长白猪群体生长性状的遗传参数及进展分析
2022-12-02闫翘楚林清冯雪燕苏展勤吴细波司景磊张哲李加琪
闫翘楚,林清,冯雪燕,苏展勤,吴细波,司景磊,张哲,李加琪
(1.华南农业大学动物科学学院/国家生猪种业工程技术研究中心,广东 广州 510642;2.广西农垦永新畜牧集团有限公司,广西 南宁 530022)
【研究意义】遗传评估是猪遗传育种工作的重要环节,是对种猪进行准确选择的依据[1]。遗传参数估计作为遗传评估工作的重要依据之一[2],在育种规划决策和育种目标规划、育种值估计中具有重大意义[3-4]。遗传参数是反映群体总体遗传特征的参数,描述数量性状的3 个基本遗传参数分别是遗传力、重复力和遗传相关。猪的数量性状包括繁殖性状、胴体性状、肉质性状和生长性状等,其中生长性状包括达目标体重日龄、平均日增重、活体背膘厚和眼肌面积等性状。作为一类重要的经济性状,猪生长性状的表达对企业的经济效益影响较大[5]。平均日增重和达目标体重日龄反映了猪的发育情况和生长速度,影响企业的养殖成本和经济收益。在种猪生产性能的测定方面,达100 kg 体重背膘厚和达100 kg 体重眼肌面积已被纳入全国种猪遗传评估方案的生长性能测定项目。背膘厚和眼肌面积能够较为客观地反映种猪肌肉的生长发育情况,此外,背膘厚还能反映母猪体况,为饲养阶段的划分和饲料营养的调控提供参考[6]。【前人研究进展】当前在猪遗传繁育体系中,长白猪因其良好的繁殖性能常用作二元杂母本。作为瘦肉型种猪,长白猪生长性能优异,与地方猪种杂交能发挥杂种优势[7]。如以长白猪为父本、金华猪为母本杂交育成的长金猪背膘厚明显小于以巴克夏猪为父本、金华猪为母本杂交育成的巴金猪[8]。大白猪和长白猪遗传力的相关研究[9]结果显示,达100 kg 体重日龄、日增重和背膘厚性状属于中高遗传力性状。在对猪生长性状遗传相关研究中,长白猪达目标体重日龄和达目标体重平均日增重遗传相关性较强且为负遗传相关[2,9]。【本研究切入点】目前,在对长白猪生长性状遗传参数的研究中,有关达100 kg 体重日龄、日增重和背膘厚的研究[9]较多,但对与产肉性能密切相关的性状,如达100 kg 体重眼肌面积的研究较少。因此,对长白猪关键生长性状,如达100 kg 体重日龄、达100 kg 体重眼肌面积等进行遗传参数评估,有利于为实施精准选种选育、开展准确的遗传评估提供参考。【拟解决的关键问题】本研究基于2002—2020 年间广西某种猪场核心育种群长白猪的性能测定记录,利用R 软件[10]分析与生长性状有关的固定效应,利用DMU 软件[11-12]估计相关性状的遗传参数和育种值,同时评估性状的表型进展和遗传进展趋势,以期为长白猪后续育种工作提供理论依据。
1 材料与方法
1.1 数据来源及处理
选取来自广西某种猪场核心育种群长白 猪2002—2020 年间共计41 490 条生长性能测定记录,性状包括达100 kg 体重日龄(DAYS_100)、达100 kg 体重日增重(ADG_100)、达100 kg 体重眼肌面积(LMA_100)和达100 kg体重背膘厚(BFT_100),以及测定场、出生日期、性别、出生胎次和系谱等信息。
为确保后续分析的可靠性,剔除原始数据的异常值,包括系谱中胎次为0 记录的异常个体、表型均值μ±3σ2之外的个体、测定场小于1 500 条记录的个体。为确定出生年份和季度对表型的影响,将出生季节分为春季(3—5 月)、夏季(6—8 月)、秋季(9—11 月)和冬季(12 月至次年2 月),出生胎次分为1、2、3、4、5、6胎及6 胎以上。
1.2 统计模型
1.2.1 固定效应显著性检验 利用R 软件[10]的aov 函数对测定场、出生年份、出生季节、性别和出生胎次5 个因素分别进行方差分析,确定各生长性状的固定效应和随机效应。分析模型如下:

式中,yijklmn为第n个种猪生长性状表型值,μ为群体均值,Hi为测定场次效应,Yj为出生年份效应,Sk为出生季节效应,SEXl为性别效应,Pm为出生胎次效应,eijklmn为个体随机残差;i、j、k、l、m、n分别为不同固定因素的水平。
1.2.2 遗传参数和育种值估计模型 利用DMU软件[11-12]的多性状动物模型,通过平均信息法估计遗传参数和育种值。分析模型如下:

式中,y为种猪个体生长性状的观测值向量,b为种猪个体固定效应向量(测定场、出生年份、出生季节、性别、出生胎次),a为种猪个体加性遗传值向量,e 为随机残差向量,X、Z分别为对应于b、a的结构矩阵。其中,a~N(0,Aσ2a),e~N(0,Iσ2e),A为基于系谱的个体间亲缘关系矩阵,σ2a为个体加性遗传方差,I为单位矩阵,为残差方差。
2 结果与分析
2.1 表型数据描述性统计
本研究对长白猪种猪共41 490 条原始记录经过数据质控处理,性状DAYS_100、ADG_100、LMA_100 和BFT_100 的记录数均达到30 000 条以上,表型平均值分别为169.43 d、584.49 g、38.16 cm2和11.42 mm(表1)。性状DAYS_100和ADG_100 的变异系数较小,在7.97%~8.02%之间,BFT_100 的变异系数较大,为18.21%。

表1 长白猪群体达100 kg 体重生长性状表型统计性描述Table 1 Descriptive statistics of the phenotype for growth traits of Landrace pigs population at 100 kg body weight
对各固定效应进行方差分析,结果(表2)发现测定场、出生年份、出生季节、性别和出生胎次均对性状DAYS_100、ADG_100、LMA_100和BFT_100 存在极显著影响(P<0.01)。
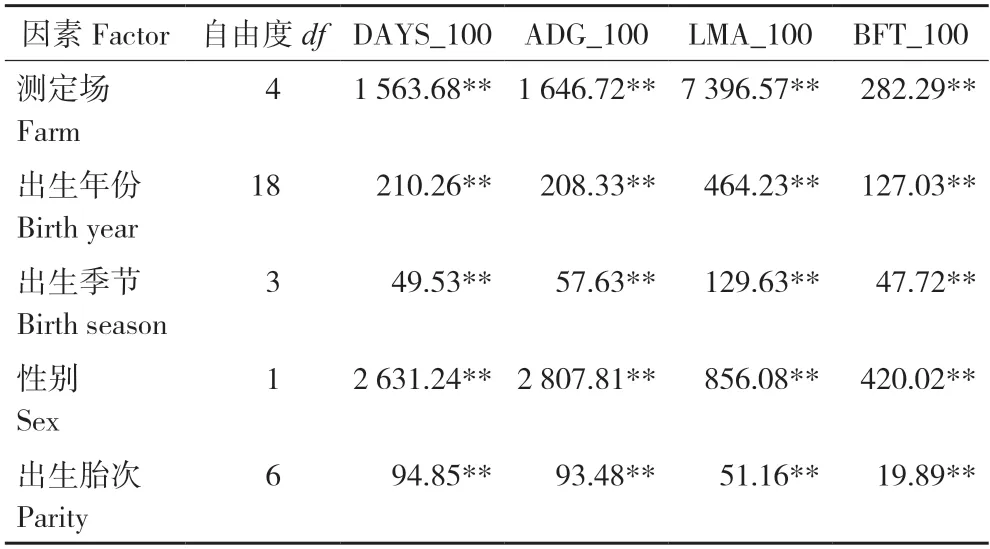
表2 长白猪群体达100 kg 体重生长性状方差分析F值Table 2 F value in the variance analysis for growth traits of Landrace pigs population at 100 kg body weight
2.2 遗传参数估计
由表3 可知,长白猪生长性状均在正常范围内,DAYS_100、ADG_100、LMA_100 和BFT_100的遗传力分别为0.399、0.391、0.433 和0.421,均为中高遗传力性状。由表4 可知,性状DAYS_100 和ADG_100 的遗传相关系数为-0.997,表型相关系数为-0.992。其他生长性状之间的遗传相关系数和表型相关系数均很小。
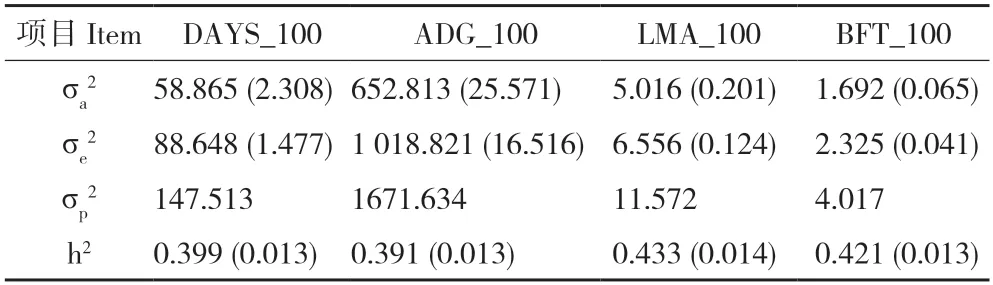
表3 长白猪群体达100 kg 体重生长性状方差组分及估计遗传力Table 3 Variance components and estimated heritability for growth traits of Landrace pigs population at 100 kg body weight

表4 长白猪群体达100 kg 体重生长性状遗传相关与表型相关分析Table 4 Analysis of genetic correlations and phenotypic correlations for growth traits of Landrace pigs population at 100 kg body weight
2.3 表型进展与遗传进展
4 个性状的表型均值和遗传进展分别通过计算不同年份的性状表型和估计育种值均值得到。由图1 可知,2011 年性状DAYS_100 和ADG_100的估计育种值均值分别达到4.991 d 的最高值和-16.817 g 的最低值;2020 年二者又达到各自的最低值和最高值,分别超过-6 d 和20 g;LMA_100 的遗传进展和表型进展趋势基本一致,在2008—2011 年快速上升,之后小范围波动,2017—2020 年一直呈下降趋势;BFT_100 的估计育种值均值在2002—2008 年稍有下降,随后3 年上升,总体呈略微上升趋势。

图1 长白猪群体达100 kg 体重生长性状表型进展和遗传进展Fig.1 Phenotypic progress and genetic progress for growth traits of Landrace pigs population at 100 kg body weight
从遗传进展变化幅度看,ADG_100 和DAYS_100 的遗传进展变化较大,ADG_100 从2002 年的2.006 g 增加到2020 年20.931 g,平均每年增加1.051 g;DAYS_100 从2002年的-0.600 d下降到2020 年的-6.108 d,平均每年下降0.306 d。
3 讨论
大量研究表明,测定场、出生年份、出生季节、性别和出生胎次会影响猪生长性状的表达[9,13-15]。贺婕妤等[9]发现出生年份和出生季节对长白猪DAYS_100、ADG_100 和BFT_100 均有极显著影响(P<0.001);林清等[13]发现测定场、性别对杜洛克猪的DAYS_100、BFT_100 和LMA_100 均有极显著影响(P<0.01);张蕾[14]发现性别对长白后备猪的DAYS_100 和BFT_100 有显著影响(P<0.05);陈海龙[15]也发现性别对长白后备猪的BFT_100 有显著影响(P<0.05)。本研究结果表明,测定场、出生年份、出生季节、性别和出生胎次对性状DAYS_100、ADG_100、LMA_100 和BFT_100 均存在极显著影响(P<0.01),与前人研究结果[9,13-15]一致。测定场对性状值影响显著,说明场间的饲养管理水平和条件差异较大,可能是导致长白猪LMA_100 和BFT_100 的变异系数较大的原因之一。
前人研究发现DAYS_100、ADG_100、LMA_100和BFT_100 遗传力的估计值多接近或超过0.3[9,16-17],本研究结果与之一致,长白猪上述4 个性状的遗传力在0.391~0.433 之间,属于高遗传力性状,可以通过个体直接选择加快遗传进展[18];但也有一些研究对上述性状遗传力的估计值较低,如张蕾[14]估计ADG_100 遗传力为0.02、DAYS_100为0.03、BFT_100 为0.01;曹建新等[19]估计DAYS_100 遗传力为0.210~0.310、BFT_100 为0.140~0.160;张婧[20]估计DAYS_100遗传力为0.08、BFT_100 为0.06。这可能与有效记录数据条数偏少[14]、数据标准差较大[20]、种猪遗传背景和环境不同[19]、估计方法差异[21]等因素有关。此外,本研究没有考虑窝效应[22],可能会导致估计遗传力偏大[23-25]。
本研究结果表明,长白猪DAYS_100 与ADG_100 的表型相关系数和遗传相关系数分别为-0.992 和-0.997,呈现较强的负相关,其他性状的相关性均较小,这与前人研究结论一致。如王青来等[2]报道长白猪30~100 kg 平均日增重与DAYS_100 的遗传相关系数为-0.941,表型相关系数为-0.829。本研究中长白猪DAYS_100 与BFT_100 的遗传相关系数为-0.056,与贺婕妤等[9]的研究结果-0.027 接近;但本研究中上述两性状的表型相关为负相关,而贺婕妤等[9]的研究结果为正相关,证明环境效应会影响性状的表型相关系数。
从整体来看,DAYS_100、ADG_100、BFT_100的表型均值和估计育种值均值呈现相反趋势,说明存在测定误差,即猪活重和BFT_100 的测定值存在偏差,推测可能由于表型测定时未严格遵守测定规范及存在系统误差引起。本研究中,2018 年前DAYS_100、ADG_100 和BFT_100 的表型均值和遗传进展趋势基本一致,虽然遗传进展在2018年后取得一定进步,但3 个性状在表型均值方面的变化并不明显,初步分析表明可能与该群体作为母系猪进行的育种目标性状选择相关;此外,BFT_100 性状的表型均值及遗传进展在2009 年前后出现较大波动,推测可能由于该场生产管理对场间设备进行升级更新有关,也可能与该群体的品种品系结构发生变动相关。LMA_100 的表型趋势和遗传趋势变化高度一致,说明该性状测定值较准确。
4 结论
本研究中长白猪4 个生长性状(DAYS_100、ADG_100、LMA_100 和BFT_100)均属于中高遗传力性状。DAYS_100 和ADG_100 间的遗传相关和表型相关均呈现强相关。整体来看,DAYS_100、ADG_100 和BFT_100 性状的表型进展及遗传进展呈相反趋势,且2018 年后取得一定的遗传进展,表明猪场的表型测定管理及群体育种目标性状的选择对猪群生长性状的表现有影响。LMA_100 的遗传趋势和表型趋势变化高度一致,说明该性状测定值较准确。同时,养殖场生产管理水平的提高和品种品系结构的变化也可能会影响遗传进展。
