全年最低,湖区在该时期正处于水华束丝藻爆发阶段。由于在富营养化或者重度富营养化湖泊中,浮游植物通常被几种蓝藻物种占据,形成主要优势物种,因此均匀度也较低[29]。杞麓湖在夏季浮游植物群落结构稳定性较弱且均匀度低,从而更容易导致蓝藻大量繁殖形成水华。
3.2 浮游植物与环境因子的关系
杞麓湖中浮游植物受关键理化因子和浮游动物共同的影响。关键理化因子中,氮、磷是浮游植物生长常见的限制营养元素,可以调节浮游植物的生长,影响浮游植物的生物量和数量[4-47]。本研究发现总氮和总磷是影响该湖浮游植物的主导因素,总氮、总磷以及两者之比对浮游植物总丰度有显著影响。Smith[48]收集17个湖泊的数据,发现了TN/TP规则。当TN/TP质量比超过29时,蓝藻很少,而当TN/TP质量比低于29时,蓝藻占优势[48]。部分学者对浮游植物种群受TN/TP比值的影响持支持观点[49-50]。也有一些持反对观点的,比如Xie等[51]的试验结果与TN/TP规则不符,在高度富营养化的东湖中,低TN/TP值不是微囊藻水华的原因,而是微囊藻水华的结果。本研究中杞麓湖无论是在TN/TP低于29的多数月份,还是在TN/TP高于29的月份,蓝藻始终占主要优势。因此,本研究结果同样不符合TN/TP规则。尽管发现湖中优势类群蓝藻的生长受到一定程度氮的限制,但是结果显示造成不同月份间浮游植物群落的差异显著贡献最高的两种蓝藻,同时又是主要优势物种的阿氏浮丝藻和水华束丝藻均未与氮磷有相关性,而是分别受到化学需氧量和原生动物的影响,并且这两种主要优势丝状蓝藻之间的相互作用机制尚不明确。因此,有关杞麓湖水华束丝藻和阿氏浮丝藻之间的相互作用等问题,值得进一步研究。
致谢:感谢中国科学院昆明动物研究所杨君兴研究员对论文写作的指导!感谢中国科学院昆明动物研究所何宇娇、西南林业大学杜小兵和郭荣、集美大学耿华礼和李垣甫在数据收集时的帮助!
