不同皮色洋葱花青素合成酶基因片段的克隆及苗期表达分析
2022-12-20王振宝徐宏志刘冰江霍雨猛孙亚玲李艳伟吴雄杨妍妍
王振宝,徐宏志,刘冰江,霍雨猛,孙亚玲,李艳伟,吴雄,杨妍妍
(1. 山东省农业科学院蔬菜研究所,山东 济南 250100;2. 东北农业大学园艺园林学院,黑龙江 哈尔滨 150036)
洋葱(Allium cepaL.)又名圆葱、葱头、球葱等,为百合科葱属二年生草本植物。洋葱于近代传入我国,由于具有适应性强、耐贮藏和运输的特点,在我国广泛栽培,是一种重要的出口创汇蔬菜[1]。洋葱以肉质鳞茎为主要食用器官,营养物质丰富,有“蔬菜皇后”的美誉;洋葱中含有的类黄酮、有机硫等还有一定的药用和保健作用,其中类黄酮具有调节血脂、抗氧化、抗肿瘤、抗病毒、降低血栓形成、消炎等作用[2]。
鳞茎颜色是洋葱的一个重要性状,主要有紫、红、黄、白等颜色。类黄酮类化合物是洋葱鳞茎产生不同颜色的主要因素。花青素作为一种重要的类黄酮类天然水溶性色素,广泛分布于植物细胞液中,是植物花、果实和储藏器官等呈现不同颜色的主要色素之一[3]。植物花青素生物合成途径的研究已较为成熟,是以直接前体苯丙氨酸为底物,经过一系列酶的催化,最终形成稳定的花色苷[4]。花青素合成酶(anthocyanidin synthase,ANS)位于花青素生物合成途径的倒数第二步,是花青素生物合成途径末端的限速酶,能催化无色花色素转化为有色花色素,其表达水平的高低会影响植物组织器官中花色苷的累积,从而产生不同的颜色[5]。
目前,已经从花生、黑稻、苹果、东方百合等多种植物中克隆了ANS基因[6-9],而且研究发现ANS基因的表达具有组织特异性和品种特异性,如黑果枸杞果实中LrANS基因的表达量显著高于其他组织,表达量由高到低依次为黑果、紫果、绿果、花、叶、茎、根[10];紫心甘薯块根、茎、叶中ANS基因的表达水平显著高于白心甘薯[11]。但有关洋葱ANS基因的研究相对较少,缪军等[12]2010年克隆了洋葱花青素合成酶基因AcANS,并进行了生物信息学分析;Kim等[13]2016年鉴定出了2 个突变的ANS等位基因,能造成洋葱花青素合成酶失活,从而影响花青素合成。本研究以30 天苗龄的紫皮和黄皮洋葱幼苗为试材克隆ANS基因片段,并对其在两种皮色洋葱幼苗不同组织中的表达特性进行研究,以期为洋葱ANS基因全长序列获得和功能研究奠定基础。
1 材料与方法
1.1 试验材料
供试材料为紫皮洋葱育种系175F 和黄皮洋葱育种系DH17-1 的30 天苗龄幼苗,均定植于山东省农业科学院蔬菜研究所试验基地。于2021年10月13 日分别取幼苗的新鲜根、叶鞘和叶片,迅速放入液氮中冷冻,于-80℃保存备用。
1.2 洋葱总DNA、总RNA 的提取及cDNA 制备
洋葱基因组总DNA 采用快捷型植物基因组DNA 提取试剂盒(Tiangen,北京)提取,具体操作参照说明书。
洋葱总RNA 采用Trizol(Invitrogen 公司)法提取,参照说明书进行操作。取完整性良好的总RNA,利用反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(宝生物工程大连有限公司)反转录合成cDNA。
1.3 PCR 扩增
分别以洋葱基因组DNA 和反转录合成的cDNA 为模板扩增洋葱花青素合成酶基因片段。所用目的基因上游引物AcANS-F 序列为5′-CTAACGATCAATCTAAAGGGA - 3′, 下游引物AcANS-R 序列为5′-GCAGTATGAACGATAAGGCAC-3′,由青岛蔚来生物科技有限公司合成。
PCR 反应总体积为25 μL,其中,2×TaqPCR MasterMix 12.5 μL,模板1 μL,上下游引物各1 μL,用灭菌蒸馏水补齐至25 μL。扩增程序为:94℃预变性5 min;94℃变性30 s,55℃退火45 s,72℃延伸30 s,35 个循环;最后72℃保温10 min,4℃保存。扩增产物经1.2%琼脂糖凝胶电泳分离后在凝胶成像系统上检测,并拍照记录。
PCR 扩增产物送青岛蔚来生物科技有限公司测序。
1.4 序列分析
利用DNAMAN 和Blast(http:/ /www. ncbi.nlm.nih.gov/blast/Blast.cgi)进行序列比对分析。
1.5 基因表达量测定
分别从两种皮色洋葱幼苗的根、叶鞘和叶片中提取RNA,反转录为cDNA。以Actin为内参[14],上游引物AcActin-F 序列为5′-ACACGGCCTGGATAGCAACAT-3′,下游引物AcActin-R序列为5′-AGAGCAGTATTCCCAAGCATT-3′,由青岛蔚来生物科技有限公司合成。PCR 反应体系参照1.3,通过PCR 扩展产物条带的亮度将cDNA 模板调整为基本一致的浓度,扩增程序为:94℃预变性5 min;94℃变性30 s,55℃退火45 s,72℃延伸30 s,30 个循环;最后72℃保温10 min,4℃保存。扩增产物经1.2%琼脂糖凝胶电泳分离后在凝胶成像系统上检测,并拍照记录。然后用基因特异引物AcANS-F 和AcANS-R 进行PCR扩增,扩增程序同上,扩增30 或35 个循环,扩增产物经1.2%琼脂糖凝胶电泳分离后在凝胶成像系统上检测,并拍照记录。
2 结果与分析
2.1 AcANS 基因片段克隆
根据已报道的花青素合成酶基因序列设计引物,分别以紫皮和黄皮洋葱的基因组DNA 和反转录合成的cDNA 为模板进行扩增,均扩增出1 条特异条带(图1)。以基因组DNA 为模板在两种皮色洋葱中均扩增出约510 bp 的目的片段,与预期片段大小相吻合;以反转录合成的cDNA 为模板在两种皮色洋葱中均扩增出约400 bp 的目的片段。分别将目的片段回收测序。
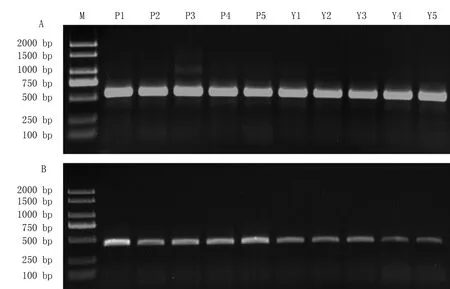
图1 不同皮色洋葱幼苗中AcANS 基因的PCR 扩增结果
2.2 AcANS 基因片段序列比对分析
经测序分析和Blast 比对,克隆到的目的基因片段序列与已报道的洋葱ANS等位基因序列的相似度均在97%以上,确认为AcANS基因片段。利用DNAMAN 软件对从两种皮色洋葱中克隆的AcANS基因片段序列进行对比分析,由图2 可以看出,克隆得到的两种皮色洋葱的AcANS基因组序列长度均为497 bp,碱基序列也完全一致,不存在多态性位点;两种皮色洋葱AcANS基因的编码序列长度均为390 bp,碱基序列也完全一致,且编码区包含一个107 bp、符合GT-AG 规则的内含子。表明克隆得到的两种皮色洋葱AcANS基因完全一致,且无多态性位点。
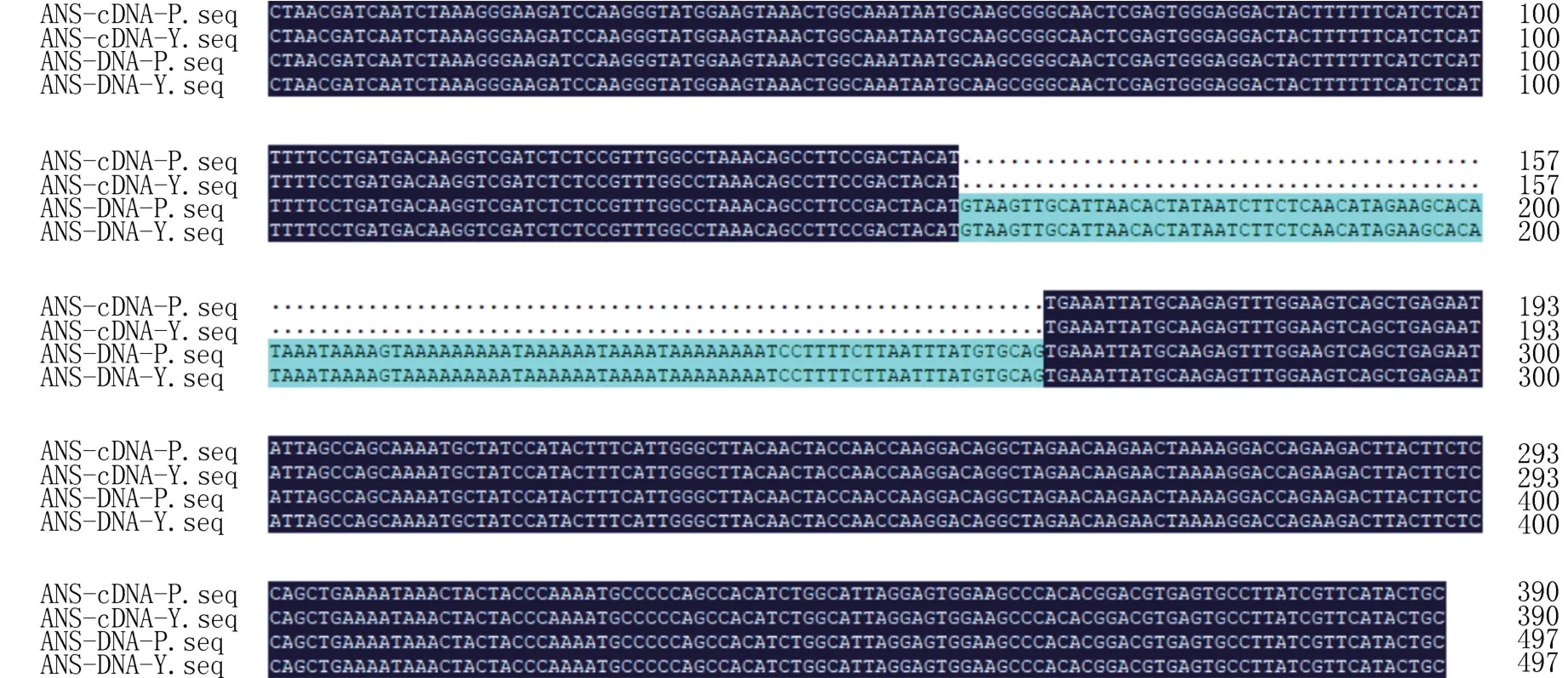
图2 不同皮色洋葱AcANS 基因组序列与编码序列比对结果
2.3 不同皮色洋葱AcANS 基因的组织表达特性
以Actin基因作为内参,采用半定量RT-PCR对紫皮和黄皮洋葱幼苗根、叶鞘、叶片中的AcANS基因表达情况进行分析,结果(图3)显示,两种皮色洋葱的AcANS基因均表现为叶鞘中表达量最高,叶片中次之,根中不表达;在相同条件下,紫皮洋葱叶鞘和叶片中AcANS基因的表达量明显高于黄皮洋葱,且在扩增30 个循环时,紫皮洋葱叶鞘中可以检测到AcANS基因的表达,而黄皮洋葱中未检测到。说明洋葱苗期AcANS基因的表达具有组织特异性,后期颜色越深的部位AcANS基因的表达量越高,而且紫皮洋葱中AcANS基因的表达量更高,这为后期鳞茎颜色不同奠定了基础。

图3 AcANS 基因在两种皮色洋葱幼苗不同组织中的表达情况
3 讨论与结论
花色苷的合成需要PAL、C4H、4CL、CHS、CHI、F3H、DFR、ANS和UFGT等结构基因的参与[4],这些基因的表达或表达量上调都可以造成植物组织器官中花色苷的累积,从而产生颜色上的差异。花青素合成酶(ANS)可以催化无色花青素转化为有色花青素[15]。ANS基因的表达与花青素的累积量呈正相关,颜色越深或越鲜艳的组织部位中ANS基因的表达量越高[15-17]。本研究结果也表明,AcANS基因在洋葱幼苗的叶鞘中表达量最高,其次是叶片中,根中不表达,而且紫皮洋葱中的表达量明显高于黄皮洋葱,这为后期鳞茎产生颜色上的差异奠定了生物学基础。
花青素合成途径相关结构基因的表达受转录因子和外界环境因素的共同调控,转录因子主要包括MYB、bHLH 和WD40 三大类[18]。MYB 转录因子是植物花青素合成途径最广泛的调节因子,也是花青素生物合成中MBW 复合物的主要调控因子,能够正、负调控植物花青素的合成。如在转基因烟草中过表达PsMYB58基因能够显著上调烟草花青素生物合成途径中ANS等结构基因的表达[19];在水仙中过表达NtMYB2基因能够抑制水仙花青素生物合成途径各结构基因的表达[20]。此外,MYB 转录因子还可以与其它转录因子如bHLH、WD40 等共同作用,形成MBW 复合物,调控花青素的合成。本研究分别克隆了紫皮、黄皮洋葱的AcANS基因片段,序列比对结果表明两个片段不论基因组序列还是编码序列都完全一致,也不存在多态性位点,并且AcANS基因在两种皮色洋葱中均能正常转录,但转录水平存在明显差异,在紫皮洋葱中的转录水平明显高于黄皮洋葱,推断这种差异的产生可能是MYB 等转录因子调控的结果,因此,后续有必要克隆洋葱MYB 转录因子,研究其与AcANS基因以及花青素生物合成途径其他结构基因的关系。
洋葱定植前很难通过幼苗准确判断后期葱球的颜色,而实际生产中,在种子或幼苗阶段就能鉴定出皮色对于洋葱种子纯度检测及种质资源鉴定具有重要意义。吴雄等[21]对洋葱花蕾期可育群体和不育群体进行cDNA-SRAP 分析获得了差异表达片段,在此基础上开发了可以鉴定洋葱细胞核育性恢复基因的cDNA 分子标记,该分子标记仅在恢复系花蕾中存在扩增,而其它组织部位和育性材料中均不存在扩增,可以有效鉴定洋葱恢复系材料。鉴于此,本研究利用AcANS基因表达水平的差异,以30 天苗龄紫皮、黄皮洋葱叶鞘cDNA 为模板进行PCR 扩增,扩增30 个循环时,所有紫皮洋葱中均有特异性扩增片段,而黄皮洋葱中无扩增,因此可以作为cDNA 分子标记用于在洋葱30 天苗龄时判断后期葱球的颜色,与传统的田间小区种植鉴定相比所需时间短、用地少、成本低、在苗期即可达到鉴定种子纯度的目的,可应用于洋葱种质资源鉴定和辅助育种等方面。
本研究克隆了不同皮色洋葱AcANS基因部分基因组序列和编码序列,为全长基因序列的克隆奠定了基础;分析了AcANS基因在两种皮色洋葱幼苗各组织中的表达情况,发现其表达具有明显的组织特异性和品种特异性,这为进一步弄清洋葱鳞茎不同皮色发生的分子机制提供了参考。另外发现,可利用不同皮色洋葱叶鞘中AcANS基因表达水平的差异作为cDNA 分子标记,在30 天苗龄时就区分出紫皮和黄皮洋葱。
