芝麻种间杂交亲和性差异及杂种生物学特征分析
2022-12-14琚铭苗红梅黄盈盈马琴王慧丽王翠英段迎辉韩秀花张海洋
琚铭,苗红梅,黄盈盈,马琴,王慧丽,王翠英,段迎辉,韩秀花,张海洋
芝麻种间杂交亲和性差异及杂种生物学特征分析
琚铭,苗红梅,黄盈盈,马琴,王慧丽,王翠英,段迎辉,韩秀花,张海洋
河南省农业科学院芝麻研究中心/河南省特色油料基因组学重点实验室/神农种业实验室,郑州 450002
【目的】探索芝麻不同种之间的杂交亲和性,分析其杂种的生物学特征,为芝麻野生种种质资源高效利用提供依据。【方法】以芝麻栽培种豫芝11号(,2n=26)和(2n=32)、(2n=32)、(2n=32)、(2n=64)等4个野生种为亲本材料,采用双列杂交方法,通过田间人工授粉配置不同种间组合;结合胚拯救方法获得种间杂种F1。根据杂交结蒴率比较组合杂交亲和性;在盛花期和成熟期观察杂种植物学性状特征,利用Alexander染色法进行花粉粒育性鉴定。通过根尖细胞染色体涂片明确杂种染色体数目及特征。选用自主筛选的胡麻属特异多态性SSR引物,分析种间杂种分子标记差异。【结果】配置了5个芝麻种间的20个正、反交组合,共授粉2 091朵花,获得杂交蒴果370个。发现以染色体数目多的种为母本更易获得远缘杂交蒴果。5个芝麻种之间杂交亲和性的变化范围为1.18%(×)—63.33%(×)。共有9个杂交组合获得杂种F1种子,F1植株的花粉败育率为35.21%—100.00%,其中,与杂交组合F1的可育株比例最高,为87.68%。杂种F1在株高、株型等性状方面均表现出明显的超亲优势。栽培种与各野生种的正反交杂种F1在叶型、花型和花色表现出双亲的局部特征。栽培种芝麻(n=13)与具有n=16染色体组型的3个野生种的杂交亲和性依次为>>;野生种(n=32)与n=16染色体组的3个野生种的亲和性依次为>>在5个种中,野生种与的亲缘关系相对最近。获得的部分杂种植株根尖细胞染色体数目观察显示,杂种的染色体数目与理论值一致。利用3对多态性SSR引物对F1植株的分子鉴定结果显示,真杂种比例为99.66%。杂种染色体核型和特异SSR标记结果显示出胡麻属不同种的遗传特征差异。【结论】胡麻属5个种之间的杂交亲和性差异显著,种间杂交后代杂种优势明显;与的亲缘关系相对最近,可用于芝麻优异种质创制和远缘杂交育种研究;其他种间杂交存在着生殖隔离障碍,可采用胚拯救、分子标记利用等手段加强芝麻野生资源利用。
芝麻;野生种;种间杂交;生物学特征;分子鉴定
0 引言
【研究意义】芝麻(L.)属胡麻科胡麻属,是古老的油料作物,距今已有5 000多年的栽培历史[1]。种植分布广泛。芝麻籽粒富含丰富的不饱和脂肪酸和抗氧化物质,油用和药用价值极高,被誉为“油料皇后”[1-2]。中国是世界五大芝麻主产国之一,因单产水平高、品质优良,在世界芝麻生产、加工和贸易中占据极为突出的地位。但是,受物种特性和主产区气候影响,品种抗病抗逆性差,年份间产量波动较大,对芝麻生产造成重要影响[3]。选育高产、抗病、抗逆新品种是芝麻育种专家长期的主攻目标。【前人研究进展】胡麻属共包含37个种,涉及3种染色体组类型,即2n=26(如,栽培种和野生种),2n=32(如,野生种和)和2n=64(如,野生种和)[4-7]。现有芝麻品种均来自胡麻属唯一的一个栽培种;遗传基础相对狭窄,抗病(如枯萎病、茎点枯病等)、抗逆(如渍害、低温等)等优异种质极其匮乏[8-9]。采用现有种质资源和育种材料及传统育种技术很难提升芝麻品种的丰产和抗病抗逆性[10-11]。检测显示,胡麻属中部分野生种具有高抗病、抗虫、耐渍、耐旱等特性[12-14]。苗红梅等[8]采用强致病力尖孢镰刀菌株室内抗枯萎病精准鉴定技术,开展了多份芝麻种质以及野生种和的枯萎病抗性鉴定。结果显示,仅和对强致病力菌株表现为免疫(DI=0);所有栽培种材料均表现为高感(DI= 100%)。为拓宽芝麻栽培种的遗传基础,加快芝麻抗病和育种研究进程,目前迫切需要深入开展芝麻远缘杂交与野生种利用研究[14-15]。芝麻野生种特征特性调查和种间杂交研究最早起始于20世纪30年代[16-18]。大量研究结果显示,因染色体组特征差异过大,芝麻栽培种与大多野生种杂交均高度不亲和[15, 17, 19],获得的少量种间杂种多表现为花粉败育特征,难以直接利用[20-21]。Tarihal等[22]研究芝麻栽培种和、、等3个野生种之间的亲和障碍,发现以栽培种为父本,可获得少量的杂交种子,但是种胚往往发育异常,没有发芽能力。Kumari等[23]研究芝麻不同种间的杂交亲和性,发现栽培种与野生种和的正反交组合均不能获得杂交种子,栽培种与配置组合可以获得杂交种子,且籽粒饱满。张海洋等[15]选用28份芝麻栽培种材料和2个芝麻野生种(、,2n=64)进行了种间杂种的生物学特性分析。结果显示,与栽培种的杂交亲和率高于。以野生种为母本,与栽培种杂交,杂交种F1在株高、根系结构等性状方面超亲表现明显,株系表现出高抗枯萎病,部分F1株系有低自交结实性。Rajeswari等[24]通过胚拯救获得了(2n=26)和栽培种的杂种。杂种对变叶病的抗性为中等抗性,介于双亲之间。赵瑞红等[25]分析了的染色体核型和rDNA分布,揭示了胡麻属不同种的遗传多样性。为加快芝麻野生种质资源利用提供了科学依据。【本研究切入点】当前芝麻野生种质资源研究不够深入,种间杂交亲和性差,导致远缘杂交育种研究进展缓慢[26-27];大多数野生种与栽培种在染色体组和基因组结构特征等方面差异大,不同种之间的亲缘关系较远,不利于开展远缘杂交育种[14, 17, 24, 27]。【拟解决的关键问题】本研究拟选用芝麻栽培种和4个野生种为试验材料,通过人工授粉和胚拯救技术,研究胡麻属5个种之间的杂交亲和性,明确杂交亲和率、种间杂种表型及分子标记特征,为创制高抗病抗逆芝麻新种质、加快远缘杂交育种进程奠定理论和材料基础。
1 材料与方法
1.1 材料
试验选用中国主推芝麻品种豫芝11号和4个野生种、、和(表1)。上述种质材料均保存在河南省农业科学院芝麻研究中心种质库。

表1 芝麻远缘杂交用亲本信息
1.2 种间杂交配制和杂种的生物学特征观察
2015—2018年,在海南三亚(108°56′E,18°09′N)试验基地和河南原阳(113°93′E,35°07′N)基地进行杂交组合配置、杂交后代种植及性状调查。
种间杂交组合配置:利用双列杂交法组配杂交组合,采用常规技术进行田间管理。参照石淑稳[28]方法,于盛花期进行人工杂交授粉,记录授粉花朵数。
杂交后代培养与杂交结蒴率调查:在授粉后第10—12天,摘取部分杂交蒴果。在超净工作台上,剥取幼胚,进行组织培养[28-29],调查胚拯救成苗率。田间调查杂交蒴果的生长发育状况及结蒴率。
成苗率(seedlings rate,%)=获得的株系数/培养的幼胚数×100%
杂交结蒴率(hybrid capsule formation rate,%)=杂交蒴果数/授粉花朵数×100%
亲本及杂种F1生物学性状调查:分别在盛花期和成熟期调查杂交亲本及杂种F1株系的重要生物学性状,分别为株型、株高、花色、每叶腋花朵数、单株蒴数、蒴果棱数、单蒴粒数及种皮颜色等8个指标。
1.3 染色体核型观察
选用胚培养植株幼嫩根尖4—5 mm,参照赵瑞红等[25]染色体涂片法,制备根尖细胞染色体切片,DFC2500(莱卡,德国)10×及40×镜下观察计数;DM6000B显微镜(莱卡,德国)100×油镜下进行染色体拍照。
1.4 杂交后代花粉粒育性观察
在盛花期,随机选取亲本及F1植株即将开放的花朵。采用Alexander染色法进行花粉粒染色[30],200×显微镜下观察成熟花粉粒形态及颜色。每份材料取5个花朵;观察统计5个视野下的可育和败育花粉数量,计算花粉败育率。花粉败育率(pollen sterility rate,%)=观察到的不育花粉粒数/观察到的所有花粉粒数×100%。
1.5 杂交后代SSR分子标记鉴定
采集亲本及种间杂种单株幼嫩叶片。采用CTAB法提取叶片DNA。选用3对SSR引物进行PCR检测[31]。SSR引物(表2)由华大基因公司合成。PCR反应体系为50 ng DNA模板、10×Buffer 1 μL、10 mmol·L-1dNTPs 0.2 μL、正反向引物(10 μg·L-1)引物各1 μL、2.5 U·μL-1Taq酶0.5 μL,加入无菌超纯水至终体积为10 μL。PCR扩增程序为94℃ 5 min;94℃ l min,58℃ 50 s,72℃ 60 s,32个循环;72℃ 10 min,4℃保存。非变性聚丙烯酰胺凝胶电泳检测PCR产物。
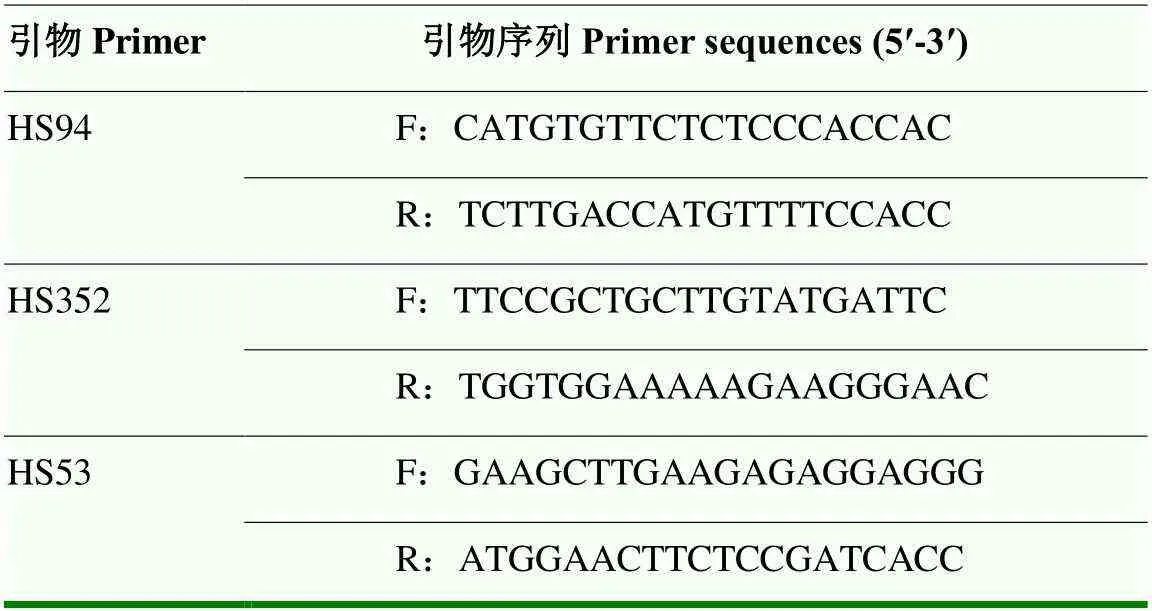
表2 引物信息
2 结果
2.1 种间杂交组合亲和性分析与比较
为分析胡麻属不同种之间的亲和性,选用芝麻栽培种(豫芝11号)和4个重要的野生种、、和,共配置正、反交组合20个(表3)。田间人工授粉共2 091朵花。统计显示,在自然条件下,20个组合共获得杂交蒴果370个,结蒴率范围为1.18%(×)—63.33%(×)。在栽培种与4个野生种的正反交组合中,(G02)×(豫芝11号)的结蒴率最高(41.11%);在野生种之间的双列杂交组合中,(Ken8)和(G01)的正反交组合(H7a和H7b)杂交结蒴率最高,分别为63.33%和48.00%。蒴果成熟后,共收获杂交种子3 151粒。其中,H4b种子无发芽力;H7a和H7b种子有胚,发芽率分别为15.44%和24.75%。
同时,试验采用幼胚拯救技术对杂交种进行组织培养(表3)。共有H3b、H4b、H4a、H6b、H8a及H8b等6个杂交组合获得后代组培苗,并移栽成活。成苗率为2%—93%。其中,Ken8与G01的正反交组合分别获得98棵和66棵组培苗,杂种成苗率显著高于其他组合。表明二者杂交的亲和性最好。
综合上述,5个种间的田间人工杂交和杂种胚拯救统计结果,比较确定,栽培种豫芝11号与4个野生种的杂交亲和性分别为G02>G01>Ken8>Ken1。4个野生种之间杂交结果显示,G01(2n=32)与Ken8(2n=32)组合的结蒴率和种子发芽率均最高,表明二者的亲和性最高,亲缘关系最近。其次是G02(2n=64)与G01(2n=32),结蒴率为34.12%。但萌发试验显示,杂种没有发芽率。
2.2 种间杂交后代生物学特征及育性分析
通过对获得的9个种间杂交组合F1进行种植,分别在盛花期和成熟期观察单株植物学性状特征(图1、图2和表4)。同时对杂种F1的花粉育性进行染色观察(图3和表4)。
调查显示,栽培种与各野生种杂交组合的正反交杂种F1在叶型,花型和花色表现出双亲的局部特征。如,豫芝11号()和Ken1()正、反交后代F1(H1a和H1b)在茎、叶、花及蒴果外形等方面,与亲本Ken1类似(图1-b、图1-f、图2-b和图2-f);但是叶片变小,茎秆变细,与亲本豫芝11号较为相似(图1-a)。F1表现出强分支性,枝条较亲本Ken1柔软。花粉粒育性鉴定结果显示,F1不育(图3-a);蒴果内形成3—4粒种子,但种胚干瘪、败育。
Ken8()×豫芝11号()杂种F1(H2b)的叶片较两亲本变大,分支增多;花冠颜色与Ken8类似,且蒴果表皮茸毛量较多(图1-g和图2-g);花粉粒染色发现个别植株的花粉可育(图3-b);绝大部分蒴果发育至0.8—1 cm时停止生长,种子内种胚干瘪,无发芽力。
G02()×豫芝11号()杂种F1(H4b)的株高比两亲本高,F1植株的形态偏向G02,分支增多;花冠颜色为淡粉色(图1-h);蒴果形态类似G02,蒴果长度发育至约1.3 cm时不再生长,成熟后里面有1—3粒种子且无种胚(图2-e和图2-h)。
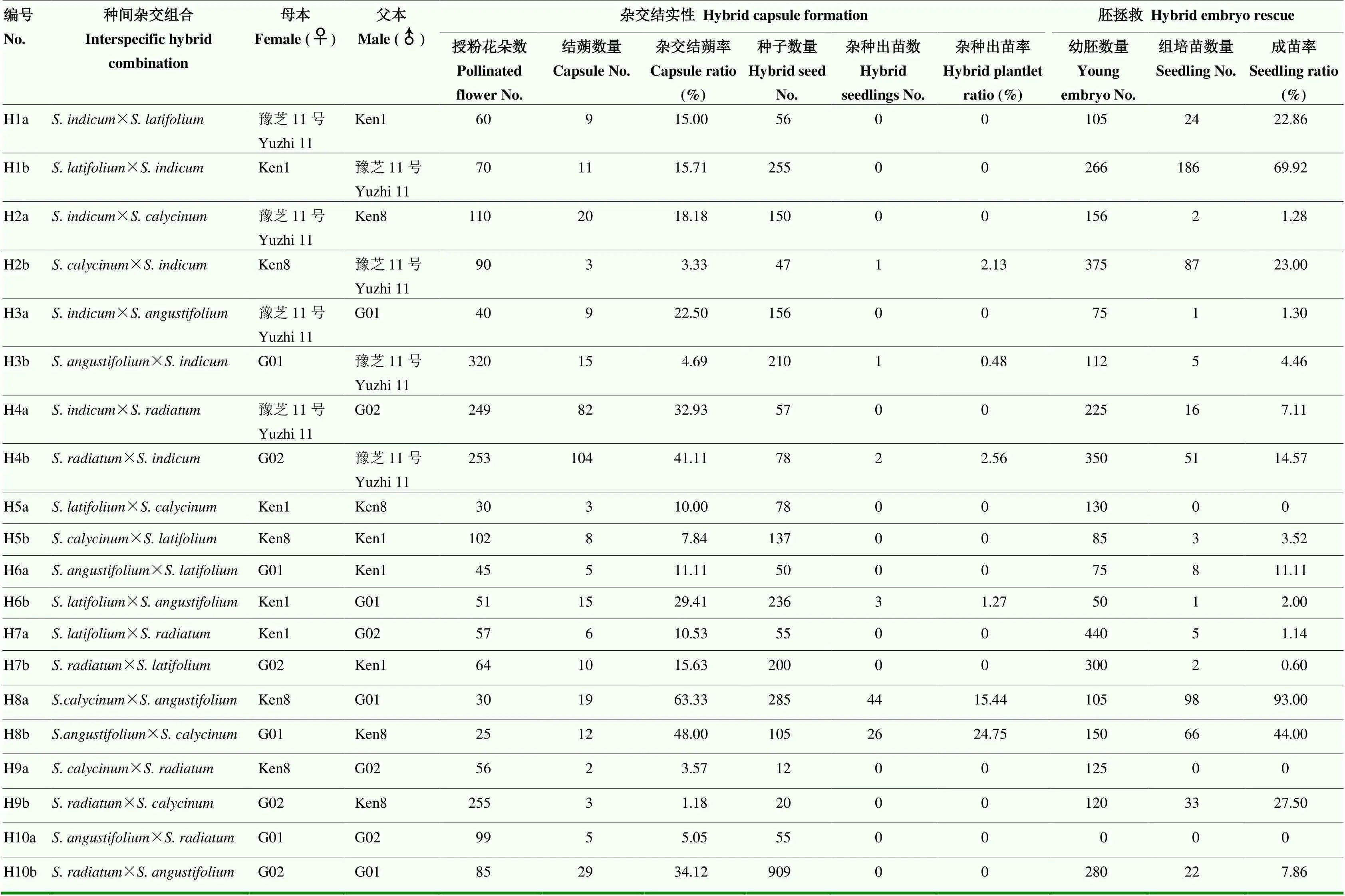
表3 种间杂交组合结实性和杂种胚拯救出苗率统计
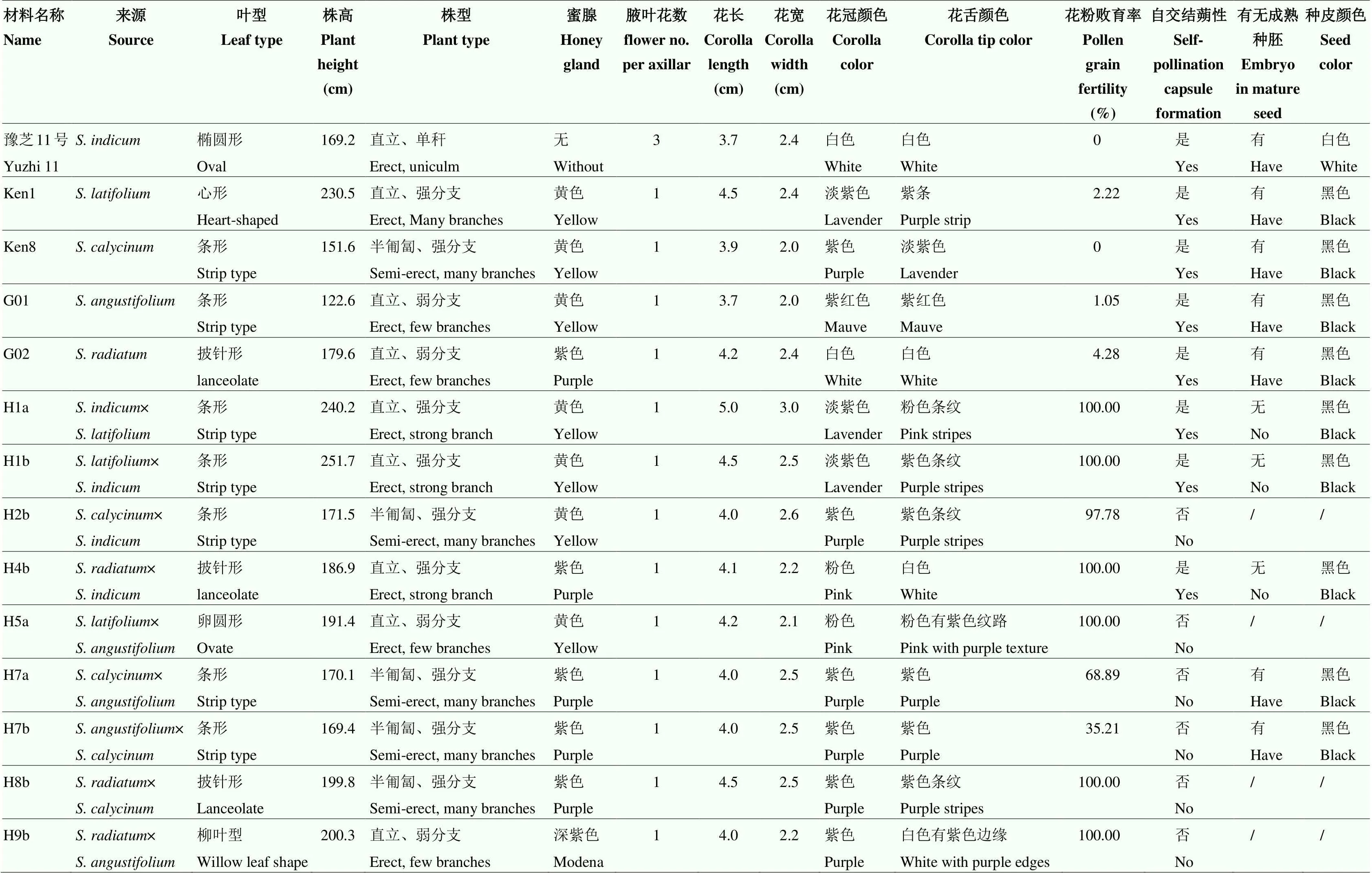
表4 芝麻不同种及种间杂交F1植株的生物学特征统计
/:F1种子无种胚或未结种子 /: represents F1seed without embryo or no seed

a:豫芝11号;b:Ken1;c:Ken8;d:G01;e:G02;f:Ken1×豫芝11号组合F1;g:Ken8×豫芝11号组合F1;h:G02×豫芝11号组合F1;i:Ken1×G01组合F1;j:Ken8×G01组合F1;k:G02×Ken8组合F1;l:G02×G01组合F1
Ken1()×G01()杂种F1(H5a)在株型、花形态方面均与亲本Ken1类似;花冠颜色较Ken1深(图1-i)。随着植株生长,茎秆紫色程度加深。二者杂种F1不能形成可育花粉粒(图3-c);植株结蒴性差,蒴果小,不能形成种子(图2-i)。
G02()与G01()及Ken8的杂种F1分别为H8b和H9b。F1在植株形态方面较为类似。H9b较H8b的茎秆和花冠颜色更深,紫色重(图1-k和图1-l)。H8b、H9b在花朵形态和大小方面与亲本G02的均较为相似。H8b不能形成可育花粉粒(图3-f),结蒴性差;蒴果内无种子形成(图2-l)。H9b可形成较少的可育花粉粒(图3-e);蒴果自然膨大,与亲本G01类似(图2-k);可形成有胚种子。
Ken8()与G01()正、反交组合的杂种F1在植株形态方面较为类似。植株株型介于半匍匐(亲本Ken8)至直立(亲本G01)中间状态;强分支,茎秆及花冠颜色与亲本G01相似(图1-j);花粉粒可育(图3-d);蒴果形态与亲本Ken8类似(图2-j),可形成正常F1种子。随后,对Ken8()×G01()F2群体的414份单株进行育性等性状调查和比较(具体数据未显示)。结果表明,该组合F2群体中,可育株占比达到87.68%,可以直接用于芝麻优异种质创制和远缘杂交育种研究。

a:豫芝11号;b:Ken1;c:Ken8;d:G01;e:G02;f:Ken1×豫芝11号组合F1;g:Ken8×豫芝11号组合F1;h:G02×豫芝11号组合F1;i:Ken1×G01组合F1;j:Ken8×G01组合F1;k:G02×Ken8组合F1;l:G02×G01组合F1
2.3 不同种种间杂交后代染色体数目鉴定
此外,对获得的部分种间杂交组合的后代植株进行了根尖细胞染色体涂片和染色体数目观察(图4)。统计显示,栽培种豫芝11号()×Ken1()(图4-a)和Ken8()×豫芝11号()(图4-b)2个组合的F1植株,其根尖细胞的染色体数目均为29。表明栽培种(2n=26)与野生种Ken1或Ken8(2n=32)杂交后代F1的29条染色体应分别来自双亲的n=13和n=16。
对于野生种之间形成杂种后代的杂交组合,统计显示,其F1染色体数目与亲本有一定变化。其中,G02()与G01()和Ken8()的F1根尖细胞的染色体均为48条(图4-c和图4-d),应该分别来自双亲的n=16和n=32。染色体数目变化证明上述种间杂交组合的后代均为真杂种。
2.4 不同种种间杂交后代分子标记鉴定
为进一步明确种间杂交后代的遗传组成,选用自主筛选的芝麻特异的3对多态性SSR引物,利用上述引物针对8个杂种587个F1植株进行了分子鉴定(图5和表5)。多态性扩增DNA条带的范围为100—250 bp。从图5可以看出,引物对HS352可以区分Ken1、豫芝11号和Ken8及其Ken1×豫芝11号、Ken8×豫芝11杂种F1。HS94可以鉴定Ken8和G01及其后代(图5-a、图5-b和图5-d)。引物HS53可区分Ken1和G01亲本及杂种(图5-c)。HS94可以区分G02和Ken8及杂种(图5-e)以及G02和G01及杂种(图5-f)。结果显示,除从Ken1与豫芝11号的杂种材料中检测出2株假杂种外,其余585份样本均为真杂种,占比为99.66%。表明芝麻不同种种间杂交结果真实可靠。此外,在部分杂种材料中,除了能够扩增出双亲的互补条带外,还出现了新的DNA条带,显示远缘杂交后代材料的遗传多样性。

a:Ken1×豫芝11号组合F1;b:Ken8×豫芝11号组合F1;c:Ken1×G01组合F1;d:Ken8×G01组合F1;e:G02×Ken8组合F1;f:G02×G01组合F1。红色和黄色花粉粒分别为可育和不育型,不同组合F1花粉育性程度不一

表5 芝麻不同种杂交后代SSR分子标记鉴定统计

a:H1b(Ken1×豫芝11号);b:H2b(Ken8×豫芝11号);c:H9b(G02×G01);d:H8b(G02×Ken8)
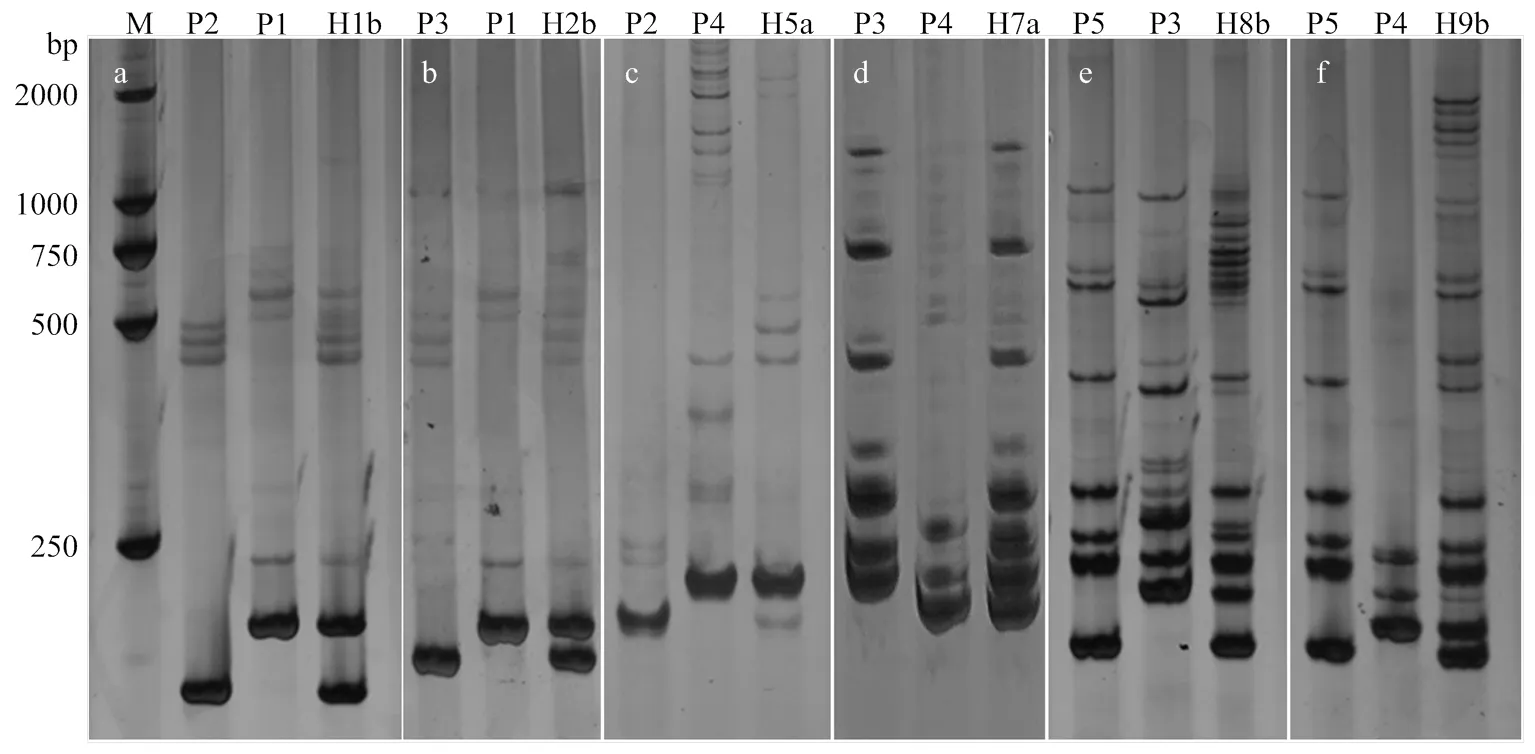
a:引物HS352扩增Ken1和豫芝11号及其杂种F1(H1b);b:引物HS352扩增Ken8和豫芝11号及其杂种F1(H2b);c:引物HS53扩增Ken1和G01及其杂种F1(H5a);d:引物HS94扩增G01和Ken8及其杂种F1(H7a);e:引物HS94扩增G02和Ken8及其杂种F1(H8b);f:引物HS94扩增G02和G01及其杂种F1(H9b)。M:Maker DL 2000;P1:豫芝11号;P2:Ken1;P3:Ken8;P4:G01;P5:G01
3 讨论
本研究选用芝麻栽培种豫芝11号及(2n=32)、(2n=32)、(2n=32)、(2n=64)等4个代表性野生种,系统开展了芝麻不同种间杂交亲合性、杂种后代生物学特征及分子标记特征比较与分析,获得一批远缘杂交材料和重要信息,为加快芝麻野生种研究利用和远缘杂交育种进程奠定了坚实基础。
3.1 芝麻种间杂种的获得与鉴定
杂种胚培养是克服种间杂交障碍的重要手段[32-33]。由于种间生殖隔离障碍,芝麻杂种胚败育在不同的种间组合中发生的时期不同。张海洋等[15]研究发现,栽培种与野生种(和Schum)种间杂交因胚败育在大田状态下很难获得种间杂交籽粒,而采用胚培养技术,获得部分种间杂种。表明栽培种与这两个野生种的不亲和性发生于胚形成-后代植株发育的整个过程。本研究结果表明,采用胚拯救技术,共有18个组合获得了杂种植株,杂种后代特征多样。如,豫芝11号()与Ken1()的胚拯救效果好,正反交胚拯救杂种后代比率均较高,分别达到22.86%和69.92%。而×的130个杂交幼胚的拯救均未获得组培苗,这可能与部分种间杂交授精不完全或杂种夭亡有关。此外,胚拯救试验未获得×的杂交幼胚,子房壁也未出现膨大。说明2个种杂交授粉后,种胚可能无法形成,进而无法诱导子房壁膨大形成蒴果。但是,以母本,配置杂交组合,通过杂种幼胚拯救,组培苗成苗率提高到7.86%。表明以染色体数目多的母本配置种间杂交组合,幼胚拯救后,可能更易获得杂种植株。这与已报道结果较一致[15, 19, 34]。因此,推测通过染色体加倍或者桥梁亲本,结合种间杂交和胚培养技术,将可能获得更多的种间杂交材料,以显著提升芝麻野生种质资源的利用效率。今后研究组将对此作进一步的尝试。
为验证种间远缘杂交材料的真实性,张海洋等[15]筛选并开发了芝麻SSR引物HS209,用于区分和及后代材料;扩增杂种阳性率为95.83%。本研究利用基于芝麻基因组信息发掘的3对种间特异SSR标记[31],对组织培养获得的杂种植株进行分子鉴定。发现不同种中3对SSR标记均表现出了种间特异性,证实了种间杂交材料的真实性和SSR标记的特异性。此外,还发现采用HS94引物扩增×、×的杂种后代,出现了双亲以外的带型。推测这可能与杂交后代的遗传信息变异有关。近期研究发现,×种间杂交后代形成过程中,F2群体后代的基因组信息出现有染色体片段的重排和重组(结果尚未发表)。因此,开展芝麻种间杂交,获得更多遗传多样性丰富的杂交后代,是拓展芝麻栽培种遗传基础的重要技术手段和途径。此外,本研究对鉴定出的真杂种进行了根尖细胞染色体数目和特征分析,表明种间杂交材料的多样性和真实性,为深入了解杂种形成、败育机理及物种进化机制提供了材料和方法。
3.2 芝麻种间的不亲和性与野生种利用
芝麻是古老的油料作物。将野生种的优良性状导入芝麻栽培种一直是研究者的目标之一。但是受生殖隔离、种间亲缘关系较远等因素限制,芝麻栽培种与部分野生种以及野生种之间的远缘杂交研究进展缓慢[18, 26]。本研究发现,除与田间杂交可以获得正常有胚杂种外,其他18个种间杂交组合直接获得可育杂种的难度较大。表明芝麻种间的亲和性普遍较差。多年来,研究组采用栽培种和持续开展了种间杂交及回交研究[15]。调查结果显示,以野生种为回交亲本可以获得较多的可育后代,且性状更接近于栽培种亲本(数据未公开发表)。同时,以栽培种为回交父本,通过胚拯救技术连续开展回交,其多代后株系在茎、叶和花等器官形态方面,则较类似于栽培种,但育性较差(数据未公开发表)。本研究中,仅和2个野生种间能够获得较多的可育杂种后代;正反交杂交结蒴率虽有差异,但杂种后代F2中仍有可育株。表明两者虽然存在着一定的生殖隔阂,但亲缘关系较近,可以用于重要性状遗传分析和远缘杂交育种研究。为加快上述野生种质的利用,近期研究组开展并完成了对上述5种的染色体级别的基因组精细图构建[3],并建立了芝麻高效转基因技术和基因编辑技术[35]。今后将进一步开展芝麻野生种抗病抗逆重要信息发掘和资源应用,以推动芝麻抗病抗逆育种和远缘杂交育种研究进程。
4 结论
芝麻栽培种与4个野生种的杂交亲和性依次为>>>;野生种与3个野生种的杂交亲和性依次为>>与种间杂交可获得大量可育杂种后代。芝麻不同种间杂交后代在株高、株型等性状方面具有超亲优势。
[1] ASHRIA. Sesame breeding.Plant Breeding Reviews,1998,16: 179-228.
[2] AMOO S O, OKOROGBONA A O M, DU PLOOY C P, VENTER S L.//VICTOR K. Medicinal Spices and Vegetables from Africa. Salt Lake City, USA: Academic Press, 2017: 549-579.
[3] ZHANG H, WANG L, MIAO H, SUN Y. Genome sequencing of the wildspecies//MIAO H, ZHANG H, KOLE C. The Sesame Genome. Compendium of Plant Genomes. Cham, Switzerland: Springer, 2021: 275-281.
[4] MIAO H M, LANGHAM D R, ZHANG H Y. Botanical descriptions of sesame//MIAO H M, ZHANG H Y, KOLE C. The Sesame Genome. Cham, Switzerland: Springer, 2021: 19-57.
[5] PHAM T D. Analyses of genetic diversity and desirable traits in sesame (L. Pedaliaceae): Implication for breeding and conservation. Superlattices & Microstructures, 2011, 36(4/6): 563-571.
[6] 张海洋, 苗红梅, 李春, 魏利斌, 马琴. 芝麻染色体核型及似近系数分析. 植物学报, 2012, 47(6): 602-614.
ZHANG H Y, MIAO H M, LI C, WEI L B, MA Q. Analysis of sesame karyotype and resemblance-near coefficient. Chinese Bulletin of Botany, 2012, 47(6): 602-614. (in Chinese)
[7] Zhao R H, Miao H M, Song W Q, Chen C B, Zhang H Y. Identification of sesame (L.) chromosomes using the BAC-FISH system. Plant Biology, 2018, 20: 85-92.
[8] 苗红梅, 常淑娴, 张海洋, 黄进勇, 段迎辉. 芝麻营养生长期枯萎病抗性鉴定技术研究. 植物遗传资源学报, 2020, 21(2): 330-337.
MIAO H M, CHANG S X, ZHANG H Y, HUANG J Y, DUAN Y H. An evaluation technique of sesame resistance toWilt disease at vegetative stage. Journal of Plant Genetic Resources, 2020, 21(2): 330-337. (in Chinese)
[9] ZHANG H Y, MIAO H M, JU M. Potential for adaptation to climate change through genomic breeding in sesame//KOLE C. Genomic Designing of Climate-Smart Oilseed Crops. Cham, Switzerland: Springer, 2019: 374-376.
[10] PHAM T D, NGUYEN T T, CARLSSON A S, BUI T M. Morphological evaluation of sesame (SL.) varieties from different origins. Australian Journal of Crop Science, 2010, 4(7): 498-504.
[11] ZHANG H Y, MIAO H M, WANG L, QU L B, LIU H, WANG Q. Genome sequencing of the important oilseed cropL.. Genome Biology, 2013, 14(1): 401-409.
[12] HIREMATH S C, PATIL C G. Genome homology and the putative progenitor of sesame. Journal of Cytology and Genetics, 1999, 34: 69-74.
[13] NIMMAKAYALA P, PERUMAL R, MULPURI S, REDDY U K. Sesamum//KOLE C. Wild crop relatives: genomic and breeding resources, vol Oilseeds. Heidelberg, Berlin: Springer, 2011: 261-273.
[14] ASHRI A. Sesame (L.)//SINGH R J. Genetic resources chromosome engineering, and crop improvement, oilseed crops. Boca Raton, US: CRC Press, 2007: 231-289.
[15] 张海洋, 苗红梅, 张体德, 魏利斌, 李春, 王慧丽, 段迎辉, 琚铭. 芝麻栽培种与野生种(Asch,Schum & Thonn)种间杂交后代的生物学特性. 中国农业科学, 2013, 46(19): 3965-3977.
ZHANG H Y, MIAO H M, ZHANG T D, WEI L B, LI C, WANG H L, DUAN Y H, JU M. Biological characters of interspecific hybrid progenies betweenL. and wild relatives (Asch,Schum & Thonn).Scientia Agricultura Sinica, 2013, 46(19): 3965-3977. (in Chinese)
[16] KOBAYASHI T. Cytogenetics of Sesame ()// TSUCHIYA T, GUPTA P K. Developments in Plant Genetics and Breeding. 1991, 2(B): 581-592.
[17] KUMAR A K. Studies on karyoptype, genome size and genome relations in some species ofL. (Pedaliaceae)[D]. Dharwar, India: Karnataka University, 2003.
[18] JOSHI A B. Sesamum//JOSHI A B. Sesamum, Hyderabad, India: Indian Central Oilseed Committee Hyderabad, 1961: 9-10.
[19] 杨敏敏, 刘红艳, 周婷, 瞿洪浩, 杨远霄, 魏鑫, 左阳, 赵应忠. 芝麻栽培种与野生种()杂种F1的获得及特性鉴定. 中国农业科学, 2017, 50(10): 1763-1771.
YANG M M, LIU H Y, ZHOU T, QU H H, YANG Y X, WEI X, ZUO Y, ZHAO Y Z. Production and identification of F1interspecific hybrid betweenand wild relative. Scientia Agricultura Sinica, 2017, 50(10): 1763-1771. (in Chinese)
[20] MEHRA N. Sesame: its uses, botany, cytogenetics, and origin. Economic Botany, 1970, 24(1): 20-31.
[21] BEDIGIAN D. Cultivated sesame, and wild relatives in the genusL.//Bedigian D. Sesame: the genus. Medicinal and Aromatic Plants - Industrial Profiles series, Boca Raton, US: CRC Press. 2010: 33-77.
[22] TARIHAL R, SRIDEVI O, SHENOY V V, SALIMATH P M. Study of fertilization barriers in crosses betweenand its wild relatives. Indian Journal of Genetics and Plant Breeding, 2003, 63(2): 132-136.
[23] KUMARI B M, GANESAMURTHY K. Study of reproductive compatibility and morphological characterization of interspecific hybrids insp. African Journal of Agricultural Research, 2015, 10(9): 911-918.
[24] RAJESWARI S, THIRUVENGADAM V, RAMASWAMY N M. Production of interspecific hybrids betweenthonn andL. through ovule culture and screening for phyllody disease resistance. South African Journal of Botany, 2010, 76(2): 252-258.
[25] 赵瑞红, 苗红梅, 马琴, 陈成彬, 宋文芹, 张海洋. 芝麻野生种与栽培种核型比较分析. 南开大学学报(自然科学版), 2018, 51(5): 27-36.
ZHAO R H, MIAO H M, MA Q, CHEN C B, SONG W Q, ZHANG H Y. Karyotype comparison analysis of the wild speciesand the cultivated Sesame. Acta Scientiarum Naturalium Universitatis Nankaiensis (Natural Science Edition), 2018, 51(5): 27-36. (in Chinese)
[26] CARLSSON A S, CHANANA N P, GUDU S, SUH M C, WERE B A. Sesame//KOLE C, HALL T C. Compendium of transgenic crop plant-Transgenic oilseed crops. Texas, USA: Wiley Blackwell, 2008: 227-246.
[27] ZHANG H Y, LANGHAM D R, ZAHO Y Z, KHALAFALLA A, MIAO H M. Traditional breeding in sesame//MIAO H M, ZHANG H Y, KOLE C. The Sesame Genome. Cham, Switzerland: Springer, 2021: 145-158.
[28] 石淑稳. 芝麻野生种与栽培种的交配能力. 中国油料, 1993, 2: 18-21.
SHI S W. Cross compability between wild and cultivated sesame. China Oil, 1993, 2: 18-21. (in Chinese)
[29] 苗红梅, 琚铭, 魏利斌, 马琴, 张海洋. 芝麻愈伤组织诱导与植株再生体系的建立. 植物学报, 2012, 47(2): 162-170.
MIAO H M, JU M, WEI L B, MA Q, ZHANG H Y. Establishment of sesame callus induction and shoot regeneration system. Bulletin of Botany, 2012, 47(2): 162-170. (in Chinese)
[30] PETERSON R, SLOVIN J P, CHEN C. A simplified method for differential staining of aborted and non-aborted pollen grains. International Journal of Plant Biology, 2010, 1(2): 66-69.
[31] WEI L B, MIAO H M, XU F F, KONG J J, ZHANG H Y. Chinese sesame cultivars, DNA fingerprinting, and two-dimensional barcodes using SNP, InDel, and SSR markers. Crop science, 2017, 57: 1-7.
[32] SUDHAKER D, SREERANGASAMY S R, 梁根庆. 芝麻种胚培养. 中国油料作物学报, 1990(3): 102.
SUDHAKER D, SREERANGASAMY S R, LIANG G Q. Cultivation of sesame embry. Chinese Journal of Oil Crops, 1990(3): 102. (in Chinese)
[33] 瞿桢. 芝麻远缘杂种胚胎的营救和植株再生. 中国油料, 1994(1): 33-35.
QU Z. Embryo rescue and plant regeneration of distant hybrid in sesame. China Oil,1994(1): 33-35. (in Chinese)
[34] 刘红艳, 赵应忠. 芝麻栽培种与野生种种间杂交亲和性研究. 中国农学通报, 2011, 27(9): 156-159.
LIU H Y, ZHAO Y Z. Studies on the hybridization compatibility between cultivated sesame and its wild species.Chinese Agricultural Science Bulletin, 2011, 27(9): 156-159. (in Chinese)
[35] MIAO H M, JU M, WANG H L, ZHANG H. Tissue culture and genetic transformation in sesame//MIAO H, ZHANG H, KOLE C. The Sesame Genome. Cham, Switzerland: Springer, 2021: 131-144.
Analysis of Cross Compatibility Variation among DiverseSpecies and Biological Characteristics of the Interspecific Hybrid Progenies
JU Ming, MIAO HongMei, HUANG YingYing, MA Qin, WANG HuiLi, WANG CuiYing, DUAN YingHui, HAN XiuHua, ZHANG HaiYang
Henan Sesame Research Center, Henan Academy of Agricultural Sciences/Henan Key Laboratory of Specific Oilseed Crops Genomics/Shennong Laboratory, Zhengzhou 450002
【Objective】The research aims to explore the cross compatibility between differentspecies and analyze the biological characteristics of interspecies hybrid progeny so as to supply the foundation for efficient application of wild sesame species. 【Method】A sesame cultivar Yuzhi 11 (, 2n=26) and 4 wildspecies including(2n=32),(2n=32),(2n=32), and(2n=64) were applied to construct interspecies cross combinations using diallel hybridization method by artificial pollination in the field.Embryo rescue method was also used to obtain interspecific hybrid F1. Interspecific hybrid compatibility was compared based on hybrid capsule formation rate. Botanical characters of hybrids were observed during flowering and mature stages. Pollen fertility was assessed using Alexander staining method. Chromosome number and karyotype characteristics of root somatic cells of hybrids were observed using smear chromosome preparation technique. Specific and polymorphic SSR primers inwere used to analyze the molecular difference in interspecific hybrids.【Result】 Twenty positive and reciprocal cross combinations were constructed for the 5species. A total of 2091 flowers were pollinated and 370 hybrid capsules were harvested. As to the female parents with more chromosomes, hybrid capsules were more easily obtained. The cross compatibility among the 5species significantly varied from 1.18% (×) to 63.33% (×). F1plants of 9 combinations produced hybrid seeds, while the ratio of pollen sterility of F1progeny ranged from 35.21%-100.00%. The cross×presented the highest sterility ratio to 87.68%. Hybrid progeny exhibited the obvious heterosis over parents in plant height, plant type, and some key agronomic traits. As to the positive and reciprocal hybrid F1derived from sesame cultivar and the wild species, leaf shape, flower shape, and flower color showed partial characters of both parents. The cross compatibility between sesame cultivar (n=13) and the 3species with chromosome group n=16 ranked as>>. The cross compatibility between wild species(n=32) and the 3 species with n=16 ranked as>>. Among the 5species, the genetic relationship betweenandis relatively closest. The chromosome number of root tip cells of some hybrid plants is consistent with the theoretical value calculated from the parents. Screening results of the 3 pairs of polymorphic SSR primers indicated that 99.66% of obtained F1plants are true hybrid. Chromosome karyotype and SSR marker screening results reflected the genetic difference and characters ofspecies. 【Conclusion】Among the 5species, the cross compatibility varies significantly and the heterosis of interspecific hybrid is obvious. Of which onlyandhave the relatively closest genetic relationship and could be directly applied for elite germplasm creation and interspecific hybrid breeding in. Reproductive isolation barriers exist in other cross combinations. Some techniques including embryo rescue and molecular marker application should be used to achieve the utilization of wildspecies for sesame breeding.
sesame; wildspecies; interspecific hybridization; botanic character; molecular assessment

10.3864/j.issn.0578-1752.2022.20.003
2022-05-29;
2022-07-18
国家现代农业(特色油料)产业技术体系(CARS-14)、河南省重大科技专项(201300110600)、中原科学家工作室建设项目(092101211100、214400510026)、中原科技创新领军人才项目(214200510020)、河南省农业科学院优秀青年项目(2020YQ19)、科技创新团队项目(2022TD05)、科技创新创意项目(2020CX02)、自主创新项目(2021ZC70)
琚铭,E-mail:jumingzz@163.com。通信作者张海洋,E-mail:zhanghaiyang@zzu.edu.cn
(责任编辑 李莉)
