脂联素在衰老和损伤中成骨作用的研究进展及主要困惑探讨*
2022-12-17龚燕平
龚燕平
(解放军总医院第二医学中心 内分泌科,北京 100853)
我国正逐渐进入老龄化社会,预计到2050年,我国老年人口将接近3.8亿,占总人口的27.9%[1]。骨折是老年人群常见且危害最大的并发症。骨折使患者活动受限、生活不能自理,增加肺部感染、压疮发生率,不仅严重影响患者生命质量、增加病死率,而且给个人、家庭和社会带来沉重的经济负担。因此,有效促进骨折愈合,降低相关并发症的发生具有重要意义。目前促进骨折愈合的各种生物制剂、无机质及药物价格昂贵,且真正能应用于临床的很少。老年人骨髓脂肪化是其骨质量下降的原因之一,因此脂肪因子脂联素与成骨作用的关系成为目前研究热点。
1 脂联素成骨作用的研究结果不一致,可能与受体后通路不同有关
脂联素是血浆中最常见的脂肪细胞因子之一,是唯一与脂肪含量呈负相关的脂肪因子。全长脂联素蛋白(244 个氨基酸)可以被切割成更小的活性成分[2-3],这些活性成分以球状结构域或不同分子量的形式存在于循环中。既往进行了大量脂联素与糖尿病、心血管疾病、胰岛素抵抗、肝脏代谢、氧化应激、抗炎等关系的研究,并证实其在代谢和抗炎方面具有重要作用[4-10]。近年来研究发现,骨骼肌[11]、骨细胞[12-13]和淋巴细胞[14]也能产生脂联素。骨髓脂肪组织(bone marrow adipose tissue,BMAT)、白色脂肪组织(white adipose tissue,WAT)内的脂肪细胞能产生所有脂联素异构体[15]。每种脂肪组织对骨骼中脂联素水平的贡献尚不清楚,其可能随着年龄和疾病发生相应改变。有研究显示,人类BMAT 与血清总脂联素水平呈正相关[16]。
后续研究证实,成骨细胞和破骨细胞中有脂联素受体1(adiponectin receptor 1,AdipoR1)和脂联素受体2(adiponectin receptor 2,AdipoR2)的表达,并验证了脂联素在骨骼中存在调节作用[17-19]。LENCHIK等[20]在研究脂肪量和骨密度的关系时,首次提出了脂联素与骨骼呈正相关。然而,脂联素对骨形成和骨转换影响的报道却并不一致。各种体外细胞实验证实,脂联素有正向调节成骨、改善骨密度的作用,即随着成骨细胞分化和活性的增加,破骨细胞生成水平降低[17-27]。但在多个基因删除/修改脂联素的小鼠模型实验、人群横断面及RCT 研究中并不能验证脂联素的骨骼效应[28-32](见表1)。
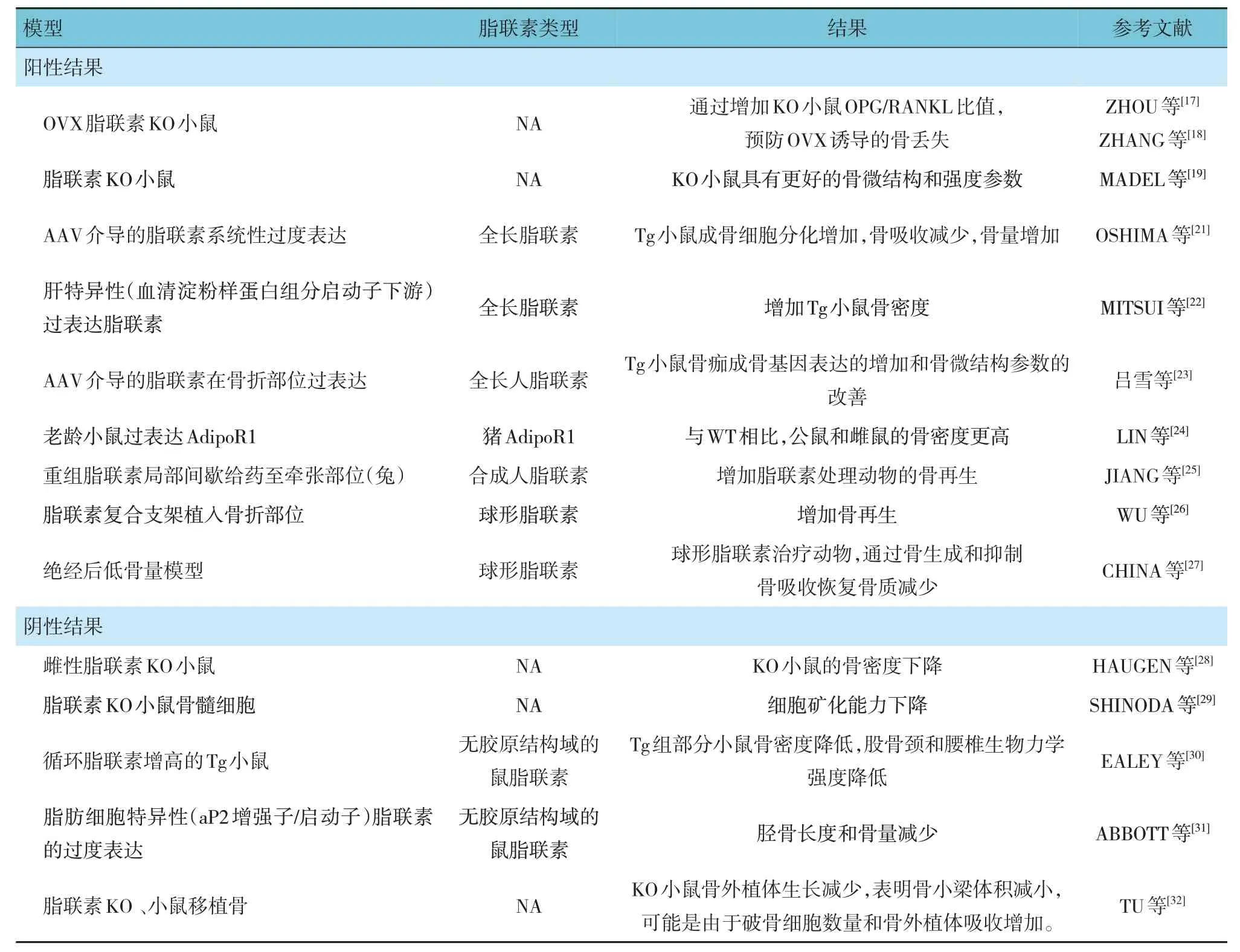
表1 脂联素成骨作用的不同研究结果
脂联素成骨作用的争议限制了药物临床转化,主要原因是全身应用效果欠佳,但其特异作用于骨或骨细胞的效果相对肯定。有研究认为,脂联素对骨骼的局部作用可能存在与中枢神经系统作用相反的效果[23-25]。还有研究认为,脂联素多样性可能是来自其与AdipoR1、AdipoR2 结合的不同,以及结合后激活的通路不同。脂联素信号通路的复杂性常涉及多个直接和间接的受体后机制。因此进一步探索脂联素信号通路,以充分了解促进或抑制脂联素在骨骼调节中的作用是非常必要的[26-30]。
2 脂联素对成骨细胞分化和迁移的作用机制研究
AdipoR1 和AdipoR2 在小鼠成骨细胞、人骨细胞前体骨髓基质细胞(bone marrow stromal cells,BMSC)[33-34]和破骨细胞前体人类外周血单核细胞(peripheral blood mononuclear cell,PBMC)中均有表达。而在所有细胞中,AdipoR1 的表达量均明显高于AdipoR2(可达100 倍)。在多数情况下,AdipoR2甚至低到不能被检测到,比如在MC3T3 小鼠骨细胞前体细胞、健康人类胫骨成骨细胞中均未发现脂联素受体[35]。此外,AdipoR1 对脂联素的球状域有较高亲和力,而AdipoR2 则对所有脂联素具有中等亲和力。因此在同一细胞内与不同蛋白受体结合也很可能产生不同的细胞反应。骨骼AdipoR2 低表达说明骨组织可能主要受球形脂联素影响。
脂联素通过AdipoR1 影响多个通路,包括单磷酸腺苷活化的蛋白激酶(protein kinase,AMPK)、磷脂酰肌醇-3-羟激酶/蛋白激酶B(phosphatidylinositide 3-kinases/protein kinase B,PI3K/PKB)、丝裂原激活蛋白激酶(p38 Mitogen-activated protein kinase,p38 MAPK)、过氧化物酶体增殖物激活受体-γ 共激活物-1α(Peroxisome proliferator activated receptor-γ coactivator-1α,PGC-1α)、信号转导和转录激活蛋白(signal transducer and activator of transcription 3,STAT3)等(见图1)。脂联素可增加成骨细胞骨桥蛋白(Osteopotin,OPN)和碱性磷酸酶(alkaline phosphatase,ALP)的表达。雌激素受体(estrogen receptor,ERα)倾向于激活脂联素受体信号,触发MAPK通路,ERα和特异蛋白1(specific protin 1,SP1)磷酸化,导致下流信号改变。笔者前期研究也表明,局部应用脂联素可通过旁分泌作用增加AdipoR1 数量,上调骨形成蛋白-2(bone morphogenetic protein,BMP-2),促进骨折修复[12]。大多数体外骨细胞模型研究结果表明,骨肉瘤细胞系AdipoR1 表达升高可调节骨细胞分化,但在小鼠成骨细胞系(MC3T3)中并未发现相应的反应[12,34]。在小鼠胚胎成纤维细胞(C3H10T1/2)中,siRNA 抑制AdipoR1 的表达,并显著降低了脂联素诱导的体外成骨细胞分化[36]。在5 周龄的脂联素KO 鼠中,BMSCs 转化成骨细胞的相关基因表达降低,例如促进溶酶特异性组蛋白去甲基化基因(KDM4B、KDM6B)、过氧化物酶体增生剂激活受体(peroxisome proliferator-activated receptor-g,PPARg)基因,从而使BMSCs 的分化由成骨转化为成脂[37]。
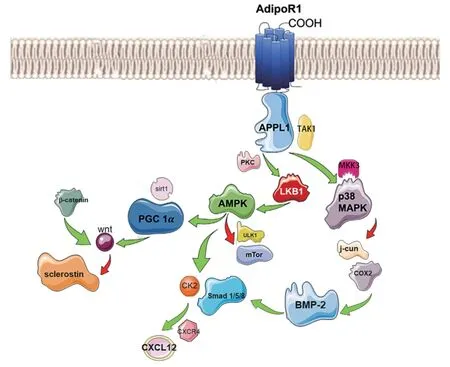
图1 AdipoR1受体后通路图
高水平的CXCL12 能够吸引表达CXCR4 的细胞迁移。系统性球状脂联素输注提高了血清CXCL12水平,促进nestin+BMSC 从骨髓进入外周血。生物活性脂质-鞘氨醇-1-磷酸(sphingosine-1-phosphate,S1P)也可影响成骨细胞和破骨细胞前体迁移[38-39]。S1P 通过鞘氨醇激酶磷酸化鞘氨醇在细胞内合成,在脂联素过表达的转基因小鼠中可以观察到循环S1P 升高。然而目前关于脂联素对SIP 调控迁移的机制知之甚少。
3 脂联素在老年人中的作用与年轻人不同
老年人群的脂联素研究结果与青年人不一致。在56 周龄AdipoR1 过表达的转基因小鼠中,可以观察到骨密度(bone mineral density,BMD)更高,以及抗酒石酸酸性磷酸酶5b(tartrated resistant acid phosphatse,Trap5b)阳性破骨细胞数减少,但在8 周龄或32 周龄的幼龄鼠中却没有观察到[40]。KAJIMURA等[41]的研究表明,6 周龄脂联素敲除小鼠的骨量和成骨细胞数量增加,表明脂联素能抑制幼龄小鼠骨量积累;然而在12 周龄脂联素敲除小鼠骨中,成骨细胞分化标志物runt 相关转录因子(runtrelated transcription factor-2,runx2)和锌指转录因子的基因表达并没有明显变化,脂联素不能再继续限制骨形成。同样地,AdipoR1 缺乏的30 周龄小鼠骨小梁体积、厚度、数量和间距减少,同时成骨细胞数量减少;而在4 周龄幼鼠中AdipoR1 的缺失未影响骨细胞数量。有研究者认为,脂联素信号通路通过AdipoR1 维持成骨细胞的生存和活性至关重要,并且随着年龄增长也能调节破骨细胞的凋亡,而成骨与破骨之间的平衡影响着骨量变化[42]。
4 目前脂联素成骨作用研究的主要困惑
目前对脂联素的研究仍存在许多问题,在许多领域有待进一步证实。①研究主要集中在血浆脂联素水平与骨的关系,对骨局部组织的研究相对较少;②鲜有研究对细胞的矿化能力与脂联素的关系进行评估;③体外细胞研究中有很多未去除培养液中血浆和蛋白的影响;④脂联素T 钙黏蛋白(Tcadherin)受体潜在信号机制还未得到充分研究;⑤仍需要充分了解这些相互作用及其功能后果,以及脂联素调控成骨细胞和破骨细胞迁移的其他潜在分子机制;⑥所有研究几乎集中在青少年或成人,而探讨脂联素在胎儿和新生儿骨骼发育中作用的研究非常有限;⑦脂联素在老龄化中的作用仍有待进一步探索;⑧AdipoR1 敲除的模型研究相对较少,明确定位骨敲除的小鼠模型更少。AdipoR1 受体后作用在全身和局部应用中存在哪些差异,从而导致作用的差异,目前并不明确;⑨对脂联素的人群研究,仍需要更详细的纵向研究,混杂因素如BMI等需要仔细匹配,以充分了解循环脂联素水平与不同年龄组骨密度和骨折率的相互作用。
5 总结
既往研究中脂联素作用效果不明确是限制脂联素临床应用转化的主要原因。探究脂联素对骨骼生物学的调控作用有助于药物开发和非药物(生活方式)干预方式的研究。针对脂联素信号进行干预,可能有助于维护衰老过程中的骨健康和损伤后的骨骼修复。对脂联素骨骼作用的研究,在破除脂联素临床应用困境的同时,也为骨折愈合的治疗提供了新的思路和方向。
