极小种群野生植物毛果木莲研究进展
2022-12-10吴哲荣艾星梅余向鑫代嫚婷安星霖王斌
吴哲荣,艾星梅,余向鑫,代嫚婷,安星霖,王斌
(1.西南林业大学园林园艺学院,云南 昆明 650224;2.云南民族大学澜湄国际职业学院,云南 昆明 650504;3.云南省农业科学院生物技术与种质资源研究所,云南 昆明 650205)
毛果木莲(Manglietia ventiiN.V.Tiep)隶属木兰科(Magnoliaceae)木莲属(Manglietia),为常绿乔木,高可达36 m,胸径可达80 cm,是自然分布范围十分狭窄的珍稀濒危野生植物[1-2]。其演化历史悠久,是一种相对较原始的木兰型植物,对探索木兰科植物系统演化历程具有重要的科研意义[2]。此外,毛果木莲的木材为散孔材,结构细致,耐腐抗虫能力强,是工业建筑、高档家具等的优等用材树种[3]。生长速度较快,树形高大,花色纯白而芳香,可作为优良的园林庭园观赏树种[4]。化学成分研究表明,毛果木莲主要产生的次生代谢物是单萜类和倍半萜类化合物,这两种化合物具有较高的抗癌活性,该植物也可作药用植物[5-6]。由此可见,毛果木莲在科研领域、经济利用、景观生态等诸多方面具有较高的价值与意义。但由于长期与反复的干扰,其适宜生境进一步破坏和丧失以及自身繁育能力退化等原因,导致其种群数量急剧减少[1-2]。早在1999年就已被列为国家Ⅱ级重点保护野生植物,2012年被列为云南省第一批优先拯救保护的极小种群野生植物,在2016年出版的木兰科红色名录中被认定为“濒危种(EN)”[7-9]。近年来,随着极小种群保护的发展与毛果木莲研究的深入,对毛果木莲的系统分类学、生态学、繁殖生物学、遗传多样性、濒危机制、资源保护等方面的研究工作迅速发展,通过对相关工作进展进行综述,并对今后的研究工作进行展望,旨在为毛果木莲的种质资源保护和可持续利用提供理论支持,还可为其他极小种群野生植物的保护研究提供参考。
1 系统分类学研究
1980年Tiep[10]在云南省屏边县首次采集到了毛果木莲的标本。根据花柱、心皮和花粉等形态特征,将毛果木莲定义为一个新的物种后归类到木兰科木莲属,发表后极少再有报道。1996年吴征镒院士根据云南省金平县采集的模式标本在中国植物志上刊登了一个新种并命名为Manglietia hebecarpa[11]。M.ventii和M.hebecarpa两个模式标本因生存环境异同导致在形态上存在一定的差异,曾一度被误认为是两个独立的种[12],后在2006年出版的云南植物志中由吴征镒院士正式归类为同一种,因此毛果木莲拉丁学名存在异名现象[13]。此外,由于木兰属与木莲属植物在外部形态与内部结构存在大量重叠交叉现象,因此在属分类的界定上存在不同观点[14-18],这导致毛果木莲的名称先后经历过以下分类学处理:Manglietia hebecarpa[11]、Magnolia hebecarpa[12]、Manglietia venti[2]和Magnolia venti[9]。从现有的研究资料来看,木兰科在科内分类界限上仍存在不同分歧,国内外提出了大量针对木莲属的系统分类位置的理论依据,但这些分类系统并没有得到人们广泛的认可与支持[19-22]。就目前而言,国内多采用刘玉壶[19]对木兰科的分类系统,将木莲属独立出来。而外国学者发表相关文献多采用的拉丁学名为Magnolia ventii,相对认可NOOTEBOOM[20]的系统。
形态解剖学通常是构建分类系统的主要依据,随着生物技术的不断突破与发展,不同的学者又从孢粉学、细胞学、分子生物学等多学科交叉展开了对毛果木莲的系统分类学研究。司马永康等[23]认为幼叶卷叠式特征具有十分重要的系统分类学意义,毛果木莲的幼叶在芽中对折且直立,这是木莲属植物的典型特征。薛晓明等[24]通过扫描电子显微镜对毛果木莲的花粉形态进行观察,其花粉粒的纹饰由褶皱和小穿孔组成,其形状与同属的毛桃木莲(Manglietia moto)、巴东木莲(Manglietia patungensis)和华木莲(Manglietia decdua)的形态较为接近。其花粉粒为两侧对称,花粉沟位于远极面,长达两极,这与已发表报道的其它木莲属植物的花粉特征基本类似[25]。叶珊珊等[26]通过PAGE技术分析了毛果木莲的醇溶蛋白特征谱带,结果发现毛果木莲与十余种木兰科种子具有相似的遗传基础。为了揭示毛果木莲在木兰科中的进化位置,Wang等[27]对毛果木莲叶绿体基因组与已报道的45个木兰科植物基因组进行对比,结果表明毛果木莲与香木莲(Manglietia aromatic)和大叶木莲(Manglietia megaphylla)的亲缘关系最为密切。王猛等[28]通过ISSR技术对多种木兰科植物的遗传多样性进行了研究分析,研究结果表明毛果木莲与山玉兰、木莲属、含笑属的部分物种亲缘关系相对较近,与刘玉壶[19]提出的分类系统存在明显异同。
综上所述,对于毛果木莲系统分类主要分歧在于木兰科内属之间的界定。尽管形态、解剖学、细胞学为木莲属的独立提供了许多有利的证据,但从分子生物学的角度分析结果表明木莲属植物与木兰属植物存在大量重叠与交叉,应合并到木兰属。目前关于木兰科属的界定还没有一致的认可,我们认为不同性状在木兰科各类群中呈网状分布,属之间的界定不应局限于单一性状,应结合多种性状来综合考虑。因此,结合多种性状研究制定一个国际统一且客观自然的系统分类标准对确定毛果木莲分类学地位显得尤为重要。
2 生态学研究
2.1 自然地理分布
毛果木莲是分布范围十分狭窄的珍稀濒危野生植物,目前零星散状分布在红河州的金平苗族瑶族傣族自治县、屏边苗族自治县、河口瑶族自治县及元阳县境内[29-30]。根据中国科学院昆明植物所与金平分水岭国家级自然保护区相关研究团队开展的样方法调查推算,毛果木莲个体数约为2452株,其中金平县有2024株、屏边县有366株、河口县有60株、元阳县有2株,大部分的植株分布于保护区外[30-31]。野生植株主要分布于东经103°31'33″~103°51'42″,北纬22°38'29″~22°53'56″,海拔在800~1500 m的山地季风常绿阔叶林中及农田村落旁[31]。
2.2 生境特征
毛果木莲分布区内干湿季节分明,夏不炎热,冬季温暖,雨量多,湿度大。年平均气温16.4~22.6℃,年降雨量1650~2960 mm,雾期相对较长[2]。植株生长的土壤质地以酸性的山地黄壤或黄棕壤为主,不同分布点各项指标的含量没有表现出相似的趋势,土壤的肥力变化较大[32]。毛果木莲与橄榄(Canarium album)、云树(Garcinia cowa)、黑桫椤(Gymnosphaera podophylla)、刺栲(Castanopsis hystrix)、川滇木莲(Manglietia duclouxii)、网叶山胡椒(Lindera metcafiana var.dictyophylla)、云南拟单性木兰(Parakmeria yunnanensis)、山乌桕(Sapium discolor)、青皮树(Altingia excelsa)、草鞋木(Macaranga henryi)、大叶桂(Cinnamomum iners)、黄心树(Machilus bombycina)、红木荷(Schima wallichii)、倒卵叶黄肉 楠(Actinodaphne obovata)和鸡 脖 子(Artocapus tonkinensis)等植物混生成林[31]。常见于人工林、草果地和天然次生林以及村落农田边与乡村道路边。以单株散生、散生林或疏林的形式存在且周围少见幼树,生境相对脆弱且易受到人为干扰破坏[6]。
2.3 种群生态学
毛果木莲的种群数量较少且分布点相对孤立,和其他乔木混生成林多不容易被注意到。野外不同分布区内幼苗和幼树都相对较少,徐清等[33]推测可能是由于毛果木莲自然生境的持续性恶化,其幼苗生长所需要的光环境受到影响,从而无法完成自然更新。毛果木莲分布区多邻近道路、村落等人为活动较频繁的区域,因此人为干扰和破坏较多,这种情况也让其幼苗很难生存[31]。王斌[32]对分布在金平县、屏边县和河口县的毛果木莲从分布格局、径级结构、年龄结构以及种群结构进行动态量化分析,结果表明该种群整体呈倒梯形,种群内幼苗与幼树相对较少,从长远来看不利于毛果木莲种群的更新与延续。
3 繁殖生物学研究
植物繁殖是植物进化过程的核心,是植物延续种族最基本的行为过程,也是生物群落变化和生态系统演替的基础。但繁殖也是植物生命周期中相对脆弱的阶段,植物繁殖过程中各个环节相互影响,一旦某一个环节出现障碍,就有可能导致繁殖过程的失败,进而使种群的生存面临危机[34-37]。因此,深入了解毛果木莲的繁殖生物学特性对探索其濒危机制与进行有效保护具有重要的研究意义。
3.1 生殖生理学研究
毛果木莲通常在4月底至5月中旬开花,5月初是开花的高峰期,花期恰逢当地雨季,属于雌蕊先熟、夜间开花的植物。花开时面朝地面呈钟形,推测可能是为了避免开花期间雨水冲刷而对花粉的形成产生影响,同时在访花昆虫到来时可以起到避雨的效果。单花花期持续大约6天,单花的开花过程为“开放-关闭-重新开放”的节奏,在该物种的花期内可观察到五个明显的阶段(表1)[38]。
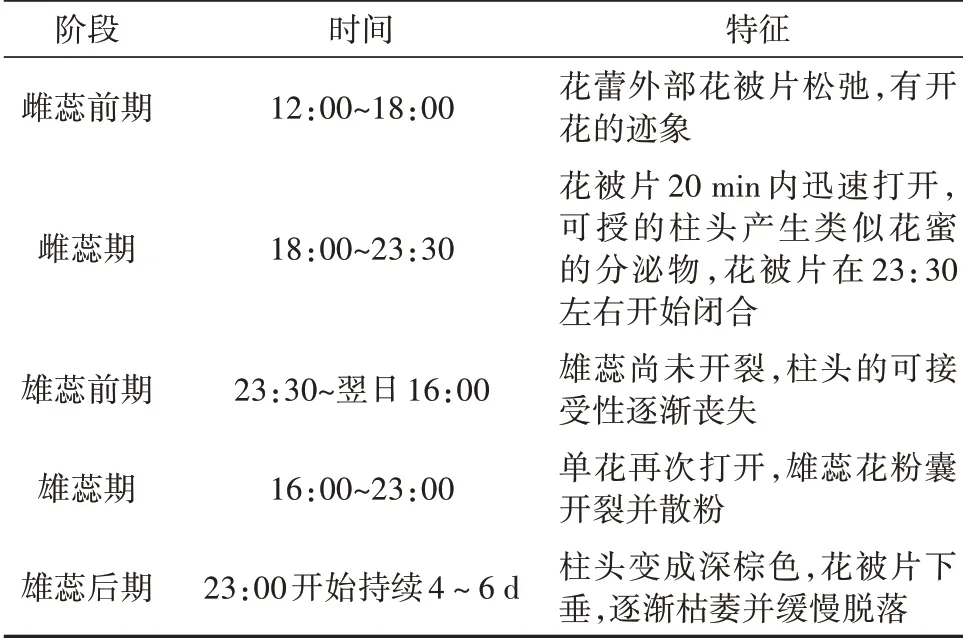
表1 毛果木莲花开时律Tab.1 Flowering process of M.ventii
毛果木莲严重依赖传粉者,主要通过花香气味引导昆虫授粉,但目前仅发现象鼻虫和金龟甲虫是有效的授粉昆虫[38]。然而,由于小的和支离破碎的生境现状存在导致昆虫的访问频率可能较低进而限制了种群的发展[39]。相比之下,通过人工进行异花授粉可明显提高种子产量。这说明毛果木莲生殖繁育过程中存在花粉限制,推测自然种群中的低结实率是花粉沉积不足的结果[32]。此外,大多数濒危的木莲属植物都存在不同程度的败育现象[40]。徐涛等[41]发现毛果木莲花粉发育也存在败育现象,花粉的败育率达到29%,但根据毛果木莲花粉量很大这一特征推测,花粉败育对其结籽率较低的影响是有限的。
3.2 种子传播学研究
种子传播是其迁移到其它地方的主要手段之一,是植物自然更新的关键阶段[42]。毛果木莲的果实在9月份成熟后蓇葖开始脱裂,种子借助重力或风力进行传播[6]。然而,发育的种子大部分依赖于动物传播得以实现自然更新,动植物之间存在明显的互惠网络[43]。种子传播会因动物取食方式、消化系统、运动距离等方面存在不同的差异而产生不同的效果[44]。通过团队野外实地调查,发现一些啮齿类动物会对毛果木莲的种子进行取食,因此存在部分种子受到破坏而不能萌发。观察其自然落果状态,采集自然落地的果实并剥离种子发现较多的种子储存在蓇葖果的果壳中而不能及时落入土壤中,这部分种子可能会因为缺乏适宜萌发条件而导致腐烂失活。这也可能致使树下缺乏相应数量的种子从而无法产生土壤种子库。此外,毛果木莲存在肉质红色假种皮,对鸟类动物在视觉上有一定吸引力,可促进其种子远距离扩散,然而毛果木莲种子传播对此方面的内容未见报道,还需要后续深入研究。
3.3 种子的储存及萌发
树木的种子和苗期被认为是植物生活史周期中相对脆弱的阶段,关系到种群的生存与发展[45]。收采时期的把控对种子的质量有重要影响,而种子的萌发与种子的质量有着密切关联,及时地收采果实能有效提高种子的萌发率[46]。毛果木莲果实成熟季节在8~9月份,当蓇葖果由黄绿色变为红褐色,蓇葖少量开裂并露出鲜红色的种粒,种子达到形态成熟状态[47]。毛果木莲种子内含有挥发性芳香油物质极易干缩变质,因此不能干藏。其种子也不宜久藏,萌发活力会因贮藏而明显减弱。一般当年采种,翌年1月播种或随采随播[48]。Wang等[49]研究粗梗木莲也提出类似的观点,所有贮藏种子的萌发活力都明显减弱。
种群的更新能力受种子自然萌发率的影响,而种子自身特性和环境因子对种子的萌发率起着决定性作用[50]。自然条件下,毛果木莲新鲜成熟的种子萌发率较低,种子存在生理休眠,严重影响其出苗率。王斌[32]测定了温度改变对毛果木莲的萌发的影响,在变温25/15℃条件下毛果木莲种子的萌发率最高,其次是在恒温(25℃)条件。研究比较了毛果木莲种子在(50 mg·L-1、100 mg·L-1、200 mg·L-1)不同浓度的赤霉素处理下的萌发试验,结果表明不同浓度赤霉素浸种处理的毛果木莲种子发芽率均高于对照,但不同实验组之间没有显著性差异。参考其余木兰科植物赤霉素对种子的影响[51-52],推测可能是赤霉素浸泡时间过短导致。不同浓度细胞分裂素处理的毛果木莲种子均未能萌发,与对照比较表现为抑制作用[32]。
3.4 生长特性研究
楚永兴等[48]根据毛果木莲种子自身特性提出种子萌发需要相对湿润荫蔽的生境,而幼苗的生长发育则需要充足的光照环境。当植物吸收的光能超过了一定的限制,过量的光能会导致植物发生光合作用的光抑制[33,53]。徐清等[33]通过实验证明过强或过弱的光照都不利于毛果木莲幼苗的生长与发育,其最适宜的光照环境为30%的光照强度。张光飞等[53]得到类似的结果,并提出在保证水分充足的条件下,毛果木莲幼苗对光强具有一定的适应性。张畋等[54]对毛果木莲种子育苗研究表明7月到9月是苗木生长的旺季。毛果木莲和同期育苗的山玉兰、香木莲、华盖木、云南拟单性木兰这几种同科植物相比苗期生长相对较快。何艾蔓等[55]对毛果木莲1年生种子苗生长规律进行研究,发现其株高速生期为6月至8月,叶片的速生期为6月至9月,叶片生长在10月份以后开始呈下降趋势。
4 遗传多样性研究
毛果木莲种群数目与有效居群数量都相对较小,这将影响基因流和遗传漂变等生命进化过程,从而导致植物生存几率降低的风险大增。种群的遗传多样性水平也在一定程度上制约着该物种应对环境持续变化的适应能力[56]。因此,研究毛果木莲的遗传多样性以及遗传结构对于制定保护策略是至关重要的。王斌[32]、李娇婕[57]和王猛等[28]分别利用SSR与ISSR分子标记技术对毛果木莲的遗传多样性以及遗传结构进行了研究。结果发现,同木兰科其他植物相比,毛果木莲保持了相对较高的遗传多样性水平与较高的遗传分化。毛果木莲的遗传变异大部分存在于居群内,而仅有27.65%的遗传变异存在于居群间[32]。杨林[9]等对毛果木莲叶绿体基因组密码子偏好性分析发现毛果木莲叶绿体基因组偏好使用A/T结尾的密码子且密码子的使用偏性与GC含量密切相关。
5 濒危机制
5.1 外部因素
毛果木莲为云南特有的极小种群,零星散状分布在山谷常绿阔叶林的山坡、路边以及农田村落旁。居群多呈片状间断分布,未见连续大面积分布的居群,生境敏感脆弱易被破坏,居群间相互隔离[31]。毛果木莲是当地优等用材树种,因此偷盗砍伐的现象时有发生,盲目毁林开荒过度以及林下种植草果等人类活动导致其赖以生存的环境被破坏,当地居民保护意识薄弱等,这些是毛果木莲濒危的重要原因[6]。此外,毛果木莲种子成熟脱落后,由于种皮颜色鲜艳、种质鲜美而遭到林间动物取食,也加剧了毛果木莲种群的濒危[32]。
5.2 物种自身因素
毛果木莲开花量大,但花期与当地雨季重叠阻碍了花香的扩散,以及对传粉者活动的高度依赖进而导致自然结实率较低[38]。结实存在大小年现象,种子存在休眠特性,属于顽拗性种子,受幼苗成长的自身特性影响,居群存在自交衰退现象等,从而导致毛果木莲面临极度濒危的生存现状[31-33,53]。
6 资源保护
6.1 资源保护现状
近几十年,很多政府机构与科研学者为保护毛果木莲做出了巨大贡献,随着极小种群野生植物的提出,以种群为基本单元的保护理念和方法,倡导实施对极小种群的种群数量、结构进行动态管理与调节,实现种群稳定发展的目标,这为毛果木莲的保护提供了科学的指导[58]。通过建设保护小区、迁地保护、近地保护、种质资源保护与野外回归等相关保护措施(表2)来减少人为活动对毛果木莲的影响,从而促进毛果木莲种群的健康发展[30]。

表2 毛果木莲保护现状Tab.2 Protection status of M.ventii
6.2 保护策略
6.2.1 全面系统开展科学研究
对保护极小种群野生植物而言,只有在对其生态学与生物学等基本特征充分认识的基础上,才能科学地采取相应的有效保护措施[59]。因此,加强毛果木莲的相关研究是对其进行有效保护不可或缺的。此外,还应通过开展毛果木莲扦插繁殖研究、组织培养试验,从而快速扩大种群数量,为其种质资源保护和可持续利用提供保障[60]。
6.2.2 加强各类保护措施实施
气候改变可能会使物种产生新的遗传变异进而导致发生显著的进化改变。因此,在进行迁地保护时,应考虑迁移地气候的相关影响[61-62]。对于就地保护,应强化对已建成保护区的有效管护、种群及生境动态监测、基础数据科学采集等。尽快增加保护小区数量,针对散生于村庄、田边地头的毛果木莲通过挂牌建档保护[30]。
6.2.3 完善法律法规及加强宣传
完善极小种群野生植物的相关保护政策,完全落实可实施方案步骤,明确相关部门的责任制[63]。通过多种途径对毛果木莲资源保护的意义进行宣传教育,禁止过量的种子采集和木材盗伐等人为干扰,引导提高在毛果木莲分布区生活群众的保护意识,最终达到对毛果木莲资源的有效保护的目的[64]。
7 研究展望
随着毛果木莲被列入优先拯救保护的极小种群野生植物之后,人们对毛果木莲的濒危机制得到了一定的认知。通过大量科研机构与研究人员的共同努力,在毛果木莲生态、生物学特性方面取得大量成果。建立自然保护区、迁地保护与挂牌建档等各类保护措施的实施,使毛果木莲的保护也取得了阶段性成效,其种群数量得到了显著提升。然而,关于毛果木莲的研究也存在一定的局限性,在种子基础特性与无性繁殖方面的研究近乎空白,其种子自然萌发率低是否与种子内部物质有关?无性繁殖相对于种子萌发是否更容易扩大其种群数量?这些问题的解决将对毛果木莲的保护产生积极的影响。因此,未来的研究应加强对种子内部成分及萌发过程中内部各类内源激素、营养物质、酶含量等动态变化的测定。开展毛果木莲扦插繁殖研究、组织培养试验等。在种子传播、植物光合特性、化感效应以及根系与微生物群落关系等领域的研究深度相对较浅,属于基础探索阶段,分析其原因可能与野外种群个体数量相对较少且分布不集中,导致相关数据采集难度较大,未来对这些方面还需要展开更加深入的研究。此外,还应深化毛果木莲的资源开发与利用。加强引种适应性研究,让毛果木莲在园林建设中得到广泛应用,这也是一种有效的保护措施。通过化学成分分析研究,从为毛果木莲药用开发提供科学的理论指导。综上所述,应从多方面、多角度来展开对毛果木莲的深入研究,全面了解其相关特性,深化资源开发,为其种质资源保护与利用提供保障。
