细胞自噬分子调控机制的研究进展
2022-12-20福建省农业科学院畜牧兽医研究所福州350013
王 劭 福建省农业科学院畜牧兽医研究所 福州 350013
自噬过程于1957年由克拉克首次使用电子显微镜描述,但“自噬”一词于1963年由因在溶酶体方面的工作获得诺贝尔奖的Christian de Duve首次引入。自噬(auto-self,phagy-eating)被定义为一种进化上保守且受严格调控的溶酶体途径,可降解细胞质物质和细胞器,甚至降解微生物入侵者[1-2]。自噬过程具有不同的清除细胞质成分的机制和途径,因此,可以更准确地细分为3种类型:巨自噬、微自噬和分子伴侣介导的自噬[3-5]。从酵母到人类,微自噬和巨自噬都是高度进化保守的,而分子伴侣介导的自噬是一种选择性自噬形式,仅在哺乳动物细胞中观察到[6-7]。巨自噬代表了最广泛研究的自噬形式,并且涉及称为自噬体的双膜囊泡的显著形成,其在与溶酶体融合之前围绕并随机隔离细胞质成分[8-10]。一般来说,自噬被认为是一种非选择性的促生存细胞反应[11]。然而,几种表现出选择性的自噬形式,例如mitophagy(降解线粒体)、reticulophagy(降解内质网)、xenophagy(降解微生物)和aggrephagy(降解聚集的蛋白质)已被认可[12-13]。
1 自噬的调控与信号转导
自噬的原始科学定义(希腊语,“自食”)是将细胞质货物运送到溶酶体进行降解[14-15]。至少存在三种不同形式的自噬(巨自噬、微自噬和伴侣介导的自噬),它们在向溶酶体输送货物的方式方面有所不同。巨自噬是真核细胞用于维持营养稳态和细胞器质量控制的主要分解代谢机制,它由一组进化上保守的基因介导,即自噬相关基因(ATG),最初是在酵母基因筛选中发现的[16]。除了少数例外,所有ATG基因都是有效形成密封自噬体所必需的,这些自噬体继续与溶酶体融合[17-18]。在高等真核生物中,许多ATG基因功能多样化,以促进细胞外货物向溶酶体递送,促进细胞内货物的质膜定位或细胞外释放,并协调细胞内与各种细胞信号通路的通讯。
在哺乳动物细胞中,饥饿诱导的自噬受大约20种核心ATG蛋白的调节[19-20],这些蛋白可分为几个功能单元:(1)ULK激酶核心复合物,包括ULK1/2、ATG13、RB1CC1/FIP200和ATG101;(2)自噬特异性III类磷脂酰肌醇3-激酶(PI3K)复合物,包括VPS34、VPS15、Beclin1和ATG14L;(3)ATG9A运输系统,包括ATG9A、WIPI1/2和ATG2A;(4)ATG12泛素-类似偶联系统,包括ATG12、ATG7、ATG10、ATG5和ATG16L1;(5)LC3类泛素偶联系统,包括LC3A/B/C、ATG7、ATG3和ATG4A/B/C/D。这些ATG蛋白在液泡附近分层募集,并组织自噬体形成所必需的自噬体前结构。在ATG8脂质系统中,活化的I型ATG8(LC3-I)被移动到ATG3,最终与磷脂酰乙醇胺结合[21]。ATG8-磷脂酰乙醇胺化合物通常被称为II型ATG8(LC3-II),是最公认的自噬活性分子标记物。p62在选择性自噬与泛素信号传导之间起作用,与多泛素化蛋白聚集体结合并与LC3-II结合,使吞噬细胞能够吞噬细胞溶质成分、生长并最终关闭自噬体。由于蛋白质p62通常被自噬降解并在自噬受到抑制时积累,因此,p62的减少表明囊泡降解和自噬通量成功[22-23]。
由于鉴定了许多自噬相关基因ATG,以及凭借于监测这一过程的实验方法,自噬得到了广泛的研究[24]。用电子显微镜检测自噬体是最传统的方法。因此,常规形态测量法是目前评估组织和培养细胞中自噬的标准。然而,电子显微镜方法的潜力有限,因此应用了用于识别和量化以及调节自噬活性的新方法,荧光显微镜是检测参与自噬体生物发生的分子的有力方法[25-26]。迄今为止,LC3(一种微管相关蛋白1轻链3)是唯一一种在从吞噬细胞到溶酶体降解的整个过程中特异性定位于自噬结构的特征明确的蛋白质[27-28]。自噬体的数量通常与LC3-II点状结构的数量密切相关,因此通常可以使用荧光显微镜对其进行可视化,并使用图像分析软件进行分析。LC3-II表达也可以通过免疫印迹分析或流式细胞术进行定量和灵敏监测[29]。尽管LC3-II是一种很有前途的自噬体标志物,但有必要仔细解释结果,因为LC3-II水平升高可能与自噬体合成增强或自噬体转换减少有关[30]。自噬是一个动态和连续的细胞内降解过程,用于去除蛋白质或因折叠错误而老化、受损的细胞器。自噬通常是身体对饥饿和其他压力的非选择性反应,令人惊讶的是,在营养丰富的条件下会发生基本自噬或组成性自噬。自噬具有复杂的过程和调控机制,在生理或病理条件下具有重要意义[31-32]。
自噬主要是一种有效的细胞内分解代谢途径,用于降解异常细胞蛋白聚集体和受损细胞器。尽管自噬最初被认为是一种细胞应激反应,但越来越多的证据表明它在多种生物过程中发挥着正常的生理作用[33]。迄今为止,已在生命体大多数器官和许多不同的发育阶段发现自噬,表明它对细胞稳态和更新至关重要[34]。自噬具有抗应激和生理功能,一方面,自噬通过降解细胞内成分在营养剥夺或其他压力下为合成代谢产生新的构件和能量;另一方面,自噬通过在正常条件下降解受损的细胞器或有害成分,参与细胞稳态调节和疾病预防。大量研究表明,在正常生理条件下,自噬发生在各种组织和器官中,并且处于不同的发育阶段。这些结果表明,自噬在正常组织的发育和功能维持中是必不可少的。自噬的生理作用可分为两个方面:在形态发生过程中促进细胞重塑以及在成熟后维持细胞稳态[35-36]。因此,自噬在生物体中发挥着重要的生理功能。
2 自噬体途径
自噬通过自噬体的形成将细胞质成分传递给溶酶体降解[37-39]。自噬的形态学试验包括几个连续的阶段:起始、成核、伸长、闭合、成熟,最后降解和向细胞质输出物质(见图1)。
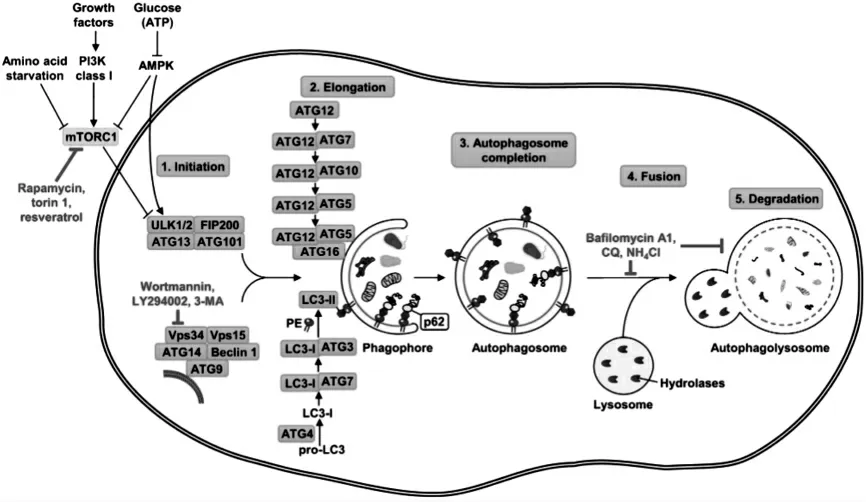
图1 细胞自噬机制及其调控
2.1 自噬诱发 在最初的步骤中,形成了一种杯状的双膜结构,称为“隔离膜”。这种独特的液泡周围结构,称为吞噬细胞,被认为是自噬体的前体,即为一种含有细胞质成分的双膜囊泡[40]。隔离膜的确切来源仍有争议,但可能没有单一来源。粗面内质网、线粒体外膜或高尔基体的无核糖体区域似乎是吞噬细胞的来源[41-43]。
2.2 自噬膜伸长和闭合 吞噬细胞伸长并完全捕获隔离的待降解内容物,形成双膜自噬体,或“自噬泡”。隔离可以是非特异性的,涉及吞噬大量细胞质;也可以是选择性的,针对特定的目标内容物,如细胞器或侵入性微生物[44]。自噬体在细胞质中随机形成,典型的自噬体双膜空泡大小为300~900 nm。这一步是一个简单的隔离,不会发生降解。
2.3 自噬体成熟和降解 完整的自噬体经历了一个逐步成熟的过程,包括自噬体与溶酶体融合,自噬溶酶体内容物被水解酶降解。最近的一些研究揭示了细胞骨架元素(如肌动蛋白微丝和微管)在自噬的特定方面的重要性[45]。自噬体以依赖动力蛋白的方式沿着微管运输到溶酶体,溶酶体聚集在微管组织中心(MTOC,位于细胞核附近)周围。在那里,自噬体与溶酶体对接,自噬体的外膜依次与溶酶体的膜融合[46]。内膜最终与货物一起被驻留的水解酶消耗掉。此外,自噬体可以首先与其他囊泡融合,包括内体和多泡体,形成称为两性体的混合细胞器,随后与溶酶体融合,其中包埋的细胞溶质内容物被降解[47]。目前尚不清楚两性体形成是否是自噬体-溶酶体融合的先决条件。与溶酶体融合的自噬体随后成熟为单膜降解细胞器自溶酶体,通常称为“自噬溶酶体”[48]。水解酶的酸化和获得,包括蛋白酶、脂肪酶、核酸酶和糖苷酶,使自溶酶体能够降解隔离的自噬体内成分,然后将分解产物转移回细胞质,在那里它们可以被回收或进一步分解代谢。自噬泡的流动性依赖于微管,而融合似乎与细胞骨架元素无关。上述液泡的定义是基于它们的功能,而不是它们的形态。因此,区分自噬体、两性体和自溶体并不容易,尽管已经提出了一些监测自噬通量的方法,但降解步骤相当难以测量[49]。自噬体的成熟是一个相对较快的过程,在饥饿条件下,所有描述的事件都发生在10~15 min内[50]。
3 自噬活性的调节
在哺乳动物中,自噬体的形成似乎是一个高度协调的多步骤过程。在自噬体形成的早期阶段,自噬泡的成核需要III类磷酸肌醇3-激酶(PI3K)的活性,PI3K是含有重要蛋白质ATG6(也称为Beclin 1)的大分子特异性复合物的一部分[51]。在哺乳动物中,至少有3种类型的III-PI3激酶复合物有助于自噬,因此,自噬体的形成受到Rab7和UV-RAGVps34-beclin1 PI3激酶复合体的正向调节,而由Rubicon-UV-RAG-Vsp34-beclin1复合物负调控[52-53]。虽然由含有Beclin-1的复合物产生的PI3标记了自噬体生成的位点,但吞噬细胞的延伸和自噬体的成熟需要2个泛素样系统[54]。ATG5和ATG12参与2个类泛素化反应中的第一个,这些反应与ATG16L(ATG16样)非共价相互作用以形成复合物[55-56]。第二个类泛素化反应涉及在哺乳动物中切割微管相关蛋白1轻链3(MAP1-LC3),也称为ATG8或LC3,随后与磷脂、磷脂酰乙醇胺(PE)结合形成LC3-II在自噬体膜中[57]。LC3-II是一种独特的蛋白质,它与新形成的自噬体膜特异性相关,而不与其他囊泡结构相关,LC3-II存在于细胞溶质和自噬体内表面,因此在自噬体形成过程中增加[58]。在哺乳动物中,LC3从LC3-I(游离形式)转化为LC3-II(PE结合形式),被认为是自噬体形成的关键步骤[59]。尽管ATG5-ATG12在自噬体形成完成后丢失,但即使在与溶酶体融合后,LC3-II仍与自噬体结合,此后自噬体细胞溶质面上的LC3-II可以脱脂并回收(至LC3-I)。因此,LC3-II的水平与自噬体数量相关,这种特定的关联使其成为监测自噬的极佳标志物[60-61]。此外,自噬体的形成受到细胞内和细胞外氨基酸浓度和ATP水平的严格调节,这些信号通路包括哺乳动物雷帕霉素靶点mTOR激酶[62]。这种进化上保守的TOR激酶复合物1是营养状态的传感器,在控制细胞生长中起重要作用[63]。
4 自噬的生理功能
自噬在所有真核细胞中组成性地以低基础水平发生,它作为细胞质量控制机制发挥作用,负责细胞内成分的正常更新和维持细胞能量平衡[64]。在大多数情况下,自噬构成了促进细胞存活的应激适应途径,因此与许多基本生理活动有关,例如发育、分化和组织重塑[65-66]。自噬产生的分解产物-氨基酸、脂肪酸、核苷和碳水化合物,由溶酶体通透酶释放,可以全身输送并输出到细胞质中以供重复使用。此外,自噬降解中释放的氨基酸可以与脂肪酸一起被三羧酸循环用于维持细胞ATP的产生[67]。自噬可以通过许多压力源迅速上调,包括营养缺乏、激素和生长因子剥夺、高温或缺氧或药物(例如雷帕霉素)[68]。真核生物中最著名的进化保守应激反应是饥饿期间在培养细胞和完整生物体中刺激自噬[69-70]。最近,人们认识到自噬不仅有助于细胞质成分的更新,而且还可以靶向细胞内病原体以在先天免疫过程中降解[71-72]。自噬的这种管家功能包括通过称为异体吞噬的过程直接消除某些细胞内微生物病原体。消除许多细菌(A组链球菌、金黄色葡萄球菌、结核分枝杆菌、冠状立克次体)、寄生虫(弓形虫)和病毒(I型单纯疱疹病毒),自噬参与细胞自主抗菌防御系统[73-75]。
此外,自噬在调节适应性免疫反应、免疫耐受以及免疫系统的发育和稳态方面发挥着关键作用[76]。在神经系统中,自噬的刺激限制了有毒产物的积累并保护神经元免于退化[77]。在肌肉中,心脏细胞的基础自噬似乎在细胞质成分(包括线粒体)的转换中具有特殊的管家作用,并且在应激反应中被上调[78]。Griffiths等研究表明,自噬通过减少氧化应激和炎症以及增加一氧化氮的生物利用度来保护动脉内皮功能。也有人提出自噬参与网织红细胞成熟过程中线粒体的降解[79]。来自肝脏的脂滴通过它们在自溶酶体中的组成性降解被消除[80],但另一方面,自噬是产生白色脂肪组织特征的大脂滴所必需的。自噬的抗衰老作用可能取决于其质量控制功能,该功能通过易于聚集的蛋白质的沉积和受损的线粒体来限制活性氧的产生[81]。越来越多的证据表明,在啮齿类动物和人类中,经典自噬(巨自噬)和伴侣介导的自噬水平随着衰老而下降[82]。
老化的有丝分裂后细胞的一个主要特征是黄褐色自发荧光脂褐质沉积物的积累[83]。自噬的一个关键生理作用似乎是细胞保护,但这一过程也通过其与细胞凋亡和坏死的交织关系与细胞死亡有关[84]。一些研究表明过度降解细胞成分促成了一种细胞死亡形式,通常称为II型程序性细胞死亡或自噬性细胞死亡(ACD),其特征是细胞质部分的大规模隔离,使细胞具有特征性的空泡状外观。ACD也以不依赖于半胱天冬酶的方式发生,没有凋亡形态的关键特征,例如染色质浓缩和核碎片[85-86]。然而,自噬和细胞凋亡有几个共同的调节元件,这表明自噬在细胞存活和细胞死亡中具有双重作用[87]。
5 小结与展望
自发现第一个酵母ATG基因以来,在了解自噬生物学及其与生物体健康和疾病的相关性方面取得了快速进展[88]。重要的是,自噬调节可能是适用于多种疾病的治疗策略。然而,对自噬蛋白在免疫相关过程中多种功能的分子机制的理解仍然相当原始,当前的蛋白质组学和基因组筛选有可能改变自噬和免疫研究。这种转变将包括通过使用现代系统生物学工具来广泛地了解自噬蛋白-蛋白相互作用和信号调控网络,从而促进对现有已知自噬免疫功能分子机制的更深入理解。也许更令人兴奋的是,这种转变可能会揭示这种古老的自卫机制在免疫方面发挥作用的新方法[89]。
近年来,关于经济动物自噬的研究数量显著增多,支持动物生产的生理功能(繁殖、生长、免疫系统和肌肉组织成熟)已被证明受自噬的调节[90]。然而,在农业面临日益严峻的社会和环境挑战背景下,仍然需要做很多工作来了解自噬在畜禽养殖中的作用,以及如何应用所获得的知识来改善动物的发育、生长和健康,以及它们的产品质量。例如,虽然现在在猪、牛、鸭或鸡中明确确定了许多病原微生物使用自噬体机制进行传播,但缺乏有关机制的精确数据,阻碍了有效免疫策略的定义[90-92]。同样,更好地了解自噬对这些物种对环境、营养或氧化应激反应的机制,将有助于定义应对生态农业绿色循环发展的科研策略。
